小麦品种周麦22号的分子遗传基础及其特异引物筛选
2017-05-15邹少奎殷贵鸿唐建卫韩玉林李顺成李楠楠王丽娜
邹少奎,殷贵鸿,唐建卫,韩玉林,李顺成, 李楠楠,黄 峰,王丽娜,张 倩,高 艳
(周口市农业科学院/河南省小麦种质改良工程技术研究中心,河南周口 466001)
小麦品种周麦22号的分子遗传基础及其特异引物筛选
邹少奎,殷贵鸿,唐建卫,韩玉林,李顺成, 李楠楠,黄 峰,王丽娜,张 倩,高 艳
(周口市农业科学院/河南省小麦种质改良工程技术研究中心,河南周口 466001)
国审小麦品种周麦22号具有高产、稳产、多抗和广适等突出优点,是目前全国种植面积居于前列的品种,也是优异的育种亲本材料。为研究周麦22号的分子遗传学基础以及筛选周麦22的特异引物,利用覆盖小麦全基因组的340个SSR标记对周麦22号及其亲本周麦12号、温麦6号、周麦13号进行SSR标记分析。结果表明,温麦6号对周麦22号的遗传贡献最大(37.35%),其次是周麦13号(36.14%),周麦12号贡献最小(26.51%);周麦22号和其3个亲本间的遗传相似系数的聚类结果与系谱分析不一致,表明在选育过程中亲本遗传物质的传递发生了偏分离;在不同基因组水平上,3个亲本对周麦22号的遗传贡献率差异较明显,在A、B、D三个染色体组上对周麦22号的遗传贡献各有侧重;从340个SSR标记中筛选出28个周麦22号的特异标记,并通过与周麦22号的姊妹系、相似品种、衍生品种及黄淮麦区主推的小麦品种相比较,进一步筛选出1个周麦22号的特异引物Xgwm577,建立了一种能够准确、快速、简便、稳定检测周麦22号品种真实性的检测手段,为周麦22号进一步的遗传改良和推广应用提供了理论参考。
普通小麦;周麦22号;分子遗传;特异引物
小麦是全世界分布最广、种植面积和贸易总量最大的作物。近年来,我国小麦虽连年丰收,但仍处于“紧平衡”状态,迫切需要大幅度提高小麦单位面积产量。在小麦增产诸因素中,新品种的贡献十分突出,约占40%左右。随着小麦新品种逐年增多,对植物新品种权的保护也越来越受到重视,因此,建立一种简便、快捷、有效的鉴定小麦新品种的方法至关重要。
在过去,育种家通过常规系谱分析追溯小麦品种的亲缘关系,但这种方法存在准确性差、不能从分子水平解释遗传构成等问题。随着科技的发展,分子标记技术已广泛应用于小麦骨干亲本及其衍生后代的遗传多样性分析。简单重复序列(Simple sequence repeat,SSR)标记,具有数量多、位置明确,具有共显性、多态性好且稳定等特点,在小麦遗传关系分析和基因定位领域被广泛应用。前人关于小麦遗传构成方面的研究很多。陈国跃等[1]研究揭示了繁6及其衍生品种的遗传规律;王珊珊等[2]和肖静等[3]分析了矮孟牛及其衍生后代的遗传多样性;李琼等[4]分析了小偃6号及其衍生品种(系)的遗传多样性。但前人的研究多数是关于小麦骨干亲本与衍生后代或姊妹系之间的遗传差异,在单一品种与其亲本之间遗传物质的传递规律和遗传差异方面的研究较少。赵春华等[5]研究了科农9204重要染色体位点在衍生后代的传递规律,发现优异基因具有较高的传递率;邹少奎等[6]揭示了周麦23号与亲本间遗传物质的传递规律;李 俊等[7]发现川麦104较多地继承了川麦42的遗传物质,聚合了提高千粒重的QTL位点。
周麦22号由河南省周口市农科院以周麦12号/温麦6号//周麦13号杂交组合选育而成,2007年通过国家农作物品种审定,属半冬性小麦新品种,具有高产、稳产、多抗、广适等优点。目前,周麦22号是国内第二大和河南省第一大推广小麦品种,累计推广面积533万hm2。周麦22号综合性状优良,产量潜力巨大[8],利用其作为亲本已育成国审周麦26、国审周麦28、新麦32、郑麦1860、中育1215和中育1220等20多个小麦新品种,应用前景十分广阔。本研究利用SSR分子标记技术解析周麦22号21条染色体的分子遗传结构,旨在探究周麦22号的分子遗传学基础,同时筛选出1~2个特异引物,以期建立一种简便、快捷、有效的鉴定周麦22号品种真实性的检测手段。
1 材料与方法
1.1 试验材料
试验用种子由河南省周口市农科院小麦研究所提供,遗传构成分析所用的种子包括周麦22号的标准种子(国家种质资源库植物新品种权保护种子)及其3个亲本周麦12号、温麦6号和周麦13号的高纯度种子。特异引物筛选所用的种子共计41份,具体见表1。
1.2 SSR引物标记来源
选用覆盖小麦全基因组的340对SSR引物标记,包括Genomic-SSR引物(BARC、CFA、CFD、GDM、WMC系列)和EST-SSR引物(CWM、KSUM、CWEM、SWES和CNL系列)。各引物标记的序列信息通过GrainGenes2.0 (http://wheat.pw.usda.gov/)查阅获得。
1.3 基因组DNA提取
采用SDS单籽粒DNA提取法提取小麦基因组DNA,每个品种取3粒种子,充分研碎后放入一只2 mL离心管中加入1 mL DNA 提取液(1 mol·L-1Tris-HCl (pH 8.0)、5 mol·L-1NaCl、0.5 mol·L-1EDTA(pH 8.0)、10% SDS )和2 μL β-巯基乙醇;65 ℃下轻摇30~60 min,使其充分混匀;4 ℃下 12 000 r·min-1离心15 min;移上清液至一新的2 mL离心管中,再加等体积的酚/氯仿(1∶1),轻摇5~10 min;4 ℃下12 000 r·min-1离心15 min;移上清至一新的2 mL离心管中,重复5~6步骤;移上清至一新的2 mL离心管中,再加等体积的氯仿/异戊醇(24∶1),轻摇5~10 min;4 ℃下12 000 r·min-1离心15 min;移上清至一新的1.5 mL离心管中,再加0.6倍上清液体积的异丙醇,轻摇混匀,切勿剧烈震荡,之后置于-20 ℃沉淀1 h以上;4 ℃下12 000 r·min-1离心15 min,移上清液,再加0.5 mL 70%的乙醇,静置5 min;4 ℃下12 000 r·min-1离心15 min,去70%的乙醇;自然干燥1.5 h以上;加100 μL的双蒸水,过夜。
1.4 PCR扩增及扩增产物的电泳
PCR反应在美国伯乐公司(BIO-RAD)S1000-PCR仪上运行,每管反应体系为15 μL,包括2×Taq PCR Mix (0.1 UTaq·μL-1,500μmol·L-1dNTPs each,20 mmol·L-1Tris-HCl,100 mmol·L-1KCl,3 mmol·L-1MgCl2) 7.5 μL,引物(10 μmol·L-1)各1 μL,模板DNA 1.5 μL,超纯水4 μL。2×TaqPCR Mix 购于北京汇天东方科技有限公司。SSR扩增程序为94 ℃预变性5 min;94 ℃变性1 min,50 ℃~60 ℃退火1 min(视具体引物而定),72 ℃延伸1 min,35个循环;72 ℃延伸7 min。最后PCR产物置于4 ℃保存。扩增产物采用6%变性聚丙烯酰胺凝胶电泳进行分离,硝酸银染色并拍照留存,最后统计电泳条带的类型。根据电泳结果,按照在相同迁移率的位置上有带记为1,无带记为0的方法,记录每个样品的电泳条带,创建0、1数据库[9-10]。
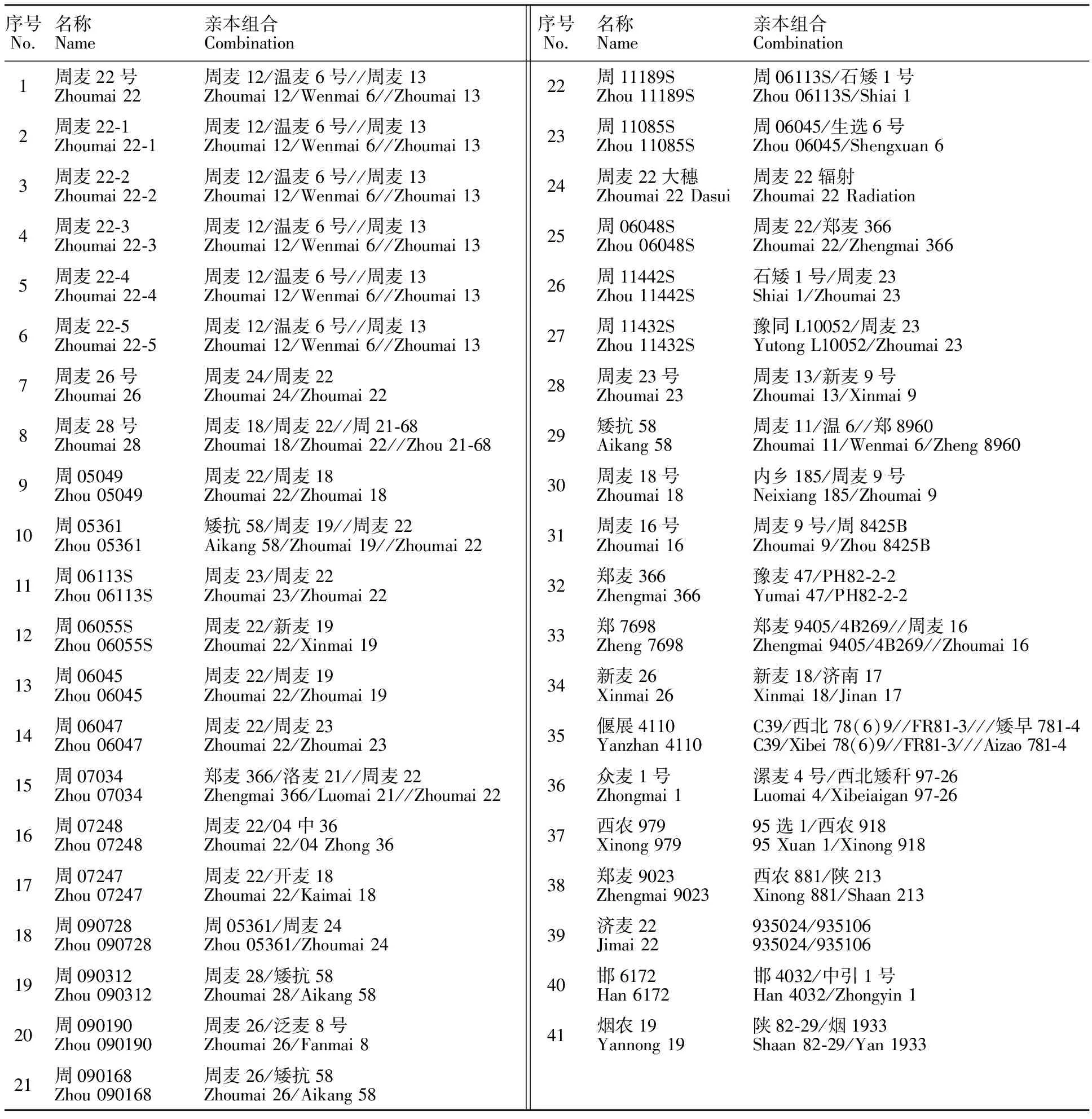
表1 供试材料及其亲本组合
1:周麦22号的标准种子(国家种质资源库植物新品种权保护种子);2~6:周麦22号5个姊妹系;7~23:周麦22号的衍生品种; 24~27:田间长相与周麦22号非常相似的新品种;28~41:目前黄淮麦区主推的小麦品种。
1: Standard seeds of Zhoumai 22(seeds of national germplasm repository of new plant varieties protection); 2-6:Five sib-lines of Zhoumai 22; 7-23:Derived cultivars of Zhoumai 22; 24-27:New varieties look like Zhoumai 22 in the field; 28-41:main wheat varieties in huanghuai region.
1.5 亲本对周麦22号的遗传位点数统计
亲本与子代之间的电泳带型呈多态性,若子代在染色体上的某一位点与其一个亲本的带型相同,且与该杂交组合中的其他亲本的带型不同,则认为该子代继承了这个亲本的遗传位点。统计亲本对子代有贡献的遗传位点数,亲本对子代的遗传贡献率=子代继承一个亲本的位点数/其来自所有亲本的总位点数[11]。
1.6 基因型图谱构建
根据Somers等[12]报道的小麦遗传图谱中的标记顺序,将本研究所用标记按照其在不同染色体上的位置,从短臂至长臂方向排列,利用软件GGT2.0(http://www.dpw.wau.nl/pv/pub/ggt/)绘制基因型图谱。各组合中的亲本根据其带型的异同,在染色体上赋予不同的颜色,而子代根据其与亲本带型的异同也赋予不同的颜色。
1.7 周麦22号特异引物的筛选
利用筛选出的周麦22号不同于3个亲本的引物,先对周麦22号及其姊妹系(1~6号)进行筛选,从中筛选出的引物再对周麦22号衍生的新品种(7~27号)进行筛选,最后再选用黄淮南片麦区主推品种(28~41)进行检测验证。小麦基因组DNA提取、PCR及电泳具体方法同上。
2 结果与分析
2.1 三个亲本对周麦22号的遗传贡献率
340个SSR标记中,287个能扩增出清晰的条带,其中199个在4个供试材料间表现多态性,多态性比例达69.3%。周麦22号在83个标记位点可以确定其亲本来源,其中,来自亲本周麦12号、温麦6号、周麦13号的位点数分别为22、31、30个;176个标记来自2个或以上亲本,不能确定其亲本来源;28个标记位点与3个亲本均不相同,为等位变异位点(表2)。在全基因组水平,周麦22号染色体遗传物质中,26.51% 来自周麦12号、37.35% 来自温麦6号、36.14% 来自周麦13号,而其理论遗传贡献率应该分别是25%、25%、50%。对参试材料做UPGMA聚类分析,结果表明,4个品种间遗传相似系数在0.44~0.65之间,平均为0.54(图1)。同时,亲本周麦12号、温麦6号、周麦13号与周麦22号的遗传距离分别为0.48、0.62、0.47,经过分析发现,4个品种间亲缘关系不一致,即温麦6号对周麦22号的遗传贡献率最高,但UPGMA聚类结果品种间遗传距离表明,温麦6号与周麦22号之间遗传关系较远。
基因组水平上的遗传贡献率计算方法(以基因组A为例):在A基因组上,子代继承一个亲本的遗传位点数与继承所有亲本遗传位点数的比值。在不同基因组水平上,分析3个亲本对周麦22号的遗传贡献率(图2)可知,在A基因组上,温麦6号对周麦22号的遗传贡献率为54.2%,高于周麦12号(16.7%)和周麦13号(29.1%);在B基因组上,周麦13号对周麦22号的遗传贡献率为46.3%,高于周麦12号(22.0%)和温麦6号(31.7%);在D基因组上,周麦12号对周麦22号的遗传贡献率为50%,高于温麦6号(27.8%)和周麦13号(22.2%)。以上结果说明,3个亲本对周麦22号A、B、D基因组的遗传贡献各有侧重。
在染色体水平上,3个亲本对周麦22号的遗传贡献率变化较大(图2)。3个亲本对周麦22号的遗传贡献率范围都在0~100%内。周麦12号在4D上,温麦6号在1A、3A、4A、1D、6D 上,以及周麦13号在4B、6B上对周麦22号的遗传贡献率均达到100%。
2.2 周麦22号与其3个亲本的基因型图谱
查阅Somers等[12]和GrainGenes 2.0(http://wheat.pw.usda.gov)上的遗传图谱信息,得到部分多态性引物在染色体上的遗传距离,然后采用软件 GGT2.0 绘制出周麦22号及其亲本的基因型图谱(图 3),但有 14个引物(包括CWM、KSUM、CWEM、SWES和CNL等系列)没有查到相关位点信息,故没有整合到基因型图谱上。
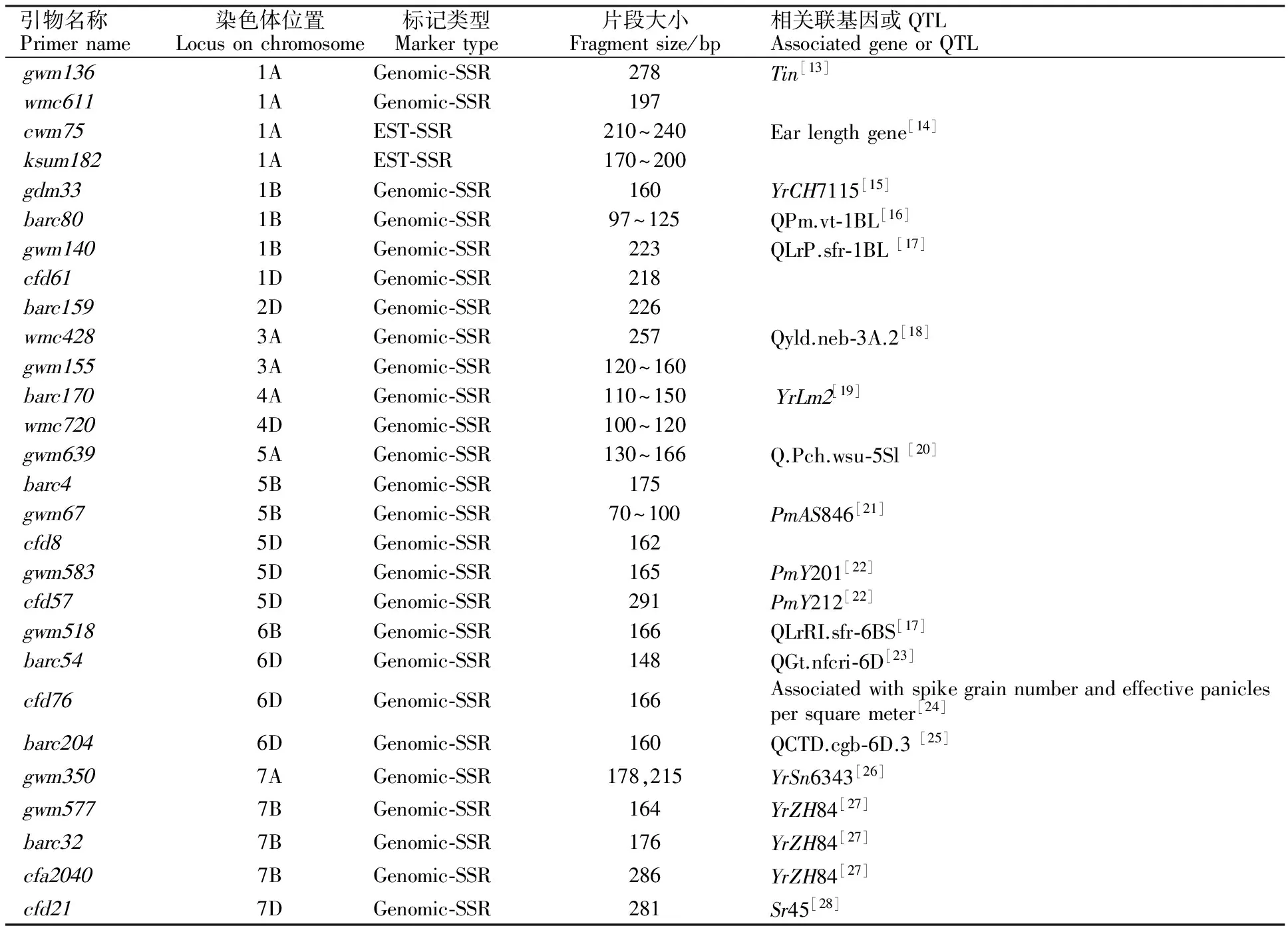
表2 周麦22号的28个特异引物

1:周麦12号; 2:温麦6号;3:周麦13号;4:周麦22号。
1:Zhoumai 12;2:Wenmai 6;3:Zhoumai 13;4:Zhoumai 22.
图1 3个亲本与周麦22号的UPGMA聚类图
Fig.1 UPGMA cluster tree of the Zhoumai 22 and its parents
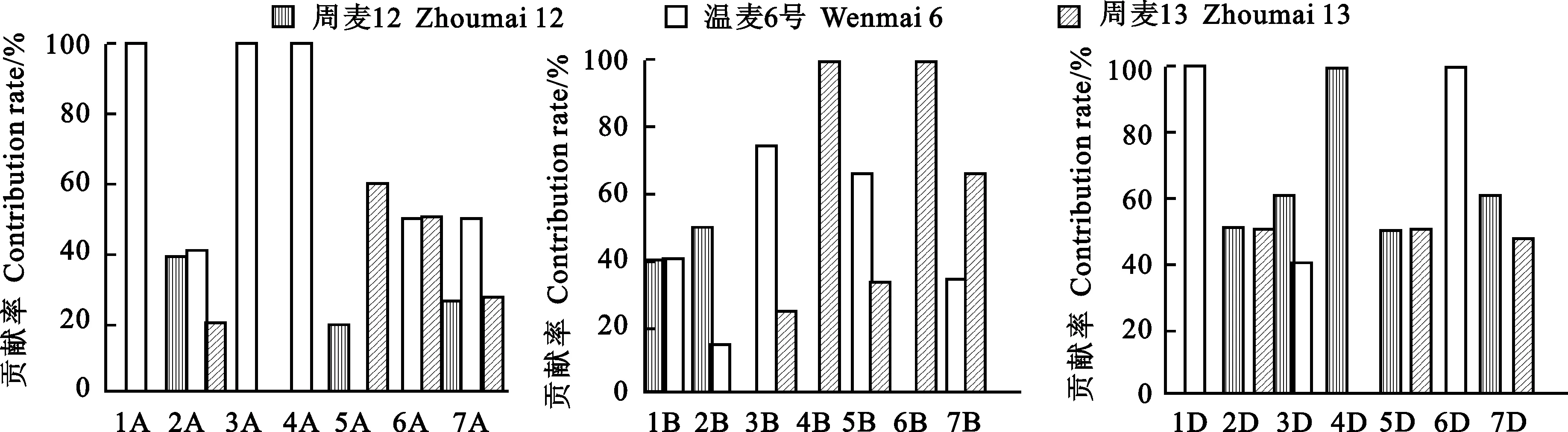
图2 亲本对周麦22号在不同染色体及基因组的遗传贡献率

(图续转下页 Be continued in next page)

1:周麦12号;2:温麦6号;3:周麦13号;4:周麦22号。红色:周麦12区段;蓝色:温麦6号区段;紫色:周麦13号区段;粉红色:周麦22号特有区段;绿色:周麦12号、温麦6号和周麦13号共有区段;黄色:缺失区段。
1:Zhoumai 12; 2:Wenmai 6; 3:Zhoumai 13; 4:Zhoumai 22.Red:Zhoumai 12 fragments; Blue:Wenmai 6 fragments; Violet:Zhoumai 13 fragments; Pink:Zhoumai 22 specific fragments; Green:Identical fragments among Zhoumai 12,Wenmai 6 and Zhoumai 13; Yellow:The missing fragments.
图3 周麦22号与其亲本在21条染色体上的SSR基因型图谱
Fig.3 Genotypic patterns of the samples in the Zhoumai 12/Wenmai 6//Zhoumai 13 cross based on SSR markers
2.3 周麦22号特异引物的筛选结果
在筛选周麦22号特异引物的研究中,获得1个稳定性较好的引物Xgwm577,能把周麦22号与其姊妹系、相似品种、黄淮麦区主推品种很好的区分开(图4)。图4中,1号为周麦22号,扩增出4条带,2~6号为周麦22号的姊妹系,均扩增出3条带,所以周麦22号与其姊妹系为不同的品种。7~23号材料中,11号和20号是1条带,8号和19号为2条带,21号为4条带,其余各品种的均为3条带。21号和周麦22号(1号)同为4条带,但很明显,周麦22号与21号材料的带型差异很大,故周麦22号与7~23号材料中各品种均不相同,因此可将周麦22号与其衍生品种区分开来。24~27号材料是田间农艺性状与周麦22号非常相似的品种,且均扩增出3条带,故与周麦22号不相同。28~41号为黄淮麦区的主推品种。28、38、41号扩出1条带,30、31、33、34、37号扩出2条带,40号为4条带,其余各品种均是3条带。40号与周麦22号的电泳条带数目相同,由图4可以看出,二者有较大差异,故40号与周麦22号属于不同的小麦品种,因此将周麦22号与各品种区分开。综上可知,引物Xgwm577能够将周麦22号与本试验参试的其他小麦品种完全区分开来。由此可以初步判定,这个引物可以作为周麦22号的特异引物,用于检测小麦品种周麦22号的品种真实性。
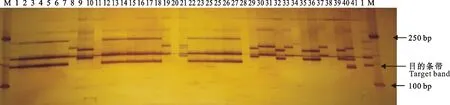
M:Marker;1:周麦22号;2~6:周麦22号姊妹系;7~23:周麦22号衍生品种;24~27:田间性状与周麦22号相似的品种;28~41:黄淮麦区主推品种。
M:Maker; 1:Zhoumai 22; 2-6:Sister lines of Zhoumai 22; 7-23:Derived varieties of Zhoumai 22; 24-27:Varieties look like Zhoumai 22; 28-41:Main varieties in Huanghuai wheat area.
图4 SSR标记Xgwm577扩增参试材料的PCR产物在变性聚丙烯酰胺凝胶上的电泳图
Fig.4 Electrophoresis of PCR products amplified with SSR marker Xgwm577 in polyacrylamide gel
3 讨 论
3.1 SSR标记在遗传构成分析中的应用
周麦22号的亲本组合为周麦12号/温麦6号//周麦13号,而周麦12号、周麦13号均含有周8425B(A)的血缘,可能因为遗传了周8425B较多优良基因[29],周麦22号因此成为目前黄淮南片麦区产量较高、综合性状较好、主栽的半冬性小麦品种。周8425B是河南周口市农科院通过小黑麦与遗传背景不同的普通小麦杂交、回交、辐射和阶梯杂交改良技术集优利用,创育出的矮秆、大穗、抗病新种质,携带1个抗条锈病新基因YrZH84和抗叶锈病新基因LrZH84。周8425B是高产抗病的优异种质,其作为骨干亲本,已育成衍生品种(系)100多个,其中矮抗58、郑麦7698、郑麦379、新麦23、平安8号、洛麦21、洛麦22、漯麦18、中麦895、西农882、农大1108、徐麦9074和淮麦0705等国家和省级审定的品种79个,已成为目前黄淮南片麦区最重要的骨干亲本。
Bohn等[30]和Plaschke等[31]认为,在研究普通小麦品种间的遗传差异方面,SSR标记较RELP和RAPD标记,其多态性和检测效率更高。前人在小麦骨干亲本及其衍生后代或姊妹系之间的遗传解析,在很大程度上促进了小麦骨干亲本衍生规律的研究。袁园园等[32]研究了碧蚂4号的遗传构成及其特异位点在衍生后代中的传递规律;徐鑫等[33]认为系选品种可以缩短品种选育时间,且与亲本的遗传差异较大,可以获得优异的甚至超亲的小麦品种。本研究利用覆盖小麦全基因组的340个SSR标记对周麦22号及其亲本进行遗传解析,阐明了周麦22号分子遗传构成。本研究发现,亲本的遗传物质在传递过程中发生了偏分离行为。3个亲本遗传基础差异较大,遗传相似系数的聚类结果与系谱分析不一致。分析其可能的原因,一是周麦12号(周8425A/SW73295)和周麦13号(周8425B/周麦9号)的亲缘关系太近,在进行电泳带型分析时舍去的非特异性带较多;二是育种者在选育品种时偏好矮秆、大穗大粒等某些性状,使选育的后代遗传物质发生偏离,因此导致品种间亲缘关系不一致。前人对玉米[34]、大豆[35]、大麦[36]的研究中也发现亲本遗传贡献在基因组水平与系谱分析结果存在不一致性,与本研究的结论相似。
3.2 特异引物筛选在品种真实性检测中的应用
本研究筛选出周麦22号的Xgwm136、Xgdm33、Xgwm67、Xbacr32、Xcfa2040、Xgwm577等28个特异标记位点,可以用来鉴定周麦22号和其3个亲本,但最终仅筛选出1个特异标记Xgwm577可用来快速、简便、有效的鉴定区分周麦22号与其姊妹系、衍生品种、外观相似品种和黄淮麦区主推品种。李在峰[27]用抗病亲本周8425B、感病亲本中国春和杂交后代F1、F2和F3在周8425B的7B染色体上定位了抗条锈病基因YrZH84,其与两侧标记Xgwm577、Xcfa2040和Xbacr32紧密连锁;殷贵鸿[37]用标记Xrga-1、Xcfa2040-7B检测了周8425B的衍生品种和黄淮麦区的主推品种,证实周麦22号的7B染色体上存在抗条锈病基因YrZH84。周麦22号农艺性状优良,其他特异标记是否与其高产、抗病、优质等性状关联,还有待进一步试验验证。
小麦种子推广过程中,侵权假冒现象较多。通过籽粒、幼苗和植株等性状的表现型进行鉴定(往往需要种植小区),需要经验丰富的专家进行,往往存在时间较长、人为因素影响等弊端,法院不易采用;采用蛋白标记[38]鉴定小麦品种的真实性往往存在条带较多、受种植环境制约,难以鉴定亲缘关系较近的品种等问题。正是鉴定小麦真伪的技术存在诸多困难,导致很难开展小麦新品种维权打假工作,严重制约小麦种子产业健康发展。
本研究从28对引物中筛选出1个特异引物Xgwm577,能将周麦22号与其姊妹系、衍生品种、相似品种和黄淮麦区主推品种区分开来,初步认定Xgwm577可作为周麦22号的特异引物,建立了一种快速、简便、准确的鉴定周麦22号品种真实性的检测手段。虽然本研究用于检测的品种都是黄淮麦区的主推品种,但用于检验验证的主推品种数目还是偏少,还需要进一步扩大验证品种的范围。
[1] 陈国跃,刘 伟,何员江,等.小麦骨干亲本繁6条锈病成株抗性特异位点及其在衍生品种中的遗传解析[J].作物学报,2013,39(5):827.
CHEN G Y,LIU W,HE Y J,etal.Specific loci for adult-plant resistance to stripe rust in wheat founder parent Fan 6 and their genetic dissection in its derivatives [J].ActaAgronomicaSinica,2013,39(5):827.
[2] 王珊珊,李秀全,田纪春.利用SSR标记分析小麦骨干亲本“矮孟牛”及衍生品种(系) 的遗传多样性[J].分子植物育种,2007,5(4):485.
WANG S S,LI X Q,TIAN J C.Genetic diversity of main parent of wheat 'Aimengniu' and pedigree on SSR markers [J].MolecularPlantBreeding,2007,5(4):485.
[3] 肖 静,王珊珊,李秀全,等.矮孟牛及其后代育成品种的多态性分析与指纹鉴定[J].分子植物育种,2009,7(3):483.
XIAO J,WANG S S,LI X Q,etal.Genetic diversity analysis and fingerprint identification of 'Aimengniu' and its deritives[J].MolecularPlantBreeding,2009,7(3):483.
[4] 李 琼,王长有,刘新伦,等.小偃6号及其衍生品种(系)遗传多样性的SSR分析[J].麦类作物学报,2008,28(6):950.
LI Q,WANG C Y,LIU X L,etal.Genetic diversity of Xiaoyan 6 and its deritives by SSR [J].JournalofTriticeaeCrops,2008,28(6):950.
[5] 赵春华,樊小莉,王维莲,等.小麦候选骨干亲本科农9204遗传构成及其传递率[J].作物学报,2015,41(4):574.
ZHAO C H,FAN X L,WANG W L,etal.Genetic composition and its transmissibility analysis of wheat candidate backbone parent Kenong 9204 [J].ActaAgronomicaSinica,2015,41(4):574.
[6] 邹少奎,殷贵鸿,唐建卫,等.小麦新品种周麦23号的遗传构成分析及其特异引物筛选[J].中国农业科学,2015,48(19):3941.
ZOU S K,YIN G H,TANG J W,etal.Genetic analysis of new wheat variety Zhoumai 23 and screening of specific primers [J].ChineseAcademyofAgriculturalSciences,2015,48(19):3941.
[7] 李 俊,万洪深,杨武云,等.小麦新品种川麦104的遗传构成分析[J].中国农业科学,2014,47(12):2281.
LI J,WAN H S ,YANG W Y,etal.Dissection of genetic components in the new high yielding wheat cultivar Chuanmai 104 [J].ScientiaAgriculturaSinica,2014,47(12):2281.
[8] 黄 峰,李新平,殷贵鸿,等.豫东潮土区不同氮肥用量和基追比对周麦22号农艺性状及产量的影响研究[J].作物杂志,2011(2):75.
HUANG F,LI X P,YIN G H,etal.Studies on different nitrogen application amount and ratio of basic fertilizers to supplement fertilizers have effect on agronomic trait and yield in Eastern Henan meadow soil district [J].Crops,2011(2):75.
[9] 王立新,赵昌平,邱 军,等.记录小麦SSR带型的快捷方法[J].麦类作物学报,2006,26(4):164.
WANG L X,ZHAO C P,QIU J,etal.A new scoring method of SSR patterns for wheat [J].JournalofTriticeaeCrops,2006,26(4):164.
[10] 王立新,李云伏,常利芳,等.建立小麦品种DNA指纹的方法研究[J].作物学报,2007,33(10):1738.
WANG L X,LI Y F,CHANG L F,etal.Method of ID constitution for wheat cultivars [J].ActaAgronomicaSinica,2007,33(10):1738.
[11] 李小军,李 淦,董 娜,等.小麦优异品系遗传构成的简单重复序列(SSR)分析[J].农业生物技术学报,2012,20(3):235.
LI X J,LI G,DONG N,etal.Genetic analysis of excellent wheat breeding lines based on simple repeat sequence (SSR) markers [J].JournalofAgriculturalBiotechnology,2012,20(3):235.
[12] SOMERS D J,ISAAC P,EDWARDS K.A high-density microsatellite consensus map for bread wheat (TriticumaestivumL.)[J].TheoreticalandAppliedGenetics,2004,109 (6):1105.
[13] SPIELMEYER W,RICHARDS R A.Comparative mapping of wheat chromosome 1AS which contains the tiller inhibition gene(tin) with rice chromosome 5S [J].TheoreticalandAppliedGenetics,2004,109(6):1303.
[14] 卢 翔,张锦鹏,王化俊,等.小麦-冰草衍生后代3558-2穗部相关性状的遗传分析和QTL定位[J].植物遗传资源学报,2011,12(1):86.
LU X,ZHANG J P,WANG H J,etal.Genetic analysis and QTL mapping of wheat spike traits in a derivative line 3558-2 from wheatAgropyroncristatumoffspring [J].JournalofPlantGeneticResources,2011,12(1):86.
[15] 刘 洁.小偃麦渗入系抗条锈基因分子标记与遗传定位[D].山西大学,2013:62-70.
LIU J.Genetic analysis and molecular mapping of strip rust resistance genes in Wheat-Thinopyrunmintrogressions [D].Shanxi University,2013:62-70.
[16] D.M.TUCKER,C.A.GRIFFEY,S.LIU,etal.Confirmation of three quantitative trait loci conferring adult plant resistance to powdery mildew in two winter wheat populations[J].Euphytica,2007,155:1.
[17] T SCHNURBUSCH,S PAILLARD,A SCHORI,etal.Dissection of quantitative and durable leaf rust resistance in swiss winter wheat reveals a major resistance QTL in the Lr34 chromosomal region [J].TheoreticalandAppliedGenetics,2004,108(3):477.
[18] NEWAY C M.Grain yield and yield-related QTL validation using reciprocal recombinant inbred chromosome lines in wheat [D].University of Nebraska,2010:11-24.
[19] 贺苗苗,宋晓贺,王 阳,等.小麦-柔软滨麦草易位系M853-4抗条锈病基因的分子标记[J].植物保护学报,2010,37(2):118.
HE M M,SONG X H,WANG Y,etal.Molecular mapping of stripe rust resistance gene in wheat translocation line M853-4 derived fromLeymusmollis(Trin.) Hara [J].Actaphytophylacicasinica,2010,37(2):118.
[20] SHENG H Y,DEVEN R SEE,Timothy D Murray.Mapping QTL for resistance to eyespot of wheat inAegilopslongissima[J].TheoreticalandAppliedGenetics,2012,125(2):355.
[21] 薛 飞,王长有,张丽华,等.来自野生二粒小麦的抗白粉病基因 PmAS846及其染色体定位和分子标记分析[J].作物学报,2012,38(4):589.
XUE F,WANG C Y,ZHANG L H,etal.Chromosome location and molecular mapping of powdery mildew resistance gene PmAS846 originated from wild emmer (Triticumturgidumvar.dicoccoides) [J].ActaAgronomicaSinica,2012,38(4):589.
[22] SUN X L,LIU D,ZHANG H Q,etal.Identification and mapping of two new genes conferring resistance to powdery mildew fromAegilopstauschii(Coss.) Schmal [J].JournalofIntegrativePlantBiology,2006,48 (10) :1204.
[23] 王瑞霞,张秀英,伍玲,等.不同生态环境下冬小麦籽粒大小相关性状的 QTL 分析[J].中国农业科学,2009,42(2):398.
WANG R X,ZHANG X Y,WU L,etal.QTL analysis of grain size and related traits in winter wheat under different ecological environments[J].ScientiaAgriculturaSinica,2009,42(2):398.
[24] 刘路平.黄淮小麦新品种遗传多样性及分子标记与性状的关联分析[D].杨凌:西北农林科技大学,2014:23-41.
LIU L P.Genetic diversity of new wheat cultivars in huang-huai area and association of markers with traits [D].Yangling:North West Agriculture and Forestry University,2014:23-41.
[25] 王慧茹.小麦回交导入系抗旱相关重要性状数量位点遗传剖析[D].兰州:甘肃农业大学,2013:37-56.
WANG H R.Genetic dissection of quantitative trait loci for important traits associated with drought tolerance in wheat [D].Lanzhou:Gansu Agricultural University,2013:37-56.
[26] 徐金秋.小滨麦种质系的鉴定及抗病基因的分子标记定位[D].泰安:山东农业大学,2012:40-43.
XU J Q.Identification of tritileymus germplasm line and SSR molecular identification of its resistance gene [D].Taian:Shandong Agricultural University,2012:40-43.
[27] 李在峰.中国小麦品种条锈鉴定及抗条锈新基因YrZH84的分子标记[D].北京:中国农业科学院,2006:46-54.
LI Z F.Identification of slow stripe rust resistance in Chinese wheat and molecular mapping of new stripe rust resistance geneYrZH84[D].Beijing:Chinese Academy of Agricultural Sciences,2006:46-54.
[28] SAMBASIVAM PK,BANSALUK,HAYDEN MJ,etal.Identification of markers linked with stem rust resistance genesSr33 andSr45 [C]//Proceedings of 11th international wheat genetics symposium Sydney University Press,Sydney,2008:351.
[29] 肖永贵.山东小麦产量性状遗传进度与骨干亲本周8425B的分子解析[D].杨凌:西北农林科技大学,2011:56-77.
XIAO Y G.Genetic improvement of yield traits in Shandong wheat cultivars and molecular dissection of core parent Zhou 8425B [D].Yangling:North West Agriculture and Forestry University,2011:56-77.
[30] BOHN M,UTZ H F,MELCHINGER A E.Genetic similarities among winter wheat cultivars determined on the basis of RFLPs,AFLPs and SSRs and their use for predicting progeny variance [J].CropScience,1999,39:22.
[31] PLASCHKE J,GANAL M W,RÖDER M S.Detection of genetic diversity in closely related bread wheat using microsatellite markers [J].TheoreticalandAppliedGenetics,1995,91:1001.
[32]袁园园,王庆专,崔 法,等.小麦骨干亲本碧蚂4号的基因组特异位点及其在衍生后代中的传递[J].作物学报,2010,36(1):9.
YUANG Y Y,WANGQ Z,CUI F,etal.Specific loci in genome of wheat milestone parent Bima 4 and their transmission in derivatives[J].ActaAgronomicaSinica,2010,36(1):9.
[33] 徐 鑫,李小军,李立会.小麦系选品种与其亲本的遗传差异分析[J].麦类作物学报,2013,33(2):255.
XU X,LI X J,LI L H.Genetic difference between seven wheat accessions developed by direct selection from a parental genotype and their parents[J].JournalofTriticeaeCrops,2013,33(2):255.
[34] BERNARDO R,MUFIGNEUX A,MAISONNEUVE J P,etal.RFLP-based estimates of parental contribution to F2and BC1-derived maize inbreds [J].TheoreticalandAppliedGenetics,1997,94(5):652.
[35] BEMARDO R,ROMERO-SEVERSON J,ZIEGLE J,etal.Parental contribution and coefficient of coancestry among maize inbreds:pedigree,RFLP and SSR data [J].TheoreticalandAppliedGenetics,2000,100(3):552.
[36] SJAKSTE T G,RASHAL I,RGDER M S.Inheritance of microsatellite alleles in pedigrees of Latvian barley varieties and related European ancestors [J].TheoreticalandAppliedGenetics,2003,106(3):539.
[37] 殷贵鸿,王建武,闻伟锷,等.小麦抗条锈病基因YrZH84的RGAP标记及其应用[J].作物学报,2009,35(7):1274.
YIN G H,WANG J W,WEN W E,etal.Mapping of wheat stripe rust resistance geneYrZH84 with RGAP markers and its application [J].ActaAgronomicaSinica,2009,35(7):1274.
[38] 滕晓月,陶龙兴,孙雷心.小麦品种的蛋白电泳鉴定[J].作物学报,1988,14(4):322.
TENGX Y,TAO L X,SUN L X.Identification of wheat proteins by electrophorsis [J].ActaAgronomicaSinica,1988,14(4):322.
Molecular and Genetic Basis of Wheat Variety Zhoumai 22 and Specific Primers Screening
ZOU Shaokui,YIN Guihong,TANG Jianwei,HAN Yulin,LI Shuncheng,LI Nannan, HUANG Feng,WANG Lina,ZHANG Qian,GAO Yan
( Zhoukou Academy of Agricultural Sciences / Wheat Germplasm Improvement Engineering Research Center of Henan Province,Zhoukou,Henan 466001,China)
Common wheat variety Zhoumai 22 with the advantages of high yield,stable yield,multi-resistance and eurytopicity,is the largest cultivar in cultivated area,and is also excellent parental material.The objectives of this study were to reveal the genetic basis of Zhoumai 22, and to screen the specific primers of Zhoumai 22.A total of 340 SSR markers covering 21 wheat chromosomes were used to analyze Zhoumai 22 and its parents (Zhoumai 12,Wenmai 6,and Zhoumai 13).The results indicated that there are large differences in genetic contribution for Zhoumai 22 and its parents,of which the rate of genetic contribution from Wenmai 6 to Zhoumai 22 was 37.35%,much higher than those from Zhoumai 12 (26.51%) and Zhoumai 13 (36.14%) to Zhoumai 22.It was found that the clustering result of the genetic similarity coefficient between Zhoumai 22 and its parents does not agreed with pedigree analysis,showing that the genetic material partial separated in the process of breeding.Genetic contribution from parents to Zhoumai 22 showed large variation in different genomes,and emphasized particularly on the genomes of A,B and D.Twenty-eight specific loci were evaluated in Zhoumai 22,of which one specific SSR marker differentiated Zhoumai 22 from other varieties.This study established a simple,rapid,accurate and stable testing method for varieties authenticity of Zhoumai 22,and laid the foundation of molecular genetics for further genetic improvement and application.
Common wheat; Zhoumai 22; Molecular and Genetic; Specific primer
时间:2017-04-07
2016-05-27
2016-07-19 基金项目:国家高技术研究发展计划(“863”计划)项目(2012AA101105);国家转基因生物新品种培育重大专项(2014ZX08002003);国家农业科技成果转化资金项目(2014D00000018);河南省杰出青年基金项目(144100510004);河南省现代农业产业技术体系建设专项(2130199-ny)
E-mail:zoushaokui2015@163.com
殷贵鸿(E-mail:yinguihong2008@163.com)
S512.1;S336
A
1009-1041(2017)04-0472-11
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170407.1020.014.html
