八个抗稻瘟病基因在华南籼型杂交水稻中的分布
2017-05-19汪文娟周继勇汪聪颖苏菁封金奇陈炳冯爱卿杨健源陈深朱小源
汪文娟周继勇汪聪颖苏菁封金奇陈炳冯爱卿杨健源陈深朱小源,*
(1广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室,广州510640;2广东省农业技术推广总站,广州510520;*通讯联系人,E-mail:zhuxy@gdppri.com)
八个抗稻瘟病基因在华南籼型杂交水稻中的分布
汪文娟1周继勇2汪聪颖1苏菁1封金奇1陈炳1冯爱卿1杨健源1陈深1朱小源1,*
(1广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室,广州510640;2广东省农业技术推广总站,广州510520;*通讯联系人,E-mail:zhuxy@gdppri.com)
【目的】已克隆的稻瘟病抗性基因Pi1、Pik-p、Pik-h、Pi2、Pi9、Piz-t、Pita、Pii对不同稻区的稻瘟病菌表现较广谱的抗性,被广泛应用于水稻抗瘟性育种。为了明确上述抗性基因在华南稻区杂交稻组合中的分布及其组合的抗病有效性,【方法】利用上述8个抗病基因的功能标记,对华南328个杂交稻组合进行了抗瘟基因型分子检测。【结果】抗性基因Pita和Pii分布频率最高,在测试组合中检出率分别为84.76%与67.68%;其次是Pi2与Pik-p,分别为22.87%与13.72%;检出频率较低的是Pi1、Piz-t和Pik-h,分别为5.18%、3.35%与2.13%,检测的品种都不携带抗性基因Pi9。在单个杂交稻组合中,检出的抗瘟基因数量最多是4个。抗病性评价结果表明,杂交稻组合中检出的抗病基因数量越多,其表现为抗病品种的频率就越高;含4个抗病基因的杂交稻组合中,抗病品种所占比率达91.67%。含有不同抗瘟基因的组合表现出不同水平的抗瘟性,其中Pi2与Pi1对华南稻区稻瘟病的抗病性贡献最大,其他抗病基因的贡献大小依次是Pik-h、Pik-p、Pita、Pii与Piz-t。【结论】本研究为华南稻区杂交稻抗病品种基因型的合理布局以及抗瘟基因的应用提供了科学依据。
杂交稻组合;抗病基因;稻瘟病;基因型分析;分子标记
水稻(Oryza sativa)是世界上重要的粮食作物,全球半数以上的人口以稻米作为主食[1]。由稻瘟病菌(Magnaporthe oryzae)引起的稻瘟病是威胁水稻粮食生产的最具毁灭性的病害之一[2],发病轻时导致减产,严重时局部田块甚至颗粒无收[3]。稻瘟病也是华南水稻生产中最重要的病害,近年来在华南发生日益严重,如华南稻区的天优998、五优308、天优368等主栽品种的抗性相继衰退或丧失,在部分稻区发生了稻瘟病大面积流行或暴发,对粮食的生产安全造成了严重影响[4,5]。目前,选育和种植抗病水稻品种是防治稻瘟病最经济、环保及有效的措施[6]。因此,摸清主栽品种与新选育的抗病品种的抗瘟基因型,是抗病品种科学应用的首要前提。
20世纪60年代中期,日本率先对水稻稻瘟病的抗病基因开展了系统的研究。随后,源自不同水稻品种的90多个主效抗稻瘟病基因被鉴定[7]。近年来,国内研究者对已鉴定的部分抗稻瘟病基因进行了抗性有效性评价。李进斌等[8]分析了22个抗瘟基因在云南省3个稻区的抗性情况,发现Pi9和Piz5在云南稻区具有较好的抗性;杨健源等[9]的研究表明,华南稻区目前有效的抗性基因主要为Pi1、Pi2、Pita等;并且发现目前华南籼稻区育种者利用的抗瘟基因多为Pi1与Pi2[10,11];张国民等[12]分析了24个抗瘟基因在我国寒地稻区的抗性情况,发现Pi9对该稻区的稻瘟病菌表现出广谱抗性。然而,我国大部分稻区水稻品种的抗瘟基因型仍不清晰,有待分析与研究;该方面研究滞后的原因,主要是缺乏基因型快速有效的检测技术。迄今,已经成功克隆出Pib、Pi9、Pi2、Piz-t、Pik、Pik-p、Pik-h、Pi1、Pita、Pii、Pi50和Pi64等26个稻瘟病抗性基因[13,14]。利用基因序列信息,发展特异性功能标记,可快速检测品种的抗稻瘟病基因型[15,16],这对全面了解我国水稻抗稻瘟病的基因型及其合理布局与轮换有着重要意义。
我们利用Pi1、Pik-p、Pik-h、Pi2、Pi9、Piz-t、Pita、Pii等8个抗病基因功能性分子标记,对华南稻区328个杂交水稻组合进行了抗瘟基因型检测,旨在摸清该稻区杂交稻组合的基因型,为华南稻区抗病品种的合理布局及进一步选育新型的抗瘟杂交稻组合提供依据。
1 材料与方法
1.1 植物材料
本研究所用材料包括328个近5年推广及新参试的华南籼型杂交稻组合,其种子由广东省省级水稻品种区域试验主持单位提供。
水稻种子经0.5%强氯精消毒5 min,25℃清水浸泡24 h,滤干种子,置于35℃培养箱中催芽24 h。萌芽的种子以4行×6列的间隔穴播于盛有栽培泥土的瓷盘里(规格30 cm×20 cm×5 cm),每天浇水1~2次。待秧苗长至1叶1心期,每盆施0.5 g硫酸铵,每间隔3 d施用一次,共施3次。待植株长至2.5~3.0叶龄,进行标样的采集。
1.2 病原菌材料、培养及接种调查
1.2.1 菌株来源
室内抗谱测定选用的50~64个菌株为单孢分离菌株,均采自主栽品种。这些菌株来源于广东、广西、福建等地,主要是根据当年或上一年稻瘟病菌生理小种的致病性测定情况挑选出的代表性菌株,对稻瘟病单基因系的致病性上具有丰富的多样性;所有菌株均保存于广东省农业科学院植物保护研究所。
1.2.2 菌株活化、繁殖及产孢
将保存的菌株转移至酵母固体培养基,放于生化培养箱中,活化培养7 d以上;然后将菌丝体转移到高温高压湿热灭菌的玉米培养基上繁殖,在生化培养箱中培养约13 d。产孢:先用灭菌过的无菌水洗去玉米粒表面的菌丝,再将玉米粒置于已消毒的搪瓷盘中(25 cm×19 cm×2 cm)铺开,并在上面覆盖一层湿纱布,然后在日光灯下光照培养3~4 d进行产孢。以上菌株的活化、繁殖及产孢均在25℃下进行。
1.2.3 接种调查
用无菌水将附在玉米粒上的分生孢子洗下,用2层塑料细纱网隔去玉米残渣,在100倍显微镜下镜检,选取浓度为5×105个/mL的孢子悬浮液进行人工喷雾接种;接种后的稻苗置于自制的相对湿度达95%以上的培养箱中暗培养24 h,温度控制在25℃左右。之后转至玻璃温室中,接种7 d后进行调查。病级调查按照国际水稻所稻瘟病圃苗瘟分级标准进行:0~3级为抗,4~9级为感。抗性频率(%)为无毒的菌株数目与总测定菌株数目之比。
1.2.4 抗病性综合评价
依据田间病圃鉴定与室内抗谱测定,综合评价杂交稻组合的抗性。田间病圃分别设在广东稻区的曲江、从化、龙川、阳江、信宜等5个自然病区。室内抗谱测定选用的菌株来源于广东、广西、福建等地,稻瘟病菌株均为单孢分离菌株,接种菌株数为50~64个。依据杂交稻组合在病区的穗瘟病级以及室内抗谱测定的数据,对组合做出综合抗性评判,抗性级别由高至低排序为:高抗、抗、中抗、中感、感、高感等6个级别。调查与评价方法参照朱小源等[17]和冯爱卿等[18]的方法。
1.3 标样采集及DNA提取
每个品种采集2个单株的叶片,混合收集于2.0 mL的离心管中,经液氮速冻后,置于-80℃的冰箱内储藏备用。
首先将装有水稻叶片的离心管放在液氮中浸泡约5 min,随后用快速组织破碎仪磨碎叶片,再用百泰克生物技术(北京)有限公司的新型快速植物基因组DNA提取试剂盒,提取水稻叶片总DNA。
1.4 抗病基因Pi2、Pi9、Pita等8个基因特异性分子标记与检测
1.4.1 抗病基因Pi2、Pi9、Pita等8个基因特异性分子标记
本研究中Pik-p、Pik-h与Pi1基因特异性分子标记是Zhai等[19]与Hua等[20]开发的Pik簇复等位基因间特异性分子标记。首先,基于日本晴参考序列与Pik簇等位基因序列间的较大差异,开发了K1标记,用于区分水稻品种含有的K型(Kusabue/K60)与N型(日本晴)两种基因型[19,21];其次,通过对Pik-m/Pik-p/Pik/Pi1及日本晴的感病等位基因编码区进行序列比对,找出各等位基因特异的SNP,利用dCAPS Finder 2.0软件(http://helix.wustl.edu/dcaps/dcaps.html)[22],针对包含该SNP位点的序列,在其两翼设计引物,开发出能区分Pik簇复等位基因Pik-p、Pik-h与Pi1的dCAPS标记。Pii特异性CAPS标记是Takagi等[23]根据Pii供体品种Hitomeborn与日本晴中pii感病等位基因序列比对,发现在PvuⅡ酶切位点处存在SNP多态性而开发的Pii-CAPS标记。Pi2、Pi9与Piz-t特异性分子标记依据华丽霞等[24]设计的标记。Pita基因特异性标记是通过将Pita全基因序列与日本晴的感病等位基因进行比对,找出特异的SNP,开发出能特异性地检测Pita基因的dCAPS标记(未发表资料)。各标记的具体信息见表1。
1.4.2 抗病基因Pi2、Pi9、Pita等8个基因特异性分子标记的检测
利用开发的Pi2、Pi9、Pita等8个基因特异性分子标记,对328个华南稻区杂交稻组合分别进行PCR扩增,并利用相应的限制性内切酶对PCR产物进行酶切,检测各杂交稻组合的基因型。分子标记PCR扩增:反应总体积为20µL,其中包括水稻基因组DNA(20~30 ng/μL)1 μL,10× PCR缓冲液10 μL,各1 μL正反向引物(10 μmol/L),ddH2O 7 μL。PCR程序如下:94℃下预变性3 min;94℃下变性30 s,55℃~60℃下退火30 s,72℃下延伸1 min,35个循环;72℃延伸5 min。PCR仪上扩增结束后,以各单管的PCR产物为模板,利用相应的限制性内切酶对PCR产物进行酶切,酶切体系为10 μL,其中,包括PCR产物10 μL,10×酶切缓冲液1 μL,限制性内切酶0.3 μL,ddH2O 3.7 μL。各单管酶切混合液,在相应内切酶最适宜的酶切温度下(一般是37℃)酶切3 h,产物加入载样缓冲液后,在6%~8%的聚丙烯酰胺凝胶上进行电泳检测,电泳条件为120 V,2.5~3.0 h。

表1 各基因特异标记检测引物Table 1.Specific marker detecting primers for each gene.

图1 基于特异性SNP分子标记的水稻品种抗瘟基因检测Fig.1.Detection of R genes in rice cultivars with specific SNP markers.
2 结果与分析
2.1 杂交稻组合的抗瘟基因型及其分布
本研究先利用能区别Pik位点K型/N型结构的引物K1,对328个杂交稻组合进行了抗瘟基因检测,结果在211个杂交稻组合中检测到含有K型的Pik等位基因簇;再利用Pik等位基因簇的Pik-p、Pik-h与Pi1的特异性SNP标记,对这211个K型品种进行Pik-p、Pik-h与Pi1基因型分析。结果发现,其中有45个品种含有Pik-p,有17个品种含有Pi1,有7个品种含有Pik-h。此外,利用Pi2、Pi9、Piz-t、Pita及Pii基因特异性SNP标记,分别对该328个杂交稻组合进行了抗瘟基因型检测。结果表明,在这些杂交稻组合中绝大部分能检测到Pi2、Pi9、Piz-t、Pita及Pii等抗病基因及其感病等位基因,部分基因特异性分子标记的检测结果如图1所示。
在检测的杂交稻组合中,抗性基因Pita和Pii分布频率最高,在测试组合中的检出率分别为84.76%与67.68%;其次是抗性基因Pi2和Pik-p,分别为22.87%与13.72%。在这些杂交稻组合中,检出频率较低的抗性基因是Pi1、Piz-t与Pik-h,分别为5.18%、3.35%与2.13%。检测的328个杂交稻组合都不携带抗性基因Pi9(图2)。
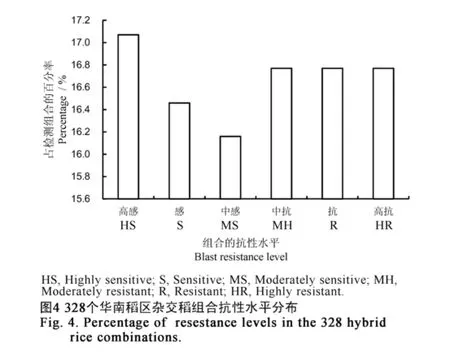
2.2 杂交稻组合聚合的抗瘟基因数量分析


对328个杂交稻组合检出的抗瘟基因数量进行分析,裕优132、恒丰优华占、吉丰优华占与金稻优618等12个杂交稻组合检出了4个抗瘟基因,是检出抗瘟基因数量最多的杂交稻组合,占总检测组合的比率是3.66%。这12个组合中除都检出了Pi2、Pii、Pita基因外,其中有5个组合检出了Pi1基因,有7个组合检出了Pik-p基因。天优613、天优173、永丰优9802与荣优华占等76个杂交稻组合检出了3个抗瘟基因,占总检测组合的比率是23.17%。在这76个杂交稻组合中,绝大多数品种都检出了抗瘟基因Pii与Pita;检出的另一个基因为Pi2、Piz-t、Pi1、Pik-p、Pik-h之一。粤良优778、华两优78、珍丰优9822与美优9802等155个杂交稻组合检出了2个抗瘟基因,占总检测组合的比率为47.25%;其中,大多数组合检出的抗瘟基因是Pii与Pita。另外,部分杂交稻组合只检测出1个抗瘟基因,其比率为20.73%,还有5.18%的组合未检测到该8个抗瘟基因中的任何一个(图3)。
2.3 检出的抗瘟基因数量、类型与杂交稻组合的抗病相关性分析
为了探明杂交稻组合的抗瘟基因类型或数量与品种抗病性间的相关性,本研究对328个杂交稻组合进行了稻瘟病抗谱测定及田间病圃鉴定,结合两组数据对这些组合进行了抗瘟性综合评价。在选取的328个杂交稻组合中,高感、感、中感、中抗、抗、高抗的组合所占比例分别为17.07%、16.46%、16.16%、16.77%、16.77%、16.77%,各抗性水平的杂交稻分布较均衡(图4)。随着杂交稻组合检出的抗病基因数量的增多,为抗病品种的比率也随之上升;其中,在含有4个抗病基因的杂交稻组合中,抗病品种比率达91.67%,在含有3个、2个与1个抗病基因的杂交稻组合中,抗病品种比率分别为76.32%、45.6%与35.29%。上述结果表明,一个品种(组合)中含有的抗病基因数量越多,其抗性品种的出现频率可以得到相应提升。
根据稻瘟病抗性综合评价的结果,进一步分析含Pi1、Pik-p、Pik-h、Pi2、Pi9、Piz-t、Pita、Pii等不同的抗性基因对提升水稻品种(组合)抗性频率的贡献。结果显示,不同的抗性基因对提升水稻品种的抗病性贡献表现出显著差异;其中,携带了抗性基因Pi2的杂交稻组合,表现为抗病品种的频率为93.33%;其次是Pi1、Pik-h和Pik-p,分别为88.24%、71.43%和60.0%;在携带Pita、Pii与Piz-t基因的杂交稻组合中,表现为抗病品种的频率分别为55.03%、49.55%与27.27%。上述结果表明,Pi2与Pi1在华南籼稻区表现出良好的抗瘟性,对该稻区稻瘟病的抗瘟性贡献最大,其他抗瘟基因的抗病性贡献大小依次是Pik-h、Pik-p、Pita、Pii与Piz-t。
3 讨论
了解病原菌的无毒基因以及品种抗病基因的构成是利用抗病品种控制稻瘟病的基础。单基因系的建立便于人们利用单基因鉴别系统鉴定稻瘟病菌无毒基因型,随着越来越多的抗稻瘟病基因被克隆,为利用特异性标记分析品种抗病基因型奠定了技术基础。国内学者利用抗稻瘟病单基因系对我国籼稻区稻瘟病菌的致病性进行了测定,研究表明华南稻瘟病菌与主效抗瘟基因的互作表现出丰富的多样性,但亦可把互作归类为几个主成份类型,即分别以Pi1、Pi2、Pita2、Pish、Piz、Pi9、Pik-h、Pik-p、Pii等9个基因与病原互作的9个互作类型[9,25]。本研究以上述基因为主要检测对象,试图探明上述基因在华南杂交稻的分布以及与抗病性间的相关性,为本地区抗病品种基因型的合理布局提供科学依据。
近年来,国内外学者对水稻品种的抗瘟基因型相继开展了研究。时克等[26]利用基因自身的功能标记检测了58个水稻品种的Pita和Pib基因情况及其分布状况;何重等[15]利用Pi-ta和Pi-b功能标记,对40个粳稻品种和665个粳稻新品系进行了基因型检测,明确了抗性基因Pi-ta和Pi-b在江苏粳稻品种中具有一定的分布,其中Pi-b的分布频率较高。马军韬等[27]对6个主要水稻品种含有的抗瘟基因型进行了分析,共检测到11个抗瘟基
因,Pi1基因出现频率最高。本研究利用Pi1、Pik-p、Pik-h、Pi2、Pi9、Piz-t、Pita、Pii等8个基因功能性标记,对来自华南稻区不同遗传背景的328个杂交稻组合进行了基因筛查,发现在这些品种中分布频率最高的基因是Pita和Pii,这可能是华南稻区选用的育种材料中多含有Pita和Pii基因的缘故。检出率较低的抗病基因是Pi1、Piz-t和Pik-h,在这些品种中均未检测到Pi9基因,这一结果与Tian等[28]的研究认为抗病基因Pi9尚未在中国籼稻品种中广泛应用相符;说明Pi9基因在华南稻区的抗病品种选育及生产上具有重要的应用潜力。本研究亦表明,一个杂交稻组合中检出的抗病基因数量最多是4个,检出3个或4个杂交稻组合的的抗瘟性是否一定优于聚合了2个或单个抗性基因的组合,还有待扩大水稻的检测群体开展进一步的研究。
本研究分析了检测的8个抗病基因对杂交稻组合的抗病性贡献,发现Pi2与Pi1对华南稻作区稻瘟病的抗性贡献最大,这2个抗病基因也是在华南稻区抗病育种工作中应用较多的广谱抗瘟基因。王丰等[29]将稻瘟病广谱抗病基因Pi1和香味基因fgr聚合到恢复系R290中,显著提高了该恢复系的抗瘟性及稻米的香味品质;柳武革等[30]将Pi1、Pi2基因导入到保持系荣丰B中,利用其配组的杂交稻组合表现出优良的稻瘟病抗性和丰产性。近年来,由于自然界中稻瘟病菌的快速变异,生产上已出现了含有广谱抗病基因Pi2与Pi1的品种抗病性衰退或丧失的事件[4,5,31]。据报道,杂交稻天优998严重感病是由于广东省稻瘟病菌稀有小种ZB3上升为优势小种所致,该型小种可致使携带Pi1及其等位基因的水稻品种严重感病[4];五优308在我国南方多个稻作区严重感病,侵染该组合的稻瘟病菌可侵染Pita(Pita2)、Pii、Pish等抗瘟基因[5]。目前,与天优998及五优308抗瘟基因类型相近的杂交稻组合有湘丰优827、双优228、金昌优3550、深优9521、美优116、五丰优587、五丰优165等,这些组合在上述两个组合感病的稻区应避免种植;另外,与天优998及五优308不同类型的杂交稻组合有五丰优9802、五优华占、五丰优116、Y两优187、珍丰优9822、美优9802、天优华占等,这些组合在天优998和五优308感病的稻区可考虑因地制宜地轮换种植。
虽然,本研究分析的抗性基因是目前华南稻区重要的抗瘟基因型,但是,随着新的抗瘟基因不断鉴定、引入及应用,今后仍十分有必要拓展对不同类型的抗瘟基因开展深入分析与研究。
[1]Liu J L,Wang X J,Mitchell T,Hu YJ,Liu X L,Dai L Y, Wang G L.Recent progress and understanding of the molecular mechanisms of the rice-Magnaporthe oryzae interaction.Mol Plant Pathol,2010,11(3):419-427.
[2]Ou S H.Rice Disease.2nd edn.Kew:Commonwealth Mycological Institute,1985:109-201.
[3]Miah G,Rafii M Y,Ismail M R,Puteh A B,Rahim H A, Asfaliza R,Latif M A.Blast resistance in rice:A review of conventional breeding to molecular approaches.Mol Biol Rep,2013,40(3):2369-2388.
[4]朱小源,杨健源,陈玉托,杨维新,陈喜劳,曾列先,陈深.引致天优998抗性丧失的稻瘟病菌小种鉴定及其致病性测定.广东农业科学,2008(12):84-86. Zhu X Y,Yang J Y,Chen Y T,Yang W X,Chen X L, Zeng L X,Chen S.Race identification and pathogenicity test of the blast fungus causing the resistance breakdownof hybrid rice Tianyou 998.Guangdong Agric Sci, 2008(12):84-86.(in Chinese with English abstract)
[5]汪文娟,韦小燕,陈凯玲,陈尉芹,陈珍,杨健源,朱小源.源自杂交稻组合五优308稻瘟病菌致病性分析.广东农业科学,2015,14:70-73. Wang W J,Wei X Y,Chen K L,Chen W Q,Chen Z, YangJY,ZhuXY.Pathogenicityanalysison Magnaporthe grisea of hybrid combination Wuyou 308. Guangdong Agric Sci,2015,14:70-73.(in Chinese with English abstract)
[6]Fjellstrom R,Conaway-Bormans C A,McClung A M, Marchetti M A,Shank A R,Park W D.Development of DNA markers suitable for marker assisted selection of threePigenesconferringresistancetomultiple Pyricularia grisea pathotypes.Crop Sci,2004,44: 1790-1798.
[7]Liu W D,Liu J L,Triplett L,Leach J E,Wang G L. Novel insights into rice innate immunity against bacterial and fungal pathogens.Annu Rev Phytopathol,2014,52: 213-241.
[8]李进斌,李成云,陈艳,雷财林,凌忠专.二十二个抗稻瘟病基因在云南的利用价值评价.植物保护学报, 2005,32(2):113-119. Li J B,Li C Y,Chen Y,Lei C L,Ling Z Z.Evaluation of twenty-two blast resistance genes in Yunnan using monogenetic rice lines.Acta Phytophyl Sin,2005,32(2): 113-119.(in Chinese with English abstract)
[9]杨健源,陈深,曾列先,李亦龙,陈珍,朱小源.稻瘟病主效抗性基因对广东省籼稻稻瘟病菌的抗性评价.中国水稻科学,2008,22(2):190-196. Yang J Y,Chen S,Zeng L X,Li Y L,Chen Z,Zhu X Y. Evaluation on resistance of major rice blast resistance genes to Magnaporthe grisea isolates collected from indica rice in Guangdong Province,China.Chin J Rice Sci,2008,22(2):190-196.(in Chinese with English abstract)
[10]柳武革,王丰,金素娟,朱小源,李金华,刘振荣,廖亦龙,朱满山,黄慧君,符福鸿,刘宜柏.利用分子标记辅助选择聚合Pi-1和Pi-2基因改良两系不育系稻瘟病抗性.作物学报,2008,34(7):1128-1136. Liu W G,Wang F,Jin S J,Zhu X Y,Li J H,Liu Z R, Liao Y L,Zhu M S,Huang H J,Fu F H,Liu Y B. Improvement of rice blast resistance in TGMS Line by pyramidingofPi-1andPi-2throughmolecular marker-assisted selection.Acta Agron Sin,2008,34(7): 1128-1136.(in Chinese with English abstract)
[11]李进波,夏明元,戚华雄.水稻抗稻瘟病基因Pi1和Pi2聚合系的获得及其抗性评价.安徽农业科学,2015, 43(5):12-14,31. Li J B,Xia M Y,Qi H X.Development and evaluation of disease resistance of pyramided lines with blast resistance genes Pi1 and Pi2 in rice.J Anhui Agric Sci,2015,43(5): 12-14,31.(in Chinese with English abstract)
[12]张国民,马军韬,肖佳雷,刘迎雪,辛爱华,任洋,张丽艳,刘东风.已知抗瘟基因在黑龙江省寒地稻区的评价与利用.植物病理学报,2011,41(1):72-79. Zhang G M,Ma J T,Xiao J L,Liu Y X,Xin A H,Ren Y, Zhang L Y,Liu D F.Evaluation and utilization of value of twenty-four blast resistance genes in north cold region, Heilongjiang.Acta Phytopathol Sin,2011,41(1):72-79. (in Chinese with English abstract)
[13]Ma J,Lei C L,Xu X T,Hao K,Wang J,Cheng Z,Ma X, Ma J,Zhang X,Guo X,Wu F,Lin Q,Wang C,Zhai H, Wan J.Pi64,encoding a novel CC-NBS-LRR protein, confers resistance to leaf and neck blast in rice.Mol Plant Microbe Interact,2015,28(5):558-568.
[14]Su J,Wang W J,Han J L,Chen S,Wang C Y,Zeng L X, Feng A Q,Yang J Y,Zhou B,Zhu X Y.Functional divergence of duplicated genes results in a novel blast resistance gene Pi50 at the Pi2/9 locus.Theor Appl Genet, 2015,128(11):2213-2225.
[15]何重,陈涛,张亚东,朱镇,赵庆勇,周丽慧,于新,王才林.江苏部分粳稻品种和品系中稻瘟病抗性基因Pi-ta和Pi-b的基因型分析.江苏农业学报,2014,30(5): 921-927. He C,Chen T,Zhang Y D,Zhu Z,Zhao Q Y,Zhou L H, Yu X,Wang C L.Genotypic analysis of blast resistance genes Pi-ta and Pi-b for japonica rice varieties and lines in Jiangsu Province.Jiangsu Agric Sci,2014,30(5): 921-927.(in Chinese with English abstract)
[16]程芳艳,李春光,刘永巍,孙翊轩,孟昭河,徐正进.水稻抗瘟基因Pi-b与Pi-5的分子检测及外引抗源的利用评价.核农学报,2016,30(1):11-18. Cheng F Y,Li C G,Liu Y W,Sun Y X,Meng Z H,Xu Z J.Molecular detection of blast resistance genes Pi-b and Pi-5 in northeast rice varieties and utilization evaluation of introduced resistant donors.J Nuclear Agric Sci,2016, 30(1):11-18.(in Chinese with English abstract)
[17]朱小源,杨健源,刘景梅,司徒志谋,康金平,胡学应,朱敏记,罗森辉,杨祁云,林佩珍,曾列先,姜先芽,陈深.广东水稻品种抗稻瘟病性分析与利用策略.广东农业科学,2006(5):34-37. Zhu X Y,Yang J Y,Liu J M,Situ Z M,Kang J P,Hu X Y,Zhu M J,Luo S H,Yang Q Y,Lin P Z,Zeng L X, Jiang X Y,Chen S.Evaluation on resistance of rice varieties in Guangdong to rice blast and strategy for its utilization.Guangdong Agric Sci,2006(5):34-37.(in Chinese with English abstract)
[18]冯爱卿,杨健源,陈深,曾列先,杨祁云,苏菁,汪文娟,朱小源.水稻资源对稻瘟病质量抗性及数量抗性评价.广东农业科学,2015,42(12):27-32. Feng A Q,Yang J Y,Chen S,Zeng L X,Yang Q Y,Su J, Wang W J,Zhu X Y.Evaluation on qualitative and quantitative resistance of rice germplasm to Magnaporthe oryzae.2015.Guangdong Agric Sci,2015,42(12):27-32. (in Chinese)
[19]Zhai C,Lin F,Dong Z Q,He X,Yuan B,Zeng X,Wang L,Pan Q.The isolation and characterization of Pik,a rice blastresistancegenewhichemergedafterrice domestication.New Phytol,2011,189(1):321-334.
[20]Hua L X,Wu J Z,Chen C X,Wu W H,He X Y,Lin F, Wang L,Ashikawa I,Matsumoto T,Wang L,Pan Q H. The isolation of Pi1,an allele at the Pik locus which confers broad spectrum resistance to rice blast.Theor Appl Genet,2012,125:1047-1055.
[21]Yuan B,Zhai C,Wang W J,Zeng X S,Xu X K,Hu H Q, Lin F,Wang L,Pan Q H.The Pik-p resistance to Magnaporthe oryzae in rice is mediated by a pair of closely linked CC-NBS-LRR genes.Theor Appl Genet, 2011,122:1017-2108.
[22]Neff M M,Turk E,Kalishman M.Web-based primer design for single nucleotide polymorphism analysis. Trends Genet,2002,18(12):613-615.
[23]Takagi H,Uemura A,Yaegashi H,Tamiru M,Abe A, MitsuokaC,UtsushiH,NatsumeS,KanzakiH, Matsumura H,Saitoh H,Yoshida K,Cano L M,Kamoun SO,TerauchiR.MutMap-Gap:Whole-genome sequencing of mutant F2progeny bulk combined with de novo assembly of gap regions identifies the rice blast resistance gene Pii.New Phytol,2013,200(1):276-283.
[24]华丽霞,汪文娟,陈深,汪聪颖,曾烈先,杨健源,朱小源,苏菁.抗稻瘟病Pi2/9/z-t基因特异性分子标记的开发.中国水稻科学,2015,29(3):305-310. Hua L X,Wang W J,Shen C,Wang C Y,Zeng L X, Yang J Y,Zhu X Y,Su J.Development of specific DNA markers for detecting the rice blast resistance gene alleles Pi2/9/z-t.Chin J Rice Sci,2015,9(3):305-310.(in Chinese with English abstract)
[25]朱小源,杨祁云,杨健源,雷财林,王久林,凌忠专.抗稻瘟病单基因系对籼稻稻瘟病菌小种鉴别力分析.植物病理学报,2004,34(4):361-368. Zhu X Y,Yang Q Y,Yang J Y,Lei C L,Wang J L,Ling Z Z.Differentiation ability of monogenic lines to Magnaporthe grisea in indica rice.Acta Phytopathol Sin, 2004,34(4):361-368.(in Chinese with English abstract)
[26]时克,雷财林,程治军,许兴涛,王久林,万建民.稻瘟病抗性基因Pita和Pib在我国水稻主栽品种中的分布.植物遗传资源学报,2009,10(1):21-26. Shi K,Lei C L,Cheng Z J,Xu X T,Wang J L,Wan J M. Distribution of two blast resistance genes Pita and Pib in major rice cultivars in China.J Plant Genet Res,2009, 10(1):21-26.(in Chinese with English abstract)
[27]马军韬,张国民,辛爱华,张丽艳,邓凌韦,任洋.水稻品种抗稻瘟病分析及基因聚合抗性改良.植物保护学报,2016,43(2):177-183. Ma J T,Zhang G M,Xin A H,Zhang L Y,Deng L W, Ren Y.Resistance analysis and improvement of rice varieties by gene pyramiding.Acta Phytophyl Sin,2016, 43(2):177-183.(in Chinese with English abstract)
[28]Tian D G,Chen Z J,Chen Z Q,Zhou Y C,Wang Z H, WangF,ChenSB.Allele-specificmarker-based assessment revealed that the rice blast resistance genes Pi2 and Pi9 have not been widely deployed in Chinese indicaricecultivars.Rice,2016,9(1):19.doi: 10.1186/s12284-016-0091-8.
[29]王丰,柳武革,刘振荣,朱小源,李金华,廖亦龙,朱满山,黄慧君,杨健源.利用分子标记辅助选择聚合Pi-1和fgr基因改良水稻恢复系.杂交水稻,2010(S1): 237-244. Wang F,Liu W G,Liu Z R,Zhu X Y,Li J H,Liao Y L, Zhu M S,Huang H J,Yang J Y.Pyramiding Pi1 and fgr genes to improve rice restorer lines by molecular marker-assistedselection.HybridRice,2010(S1): 237-244.(in Chinese with English abstract)
[30]柳武革,王丰,刘振荣,朱小源,李金华,黄慧君,廖亦龙,朱满山,付崇允,陈建伟.利用分子标记技术聚合Pi-1和Pi-2基因改良三系不育系荣丰A的稻瘟病抗性.分子植物育种,2012,10(5):575-582. Liu W G,Wang F,Liu Z R,Zhu X Y,Li J H,Huang H J, Liao Y L,Zhu M S,Fu C Y,Chen J W.Improvement of rice blast resistance in CMS line Rongfeng A by pyramidingPi-1andPi-2withmolecularmarker techniques.Mol Plant Breeding,2012,10(5):575-582. (in Chinese with English abstract)
[31]汪文娟,苏菁,张杰,李亦龙,陈深,曾列先,杨健源,朱小源.源于粤晶丝苗2号穗瘟的稻瘟病菌致病性分析.广东农业科学,2012,23:59-61. Wang W J,Su J,Zhang J,Li Y L,Chen S,Zeng L X, Yang J Y,Zhu X Y.Pathogenicity analysis of the rice blastfungusisolatedfromtheblastpaniclesof Yuejingsimiao 2.Guangdong Agric Sci,2012,23:59-61. (in Chinese)
Distribution of Eight Rice Blast Resistance Genes in indica Hybrid Rice in China
WANG Wenjuan1,ZHOU Jiyong2,WANG Congying1,SU Jing1,FENG Jinqi1,CHEN Bing1,FENG Aiqing1, YANG Jianyuan1,CHEN Shen1,ZHU Xiaoyuan1,*
(1Plant Protection Research Institute,Guangdong Academy of Agricultural Sciences/Guangdong Provincial Key Laboratory of High Technology for Plant Protection,Guangzhou 510640,China;2Guangdong Agricultural Technology Extension Station,Guangzhou 510520,China;*Corresponding author,E-mail: zhuxy@gdppri.com)
【Objective】The cloned rice blast resistant genes,Pi1,Pik-p,Pik-h,Pi2,Pi9,Piz-t,Pita and Pii,which showed broad-spectrum blast resistance in different rice growing regions,have been widely used in rice breeding for blast resistance.To clarify the distribution of these genes in hybrid rice in South China,【Method】the genotypes of resistance genes Pi1,Pik-p,Pik-h,Pi2,Pi9,Piz-t,Pita and Pii in 328 hybrid rice combinations were detected with functional markers.【Result】The distribution frequency of Pita and Pii was the highest,reaching 84.76%and 67.68%in the tested rice combinations,respectively;followed by Pi2 and Pik-p with the proportion of 22.87%and 13.72%, respectively;Pi1,Piz-t and Pik-h had the lowest distribution frequency,accounting for 5.18%,3.35%and 2.13%of total; all of these varieties didn’t carry resistance gene Pi9.The number of blast resistance genes detected in hybrid rice combinations were at most four.The resistance evaluation showed that the frequency of resistant varieties rose with increasing number of resistance genes in hybrid combinations.In a hybrid combinations with four resistance genes,the distribution frequency of resistant varieties was 91.67%.Blast resistance contribution assay of the eight resistance genes revealed that Pi2 and Pi1 displayed the greatest resistance contribution in the local rice in South China,followed by Pik-h,Pik-p,Pita,Pii and Piz-t.【Conclusion】This study provided a scientific basis for the rational distribution of resistant varieties with different genotypes in South China.
hybrid rice combinations;resistance gene;rice blast;genotype analysis;molecular marker
S435.111.4+1;S511.034
:A
:1001-7216(2017)03-0299-08
2016-07-26;修改稿收到日期:2016-11-05。
国家重点研发计划项目(2016YFD0300700);广东省科技计划资助项目(2015B020231003;2016B090918116;2016A050502030);国家水稻产业技术体系资助项目(CARS-01-24)。
