井冈山山顶矮林优势种云锦杜鹃和猴头杜鹃叶片解剖结构比较
2017-04-26邓贤兰曹裕松
邓贤兰,刘 鑫,曹裕松,管 杰
(井冈山大学 生命科学学院,江西 吉安 343009)
井冈山山顶矮林优势种云锦杜鹃和猴头杜鹃叶片解剖结构比较
邓贤兰,刘 鑫,曹裕松,管 杰
(井冈山大学 生命科学学院,江西 吉安 343009)
以井冈山山顶矮林的优势种云锦杜鹃和猴头杜鹃为研究对象,采用石蜡切片法和光镜观察技术对其叶片解剖结构进行了研究,并探讨其与生境的适应关系,为山顶矮林的保护提供科学依据。结果表明:云锦杜鹃和猴头杜鹃叶片的上表皮由2层细胞组成,为复表皮,下表皮由1层细胞组成,下表皮细胞外周壁向外形成很多乳头状突起,上、下表皮外均具有角质膜,气孔位于下表皮;云锦杜鹃和猴头杜鹃叶肉组织分化为栅栏组织和海绵组织,为典型的异面叶,栅栏组织发达,海绵组织胞间隙发达,栅海比分别为0.995和0.637,栅栏组织和海绵组织细胞中含有星状的簇晶;中脉维管组织和机械组织发达,维管束类型为周韧维管束。这些特征表明,井冈山山顶矮林优势种云锦杜鹃和猴头杜鹃叶为典型的旱生植物叶,其结构特征与其所处的井冈山高海拔山顶严酷环境相适应。
云锦杜鹃;猴头杜鹃;山顶矮林;叶片解剖结构;优势种;井冈山
由于自然和人为因素的影响,全球生态环境问题日益突出,作为生态系统的第一生产者,植物受环境变化的影响是直接且多方面的[1],尤其是处于极端环境下的植物。叶片是植物进化过程中对环境变化较敏感且可塑性较大的器官,其结构特征最能体现环境因子对其的影响或植物对环境的适应[2-5]。山顶矮林作为中国森林生态系统的一个植被型,其所处的生态环境条件独特,生态系统比较脆弱,对环境的敏感性较强,气候的变化可能导致该生态系统退化,甚至消失[4]。因此对山顶矮林优势种叶片形态解剖结构进行研究,了解其叶片形态解剖结构对环境变化的响应与适应,是探索植物对环境变化的适应机制和制定相应保护策略的基础。
云锦杜鹃(RhododendronfortuneiLindley)和猴头杜鹃(RhododendronsimiarumHance)为杜鹃花科(Ericaceae)杜鹃属(Rhododendron)常绿小乔木或灌木,是我国的特有种[6],也是组成我国亚热带山顶矮林的优势种,对森林生态系统稳定性的维持、生态环境的保护和森林景观的形成起着不可替代的作用,同时具有重要的观赏价值[7-9]。近年来,由于人为因素的影响(如掠夺式的采挖),以及云锦杜鹃和猴头杜鹃群落所处的高山环境条件严酷(不利于种子萌发和幼苗生长),群落中2种杜鹃幼苗和幼树较少,群落处于衰退阶段[10-11],因此,有必要对其开展相关研究。
我国有关云锦杜鹃和猴头杜鹃的研究较多,主要集中在生态学[12-14]、开花特性[15]、栽培繁殖[16-17]、菌根[18]和遗传多样性[19]等方面。而有关山顶矮林优势种云锦杜鹃和猴头杜鹃叶片解剖结构的研究尚未见报道。因此,本实验以分布于江西井冈山山顶矮林的优势种云锦杜鹃和猴头杜鹃为研究材料,通过对其叶片解剖结构特征的研究、分析和比较,了解其叶片形态解剖结构对环境变化的响应与适应,探索植物对环境变化的适应对策和机制,为揭示云锦杜鹃和猴头杜鹃等山顶矮林植物对生态环境的适应对策和机制,以及预测植物对未来气候变化的响应提供科学的理论依据,为我国山顶矮林的保护提供有价值的参考。
1 研究地概况
井冈山位于江西省西南部,地处湘赣两省交界的罗霄山脉中段,地理坐标为26°27'~26°40' N、113°39'~114°23' E。整个山体呈东北—西南走向,境内山势高峻、群山层叠、峰丘相间、岭谷夹持,地形复杂。最高海拔1 841 m,最低海拔202 m;土壤以山地黄壤为主。井冈山属亚热带温暖湿润气候区,年平均气温14.2 ℃,年降雨量1 856.2 mm。境内植被类型丰富,主要有常绿阔叶林、常绿落叶阔叶混交林、针阔混交林、针叶林和山顶矮林。山顶矮林分布于井冈山海拔900~1 800 m的山脊或山顶云雾线上,是半地带性的地形顶级群落[7,11]。
2 材料与方法
2.1 研究材料
云锦杜鹃叶片采自江西省井冈山江西垇和坪水山海拔1 700~1 850 m的山顶矮林云锦杜鹃群落,猴头杜鹃叶片采自江西省井冈山笔架山海拔1 050~1 300 m的山顶矮林猴头杜鹃群落,凭证标本藏于井冈山大学生命科学学院植物标本室。在2个采集地点分别选取10株云锦杜鹃和10株猴头杜鹃植株(株间距大于100 m)进行叶片采集,在植株距地面1.5 m处随机剪取向阳面的成熟叶片6片,并在主脉中部取样,样品长宽均为1 cm。立即用FAA固定液[V(甲醛)∶V(冰乙酸)∶V(70%乙醇)=1∶1∶18]固定。
2.2 实验方法
石蜡切片制作:采取常规石蜡切片法处理叶片。即将固定好的材料经各级浓度(50%、70%、85%、95%、100% 2次)的乙醇脱水,二甲苯透明、浸蜡、包埋修整后,用旋转式切片机进行切片,切片厚度为10~15 μm,切片用二甲苯脱蜡,番红固绿染色,树胶封片[20-21]。采用蔡司光学显微成像系统进行观察、照相和测量。
观测指标的测量:利用测量软件测量上下表皮厚度、中脉厚度、叶片厚度、栅栏组织厚度、海绵组织厚度和最大导管直径等[22],每类数据均为6个切片的随机测量值,采用Microsoft Excel进行数据处理,计算叶片各指标平均数和标准差,同时观察结晶细胞情况等。
3 结果与分析
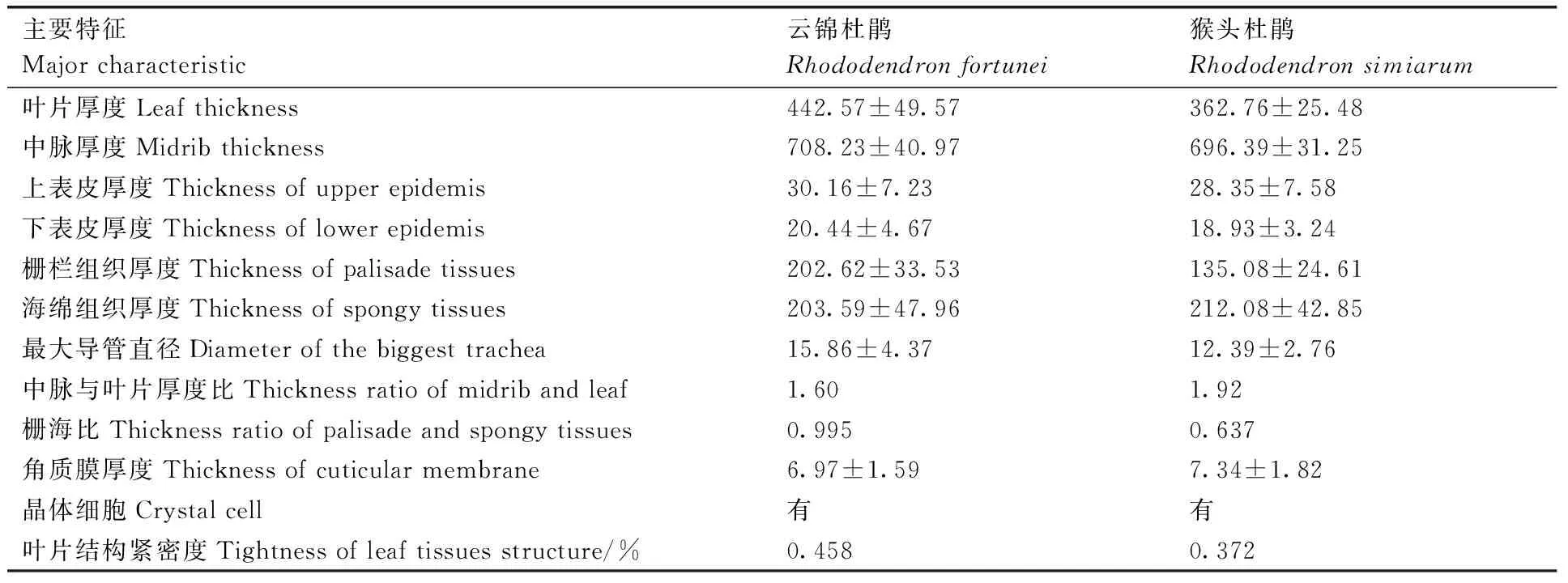
云锦杜鹃和猴头杜鹃叶片均由表皮、叶肉组织和叶脉组成,叶肉组织明显分化为栅栏组织和海绵组织,为典型的异面叶,云锦杜鹃和猴头杜鹃叶片平均厚度分别为(442.57 ± 49.57)μm和(362.76±25.48)μm,自上而下依次为上表皮、栅栏组织、海绵组织和下表皮(图1,图2,表1)。

图1 井冈山山顶矮林优势种云锦杜鹃叶片解剖结构图Fig.1 Anatomical structure of leaf blade of dominant species Rhododendron fortunei Lindley in montane elfin forest in Jinggangshan

图2 井冈山山顶矮林优势种猴头杜鹃叶片解剖结构图Fig.2 Anatomical structure of leaf blade of dominant species Rhododendron simiarum Hance in montane elfin forest in Jinggangshan
3.1 表皮
云锦杜鹃和猴头杜鹃叶片上表皮都由2层细胞组成,排列整齐紧密,细胞为不规则长方形或近椭圆形,内平周壁呈弧形略向内凸起,上表皮厚度分别为(30.16±7.23)μm和(28.35±7.58)μm(图1-D,图2-D)。上表皮外层细胞较小,细胞外平周壁被发达角质膜,角质膜厚度分别为(6.97±1.59)μm和(7.34±1.82)μm;上表皮内层细胞较大,内可见到原生质体(图1-E,图2-D)。下表皮细胞均为1层,排列紧密,细胞较小,均为不规则的方形或圆形;细胞外周壁向外形成乳头状突起,外壁被角质膜;其中,云锦杜鹃乳状突较多、较密,且连接在一起,下表皮厚度为(20.44±4.67)μm(图1-C,F),猴头杜鹃的乳头状突起较疏,下表皮厚度为(18.93±3.24)μm(图2-B,C)。猴头杜鹃的下表皮具有较密的表皮毛(图2-J)。
3.2 叶肉
表1 井冈山山顶矮林2种优势种杜鹃叶片主要解剖结构特征
Table 1 Anatomical characteristic of leaf blade of two dominant species in montane elfin forest in Jinggangshan
μm
云锦杜鹃和猴头杜鹃叶肉组织发达,均分化为栅栏组织和海绵组织。云锦杜鹃栅栏组织由2~6层长柱状细胞组成,厚度为(202.62±33.53)μm,细胞排列较为紧密,内富含叶绿体(图1-B,C,D,E);海绵组织厚度为(203.59±47.96)μm,海绵组织细胞较短,形状不规则,排列疏松,细胞间隙发达,细胞内也含有叶绿体(图1-B,C,F);其栅栏组织与海绵组织厚度比(栅海比)为0.995。猴头杜鹃栅栏组织由2~3层柱状细胞组成,厚度为(135.08±24.61)μm,细胞排列较为疏松,内含叶绿体(图2-B,C,D);海绵组织厚度为(212.08±42.85)μm,细胞形状为近圆形或不规则形,胞间隙特别发达,细胞内也含有叶绿体(图2-D,E);其栅海比为0.637。2种杜鹃栅栏组织和海绵组织细胞中均含有簇状的草酸钙结晶(图1-E,F,图2-D,E,F)。
3.3 叶脉
云锦杜鹃和猴头杜鹃叶脉发达,中脉较粗,各级侧脉逐渐变小,横切面上,中脉在叶的近轴面凹陷,在远轴面凸起,整体近半圆形(图1-A,G;图2-A,H),厚度分别为(708.23±40.97)μm和(696.39±31.25)μm,中脉与叶片厚度比分别为1.60和1.92。
云锦杜鹃叶中脉处上、下表皮细胞均较小,上表皮细胞2层,下表皮细胞1层,细胞形状为近圆形或椭圆形,排列紧密,外层细胞外均具有角质膜(图1-G,H)。上表皮下为4~5层排列紧密的厚角组织细胞,下表皮内为5~6层排列紧密的厚壁组织细胞,厚壁组织细胞与中脉维管束之间为一些薄壁细胞(图1-G,H)。猴头杜鹃叶中脉及侧脉的近轴面都有厚角组织细胞,远轴面有厚壁组织细胞,厚壁组织细胞与下表皮之间的薄壁组织细胞排列较疏松,胞间隙发达(图2-G,H)。
云锦杜鹃和猴头杜鹃叶中脉维管束较为发达,横切面观为近圆形(图1-A,G,I;图2-A,H,I)。维管束类型均为周韧维管束,圆环状的韧皮部环绕木质部排列,且远轴面一侧韧皮部比近轴面发达(图1-G,I;图2-H,I);木质部较为发达,导管分子成链状单列排列,每列导管分子6~8个,1~2列导管分子与1列木射线薄壁细胞相间排列,云锦杜鹃和猴头杜鹃最大导管直径分别为为(15.86±4.37)μm(图1-I,J)和(12.39±2.76)μm(图2-I)。
4 结论与讨论
井冈山山顶矮林优势种云锦杜鹃和猴头杜鹃叶片均为典型的异面叶,叶片较厚,上表皮为复表皮,下表皮由单层细胞构成,细胞外壁具有乳头状突起,上、下表皮细胞外壁均具有角质膜,叶肉组织分化为栅栏组织和海绵组织,叶肉组织细胞内具有草酸钙结晶,栅栏组织、叶脉机械组织和维管组织发达,这些特征表明云锦杜鹃和猴头杜鹃叶片为典型的旱生植物叶。
山顶矮林是一类分布于我国热带和亚热带山顶或山脊的特殊植被类型,紫外线辐射较强、山风大、日照少、云雾多、土层薄、霜冻、生理干旱等伴随其整个生活周期。严酷的环境条件,塑造了这类植物特殊的适应机制。本研究中,云锦杜鹃和猴头杜鹃分别生长于井冈山海拔1 700~1 850和1 050~1 300 m的山顶或山脊环境,2种植物的叶片均为异面叶,叶片较厚,具有复层上表皮和单层下表皮,栅栏组织和机械组织发达,栅海比较大,这些特点与张春影等[23]和曹晓娟等[24]对杜鹃属植物叶片解剖结构研究结果相一致。较厚的叶片具有保温和耐辐射的作用,以适应高海拔地区寒冷气候和近日照的环境条件;复层上表皮,上下表皮细胞外均具有角质膜,对增强高山植物的耐旱性和御寒性有一定作用,一方面能有效地抵御干旱、寒潮、冰霜等不良天气的侵害,防止冻伤、蒸腾失水和反渗透失水,另一方面还能增强叶表面的机械支持强度和保护功能;下表皮细胞外壁具有乳头状突起,表现为明显的高山植物形态结构和抗寒特征,能更有效地抵御低温侵袭,同时适应林下弱光和阴湿环境;叶肉组织细胞含有草酸钙结晶,结晶积累可造成细胞解体,导致气道(通气组织)的形成和扩大,从而适应高山上空气稀薄的环境。
虽然所研究的云锦杜鹃和猴头杜鹃均为井冈山山顶矮林优势种,叶片解剖结构均表现为旱生植物叶的特点,但其叶片解剖结构仍存在一些差异。首先,云锦杜鹃的叶片厚度、中脉厚度、上下表皮厚度、栅栏组织厚度、最大导管直径和栅海比均比猴头杜鹃大;其次,2种植物叶片栅栏组织和海绵组织的差异更大,云锦杜鹃叶片栅栏组织厚度和组成细胞层次比猴头杜鹃的大且多,排列更为紧密,而猴头杜鹃叶片的海绵组织厚度比云锦杜鹃的大,且细胞间隙更为发达;第三,猴头杜鹃叶片的下表皮具有较密的表皮毛,而云锦杜鹃没有,但是云锦杜鹃下表皮的乳头状突起比猴头杜鹃的下表皮多。这些差异的形成可能与2种植物所处海拔高度不同有关,云锦杜鹃所处的生境海拔比猴头杜鹃的高400~800 m,夏季光照和辐射更强,冬季更寒冷,这些更为严苛的环境条件塑造了云锦杜鹃叶片的特有结构。云锦杜鹃在井冈山主要集中分布在江西垇、坪水山和梨树洲等地海拔1 300~1 800 m的山坡和山顶,林冠整齐,长势较繁茂[7];其叶片解剖结构,如叶片的厚度、栅栏组织厚度、栅海比及叶片紧密度等是否随着海拔的升高而呈增加趋势,海拔是否是影响其叶片结构的主要因素等,有待进一步研究。
[1] 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报, 2005, 22 (S1): 118-127. LI F L, BAO W K. Responses of the morphological and anatomical structure of the plant leaf to environmental change [J].ChineseBulletinofBotany, 2005, 22 (S1): 118-127.(in Chinese with English abstract)
[2] 王勋陵, 王静. 植物的形态结构与环境[M]. 兰州: 兰州大学出版社, 1989: 105-138.
[3] 李翠, 程明, 唐宇丹, 等. 青藏高原2种柳属植物叶片解剖结构和光合特征的比较[J]. 西北植物学报, 2009, 29(2): 275-282. LI C, CHENG M, TANG Y D, et al. Characteristics comparision of leaf anatomy structure and photosynthesis of twoSalixspecies at Qinghai-Xizang Plateau [J].ActaBotanicaBoreali-OccidentaliaSinica, 2009, 29(2): 275-282. (in Chinese with English abstract)
[4] 徐捷, 王希华. 中国山顶苔藓矮曲林的分布及其特征[J]. 华东师范大学(自然科学版), 2010 (4): 44-53. XU J, WANG X H. Distribution and characteristics of the mossy dwarf forests in China [J].JournalofEastChinaNormalUniversity(NaturalScience), 2010 (4): 44-53. (in Chinese with English abstract)
[5] 姜永雷, 邓莉兰, 黄晓霞. 不同海拔川滇高山栎叶片的解剖结构特征[J]. 江苏农业科学, 2015, 43(1): 195-198. JIANG Y L, DENG L L, HUANG X X. Leaf eco-anatomical structures of Quercus aquifolioides at different altitudes[J].JiangsuAgriculturalSciences, 2015, 43(1): 195-198. (in Chinese with English abstract)
[6] FANG M Y, FANG R Z, HE M Y, et al. Flora of China [M]. Beijing: Science Press, 2005, 14: 260-455.
[7] 林英. 井冈山自然保护区考察研究[M]. 北京: 新华出版社, 1990: 140-198.
[8] 叶居新. 中国的猴头杜鹃矮林[J]. 武汉植物学研究, 1994, 12(2): 170-174. YE J X. On theRhododendronsimiarumelfin forest of China [J].JournalofWuhanBotanicalResearch, 1994, 12(2): 170-174. (in Chinese with English abstract)
[9] 邓贤兰, 刘鹏, 吴杨, 等. 井冈山云锦杜鹃群落特征研究 [J]. 亚热带植物科学, 2011, 40(4): 1-6. DENG X L, LLU P, WU Y, et al. Studies on the community characteristics ofRhododendronfortuneiin Jinggang Mountain [J].SubtropicalPlantScience, 2011, 40(4): 1-6. (in Chinese with English abstract)
[10] 赵丽娟, 李家湘, 邓家兴. 湖南平江幕阜山云锦杜鹃群落特征的分析[J]. 中南林业科技大学学报, 2005, 25(2): 81-84. ZHAO L J, LI J X, DENG J X. Primary studies ofRhododendronfortuneicommunity of Mufu Forest Park, Pingjiang, Hunan [J].JournalofCentralSouthForestryUniversity, 2005, 25(2): 81-84. (in Chinese with English abstract)
[11] 邓贤兰, 吴杨, 刘玉成, 等. 井冈山猴头杜鹃群落特征的研究[J]. 生态环境学报, 2011, 20(10): 1430-1435. DENG X L, WU Y, LIU Y C, et al. Studies on the community characteristics ofRhododendronsimiarumin Jinggang Mountain [J].EcologyandEnvironmentalSciences, 2011, 20(10): 1430-1435. (in Chinese with English abstract)
[12] 王厚麟, 黄林生, 缪绅裕, 等. 广东石门台保护区猴头杜鹃群落特征[J]. 广西植物, 2008, 28(1): 73-77. WANG H L, HUANG L S, MIU S Y, et al. Community characteristics ofRhododendronsimiarumin Shimentai Nature Reserve, Guangdong [J].Guihaia, 2008, 28(1): 73-77. (in Chinese with English abstract)
[13] 何小娟, 何东进, 洪伟, 等. 天宝岩天然猴头杜鹃种群空间分布格局研究[J]. 云南农业大学学报, 2009, 24(5): 734-738. HE X J, HE D J, HONG W, et al. Study on spatial distribution of naturalRhododendronsimiarumpopulations in Tianbaoyan Nature Reserve [J].JournalofYunnanAgriculturalUniversity, 2009, 24(5): 734-738. (in Chinese with English abstract)
[14] 黄川腾, 唐光大, 刘乐, 等. 广东天井山云锦杜鹃种群及其所处群落特征[J]. 西南林学院学报, 2010, 30(6): 15-19. HUANG C T, TANG G D, LIU L, et al. Population and community characteristics ofRhododendronfortuneiat Mount Tianjing of Guangdong Province [J].JournalofSouthwestForestryUniversity, 2010, 30(6): 15-19. (in Chinese with English abstract)
[15] 边才苗, 金则新. 天台山云锦杜鹃的开花与结实特性[J]. 园艺学报, 2006, 33(1): 101-104. BIAN C M, JIN Z X. The flowering and fruit set features ofRhododendronfortuneiin Tiantai Mountains [J].ActaHorticulturaeSinica, 2006, 33(1): 101-104. (in Chinese with English abstract)
[16] 边才苗, 金则新, 张俊会, 等. 云锦杜鹃种子萌发对干旱胁迫的响应[J]. 植物研究, 2006, 26(6): 718-726. BIAN C M, JIN Z X, ZHANG J H, et al. Response of seed germination ofRhododendronfortuneito drought stress[J].BulletinofBotanicalResearch, 2006, 26(6): 718-726. (in Chinese with English abstract)
[17] 朱春艳, 李志炎, 鲍淳松, 等. 云锦杜鹃组培快繁技术研究[J]. 中国农学通报, 2006, 22(5): 335-337. ZHU C Y, LI Z Y, BAO C S, et al.Invitrorapid micropropagation ofRhododendronfortunei[J].ChineseAgriculturalScienceBulletin, 2006, 22(5): 335-337. (in Chinese with English abstract)
[18] 张春英, 侯奕敏, 戴思兰. 云锦杜鹃菌根显微结构观察[J]. 园艺学报, 2008, 35(11): 1641-1646. ZHANG C Y, HOU Y M, DAI S L. Observationon microstructure of mycorrhizal root ofRhododendronfortuneiL [J].ActaHorticulturaeSinica, 2008, 35(11): 1641-1646. (in Chinese with English abstract)
[19] 金则新, 李钧敏, 顾奇萍. 云锦杜鹃自然居群遗传多样性的ISSR分析[J]. 园艺学报, 2006, 33(6): 1263-1267. JIN Z X, LI J M, GU Q P. Genetic diversity in the natural populations ofRhododendronfortuneirevealed by ISSR molecular markers [J].ActaHorticulturaeSinica, 2006, 33(6): 1263-1267. (in Chinese with English abstract)
[20] 王英典, 刘宁, 刘全儒, 等. 植物生物学实验指导 [M]. 2版. 北京: 高等教育出版社, 2011.
[21] 李正理. 植物制片技术[M]. 北京: 科学出版社, 1987.
[22] 容丽, 陈训, 汪小春. 百里杜鹃杜鹃属13种植物叶片解剖结构的生态适应性[J]. 安徽农业科学, 2009, 37(3): 1084-1088. RONG L, CHEN X, WANG X C. Leaf anatomical characters and its ecological adaptation of 13 Species ofRhododendronin Baili Azalea Area [J].JournalofAnhuiAgriculture.Science, 2009, 37(3): 1084-1088. (in Chinese with English abstract)
[23] 张春影, 王艳, 李美善, 等. 长白山不同海拔牛皮杜鹃叶片解剖结构的比较分析[J]. 延边大学农学学报, 2014, 36(1): 39-43. ZHANG C Y, WANG Y, LI M S, et al. Comparative analysis of anatomical structure ofRhododendronchrysanthumleaves at different altitudes in Changbai Mountain area [J].JournalofAgriculturalScienceYanbianUniversity, 2014, 36(1): 39-43. (in Chinese with English abstract)
[24] 曹晓娟, 刘建军, 杨梅. 太白山5种杜鹃属植物叶片光合特性及解剖结构的生态适应性研究[J]. 西北植物学报, 2009, 29(12): 2483-2491. CAO X J, LIU J J, YANG M. Photosynthetic characteristics and anatomical structure of five species ofRhododendronin the Taibai Mountain [J].ActaBotanicaBoreali-OccidentaliaSinica, 2009, 29(12): 2483-2491. (in Chinese with English abstract)
(责任编辑 侯春晓)
Comparison of leaf anatomical structure betweenRhododendronfrotuneiandRhododendronsimiarum, two dominant species of montane elfin forest in Jinggangshan
DENG Xianlan, LIU Xin, CAO Yusong, GUAN Jie
(CollegeofLifeSciences,JinggangshanUniversity,Ji’an343009,China)
TakingRhododendronfortuneiLindley andRhododendronsimiarumHance, two dominant species of montane elfin forest in Jinggangshan as study objects, the leaf anatomical structures were studied by paraffin-sectioning and optical microtechnique, and the adaptation relationship between its leaf anatomical structures and habitat were explored. The aim was to provide scientific theory foundation for the protection of montane elfin forest. The results showed that the upper epidermis consisted of two layers of epidermic cells, belonging to multiple epidermis. The lower epidermis consisted of one layer of epidermic cells, the peripheral wall of lower epidermis cell had formed outward a lot of convex papillate. The upper and lower epidermis were covered cuticular membrane. The stomata existed in lower epidermis. The mesophyll tissue ofRhododendronfortuneiLindley andRhododendronsimiarumHance were differentiated into palisade tissue and spongy tissue, which indicated that the leaf belonged to typical bifacial leaf. The palisade tissue was well developed; The cells of the spongy tissue lined up loosely and had the bigger cell interval. The ratios of the palisade tissue to the spongy tissue were 0.995 and 0.637, respectively. The palisade and spongy tissue cells contained the star clustered crystal. The vascular tissue and mechanical tissue of midrib were well developed, and the middle vein vascular bundle belonged to the amphicribral bundle. These characteristics indicated that the leaves ofRhododendronfortuneiLindley andRhododendronsimiarumHance of montane elfin forest in Jinggangshan were typical xerophil leaves. Their leaf structural characteristics were adapted to the higher elevation and the harsh environment of the top of the mountain in Jinggangshan.
Rhododendronfortunei;Rhododendronsimiarum; montane elfin forest; leaf anatomical structure; dominant species; Jinggangshan
http://www.zjnyxb.cn
10.3969/j.issn.1004-1524.2017.04.10
2016-10-10
国家自然科学基金项目(31660053);江西省教育厅科技项目(GJJ150784);吉安市科技支撑项目(吉市科计字[2014]36号16);大学生创新训练项目
邓贤兰(1969—),女,江西遂川人,博士,副教授,主要从事植物分类、植物生态及谱系地理学研究。E-mail: dengxianland@163.com
S685.21
A
1004-1524(2017)04-0583-07
浙江农业学报ActaAgriculturaeZhejiangensis, 2017,29(4): 583-589
邓贤兰,刘鑫,曹裕松, 等. 井冈山山顶矮林优势种云锦杜鹃和猴头杜鹃叶片解剖结构比较[J]. 浙江农业学报, 2017, 29(4): 583-589.
