NADPH氧化酶NOX1、NOX2和DUOX2在溃疡性结肠炎中的表达分析
2017-04-19轩青霞戴发亮冯丹丹金建军
轩青霞 戴发亮 陈 攀 冯丹丹 金建军 高 强,2*
(1.河南科技大学临床医学院,河南科技大学第一附属医院消化内科,河南洛阳 471023; 2.首都医科大学附属北京康复医院消化内科, 北京 100144)
·基础研究 ·
NADPH氧化酶NOX1、NOX2和DUOX2在溃疡性结肠炎中的表达分析
轩青霞1戴发亮1陈 攀1冯丹丹1金建军1高 强1,2*
(1.河南科技大学临床医学院,河南科技大学第一附属医院消化内科,河南洛阳 471023; 2.首都医科大学附属北京康复医院消化内科, 北京 100144)
目的 探讨NADPH氧化酶家族(nicotinamide adenine dinucleotide phosphate oxidases, NOXs)的主要成员NOX1、NOX2和双氧化物酶(dual oxidase, DUOX)2 (人:NOX1、NOX2和DUOX2;小鼠:Nox1、Nox2 和Duox2)在人炎症性肠病和小鼠肠炎模型结肠中的表达。方法 人结肠标本选自于通过结肠镜活检或手术切除的炎症性肠病组织及距离病变组织边缘5 cm以上正常组织; 同时选用8~10周龄的C57BL/6雌性小鼠建立结肠炎模型,采用掷硬币法将其随机分为对照组和慢性肠炎组,慢性肠炎组先自由饮用1.0%(质量分数)葡聚糖硫酸钠7 d,然后自由饮水14 d,循环3次;通过体质量变化和HE染色方法评估肠道炎性反应程度。采用实时定量PCR技术和免疫组织化学方法分别检测结肠组织中NOX1、NOX2及DUOX2 mRNA和蛋白表达情况,并分析在人炎症性肠病中三者表达情况与临床特征之间的关系。结果 NOX1、NOX2和DUOX2 mRNA在人炎症性肠病结肠组织中的表达量均显著高于自身正常对照组织,分别增高2.8、2.0和3.3倍(P均<0.01);与人不同,Nox1和Duox2 mRNA在小鼠慢性肠炎结肠组织中的表达量差异无统计学意义,Nox2 mRNA增加2.4倍 (P<0.01)。Nox1蛋白在正常结肠上皮细胞刷状缘和胞质中表达;Nox2蛋白主要表达于浸润的吞噬细胞和中性粒细胞;Duox2蛋白在正常结肠黏膜组织中低表达;三者在人炎症性肠病结肠组织中表达均上调(均P<0.01);在小鼠慢性肠炎模型中,Nox2和Duox2蛋白上调(均P<0.01),而Nox1无变化。除了NOX2 mRNA在男性病人表达高于女性病人外,未发现这些氧化酶与病人临床特征之间有关联。结论 NOX1、NOX2和DUOX2不但在维持机体正常生理功能过程中发挥重要作用,而且在炎症性肠病初期阶段的发病中也起一定的作用。
NADPH氧化酶;NOX1;NOX2;DUOX2;炎症性肠病
炎症性肠病 (inflammatory bowel disease, IBD) 是一种慢性非特异性肠道炎性反应性疾病,包括溃疡性结肠炎 (ulcerative colitis, UC) 和克罗恩病 (Crohn’s disease, CD)。目前其发病机制及病因尚未明确,多认为是免疫、肠道菌群、遗传和环境等多因素相互作用的结果[1]。在炎症性肠病发生发展过程中,多种因素可导致易感个体肠道内环境破坏,引起肠道黏膜免疫功能失调,从而造成黏膜屏障的损伤[2]。氧化应激 (oxidative stress) 被认为是导致肠道损伤的关键因素[3-4]。氧化应激是指机体内活性氧类 (reactive oxygen species, ROS) 和活性氮类 (reactive nitrogen species, RNS) 产生过多,超出机体对氧化产物的清除能力,进而导致组织细胞损伤的病理生理过程[4]。NADPH氧化酶 (nicotinamide adenine dinucleotide phosphate oxidases, NOXs) 是肠道黏膜ROS的重要来源,正常生理情况下ROS在肠道抵御病原微生物中发挥重要作用[5-6]。作为NADPH氧化酶家族的成员,NOX1、NOX2和双氧化物酶 (dual oxidase, DUOX)2(人:NOX1、NOX2和DUOX2;小鼠:Nox1、Nox2和Duox2)均能产生ROS,其中DUOX2可产生H2O2[7]。NOX1和 DUOX2在消化道黏膜上皮表达,而NOX2主要在巨噬细胞和中性粒细胞中表达[8]。研究[9]显示NADPH氧化酶在IBD疾病发生和发展过程中发挥作用,但国内外研究有限。本研究通过检测溃疡性结肠炎病人结肠组织和与溃疡性结肠炎相似的小鼠葡聚糖硫酸钠 (dextran sulfate sodium,DSS) 慢性结肠炎模型结肠中NOX1、NOX2和DUOX2的表达情况,探讨NADPH氧化酶NOX1、NOX2和DUOX2在IBD发生和发展中的作用,尤其是小鼠慢性结肠炎模型结肠中NADPH氧化酶的研究目前国内外尚未见报道。
1 材料与方法
1.1 材料
1.1.1 临床资料
取自28例2013年11月至2015年11月在河南科技大学第一附属医院就诊的复发的活动期溃疡性结肠炎病人,其中,男性病人18例,女性病人10例,年龄16~76岁,平均年龄 (46.5±14.7) 岁;病变部位位于结肠12例,直肠16例;活检伴不典型增生者21例,不伴不典型增生者7例。
1.1.2 实验动物
选取健康清洁级8~10周龄(体质量20~23 g)C57BL/6雌性小鼠[购于北京维通利华公司,实验动物许可证号:SYKK(豫)2016-0001],饲养于河南科技大学第一附属医院动物实验中心,室内温度保持 20 ℃~22 ℃,湿度50%左右,明暗交替周期12 h。各组小鼠均给予Co60辐照灭菌混合配方颗粒饲料(北京华阜康生物科技股份有限公司)饲养。
1.1.3 主要试剂
DSS(相对分子质量为36 000~50 000)购于美国MP Biomedicals公司。总RNA提取试剂TRIzol Reagent产于美国Invitrogen公司,反转录和实时定量PCR试剂盒购于日本Takara公司,引物由生工生物工程(上海)有限公司合成(表1)。免疫组织化学检测用NOX1和Nox1一抗分别购于武汉博士德生物公司和英国Abcam公司;NOX2/Nox2一抗购于美国Santa Cruz 公司;DUOX2和Duox2一抗分别购于北京博奥森和美国Gene Tex公司;SABC免疫组织化学检测试剂盒购于武汉博士德生物公司;浓缩型DAB显色试剂盒购于北京索莱宝公司。
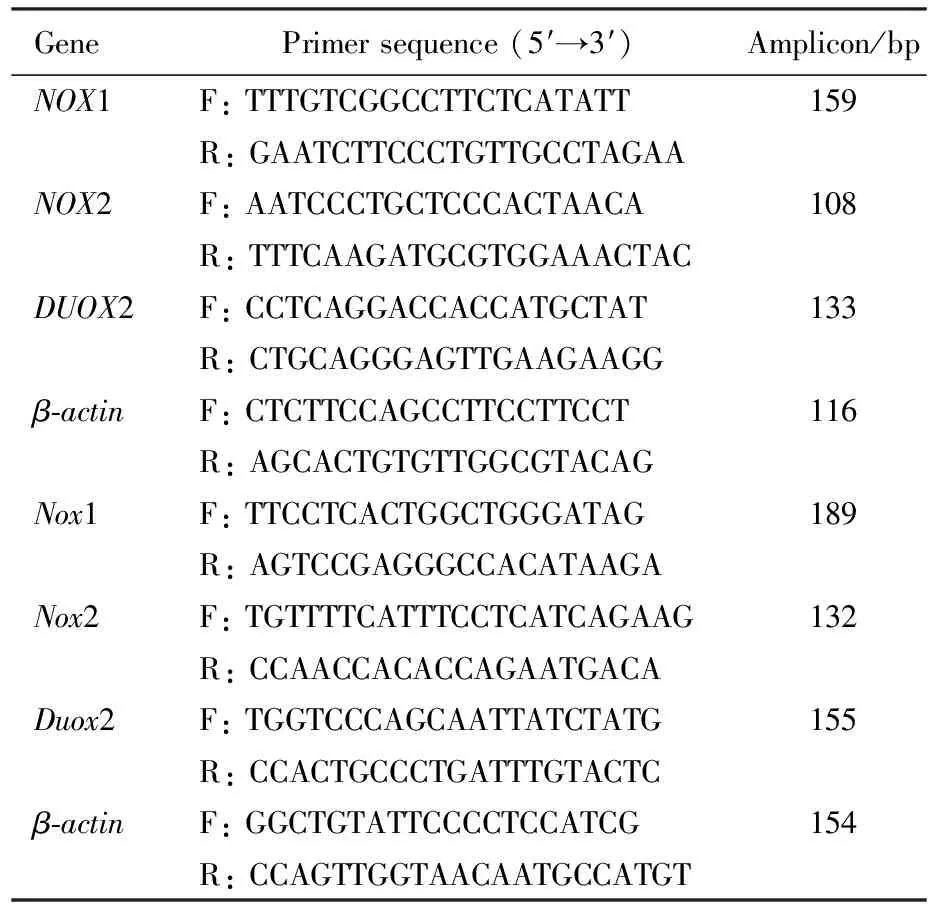
表1 引物序列
F:forward; R:reverse.
1.2 方法
1.2.1 临床取材
搜集结肠镜活检或手术切除并经病理证实的病变结肠组织标本与距病变组织边缘5 cm以上的正常组织,预冷的PBS清洗,一部分组织甲醛固定,石蜡包埋,4μm厚度切片,备免疫组织化学使用,剩余的样本在-80 ℃储存,行实时定量PCR测定。
1.2.2 动物实验设计与取材
将小鼠以个体为单位采用掷硬币的方法(正、反面分别代表实验组和对照组)随机分配到对照组和实验组(DSS组),每组8只,适应性喂养1周,对照组自由饮水;慢性肠炎组先给予含1.0%(质量分数)DSS饮用水自由饮用7 d,然后给予饮用水自由饮用14 d,循环3次。于造模第9周末脱颈椎处死所有小鼠,剖腹取结肠,PBS漂洗,自远端结肠始取约0.5 cm,甲醛液固定,经脱水后石蜡包埋,4μm厚度切片,备免疫组织化学及HE染色使用。再取1.0 cm加入RNA later液,4 ℃放置24 h后转移至-80 ℃保存,行实时定量PCR。
1.2.3 实时定量PCR检测人和小鼠NOX1、NOX2和DUOX2 mRNA表达水平
取200 mg结肠组织,放到无RNase的1.5 mL离心管中,采用Trizol法提取总RNA,用核酸定量仪测定RNA纯度和浓度。取总RNA 2μg,按照反转录试剂盒说明书反转录合成cDNA,-20 ℃保存备用。冰上配制PCR反应液,反应体系25μL:SYBR Premix EX Taq Ⅱ (2×) 12.5μL,DEPC水8.5μL,cDNA 2μL,上下游引物各1μL,于BIO-RAD Real-time PCR仪中进行PCR反应,反应条件为: 95 ℃预变性30 s,95 ℃ 5 s、59 ℃ 30 s、95 ℃ 15 s,共40个循环。结果以阈循环 (Ct) 值表示,β-actin作为内参基因,每个样本3个复孔,各样本的待测基因扩增量与β-actin基因扩增量采用Log10 (2-ΔΔCt) 法分析mRNA相对表达量。平均Ct值超过35,视为该目标基因无表达。
1.2.4 免疫组织化学方法检测人和小鼠NOX1、NOX2和DUOX2的蛋白表达
免疫组织化学染色NOX1、NOX2和DUOX2一抗工作浓度分别为1∶100、1∶200和1∶300; Nox1、Nox2和Duox2一抗工作浓度分别为1∶250、1∶200和1∶300,阴性对照组均使用PBS溶液代替一抗。在光学显微镜下进行细胞计数,每张切片随机选择10个视野,每个视野观察100个细胞。染色细胞比率评分标准:<5%计0分,5%~25%计1分,26%~50%计2分,51%~75%计3分,76%~100%计4分;染色强度评分标准:细胞无染色计0分,淡黄色计1分,棕色计2分,棕褐色计3分,根据两项评分乘积进行免疫组织化学染色评分。
1.3 统计学方法
2 结果
2.1 人结肠组织中NOX1、NOX2和DUOX2 mRNA和蛋白的表达
与正常组织相比,NOX1、NOX2与DUOX2 mRNA在炎性反应组织的表达均显著升高。炎症性肠病组NOX1、NOX2与DUOX2 mRNA的表达量分别是正常组织中的2.8、2.0和3.3倍(P均<0.01)(图1)。NOX1蛋白在正常结肠上皮细胞刷状缘和胞质表达;NOX2蛋白主要表达于浸润的吞噬细胞和中性粒细胞;DUOX2蛋白在正常结肠黏膜组织中低表达;这3种蛋白在炎症性肠病结肠组织中的表达较正常对照组均显著升高(P均<0.01),NOX2蛋白炎性反应区域可见炎性反应样细胞呈阳性染色;NOX1和DUOX2在上皮细胞刷状缘和胞质表达明显增加(表2和图2)。
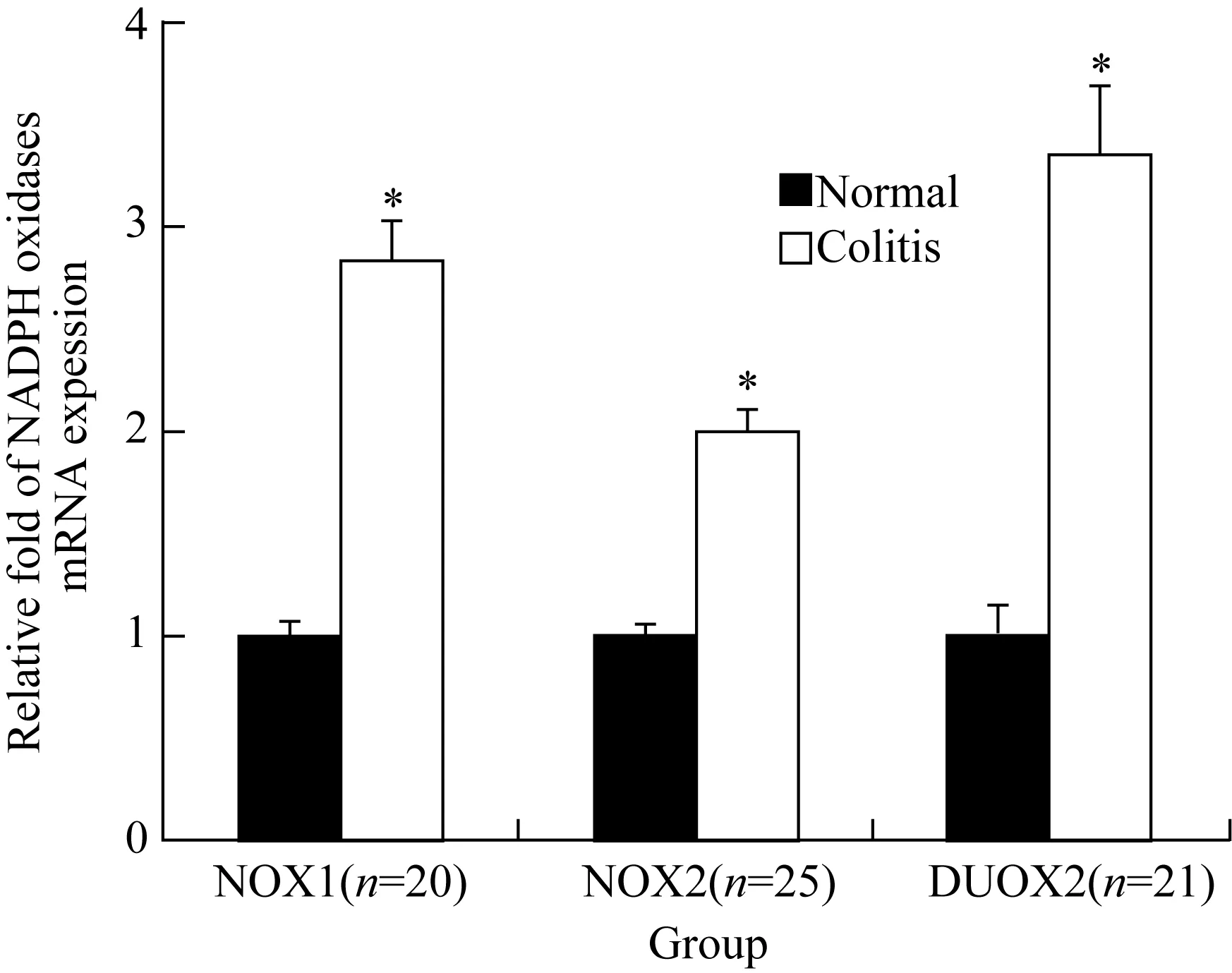
图1 炎症性肠病结肠炎性反应组织与相应正常结肠组织中NOX1、NOX2和DUOX2 mRNA的比较
*P<0.01vsnormal;NADPH:nicotinamide adenine dinucleotide phosphate.
2.2 结肠炎性反应组织中NOX1、NOX2与DUOX2 mRNA和蛋白表达与临床病理特征的关系
比较不同病人临床病理特征与NOX1、NOX2及DUOX2 mRNA和蛋白表达量之间的关系,发现除结肠炎组织中NOX2 mRNA表达水平的升高与性别相关,男性病人NOX2 mRNA表达水平较女性高外 (P<0.05),而其他临床特征如年龄、病程、病变部位以及吸烟饮酒史等与这3个基因的表达无相关性;发现NOX1、NOX2与DUOX2蛋白的表达与病人临床病理特征之间没有显著的相关性。

表2 结肠组织中NOX1,NOX2和 DUOX2蛋白表达
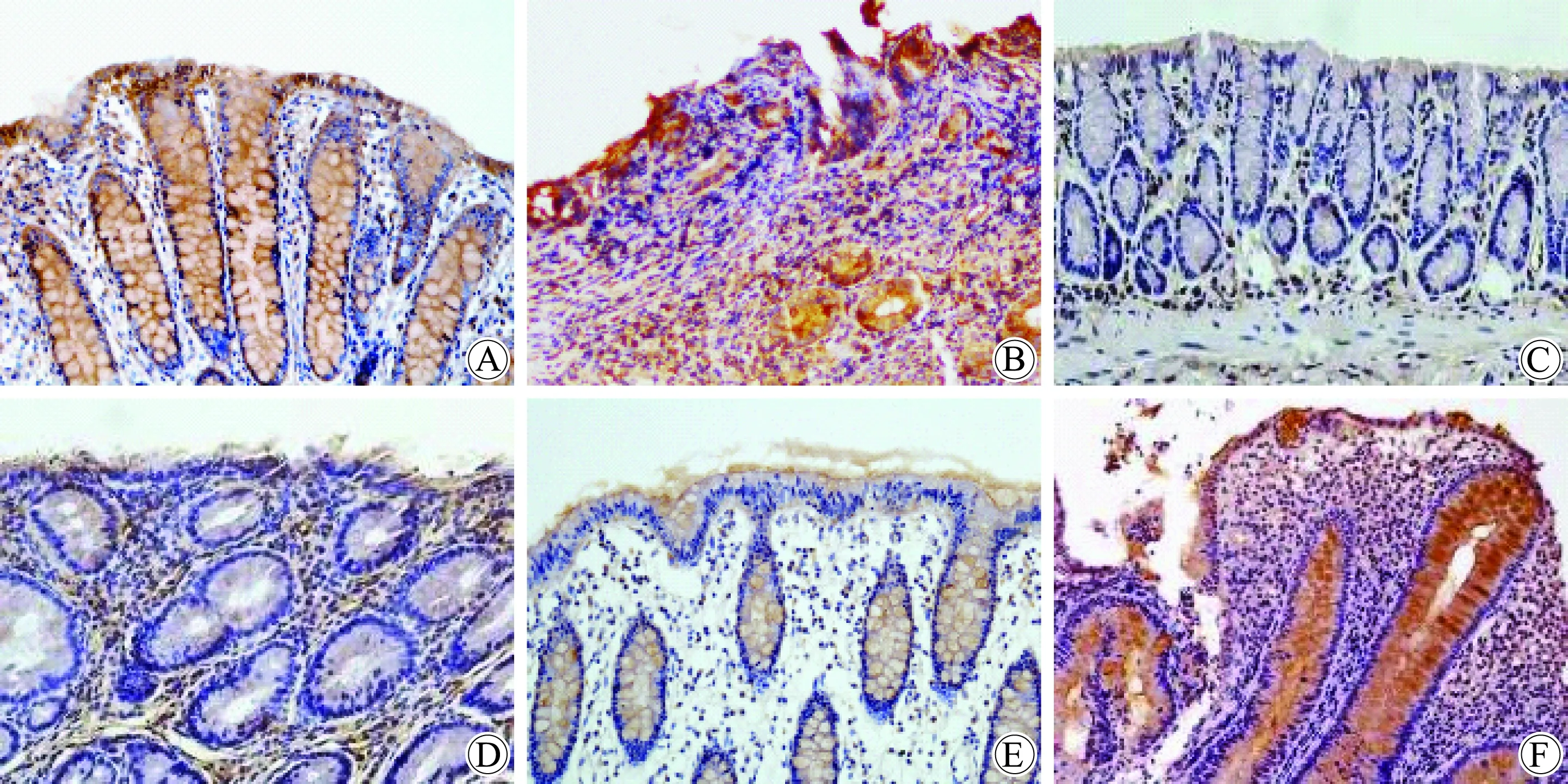
图2 结肠组织中NOX1、NOX2和DUOX2 蛋白的表达
In colitis tissue (B) NOX1 protein expression was significantly increased than normal tissue (A); NOX2 protein expression significantly increased in colitis tissue (D) than normal tissue (C), most expressed in the cytoplasm of inflammatory cell;DUOX2 protein had a little expression in brush border of epithelial cells of normal mucosa (E), the expression of DUOX2 was significantly increased in the cytoplasm of epithelial cells of colitis tissue (F).
2.3 动物实验结果
2.3.1 小鼠一般情况和体质量变化
对照组小鼠摄食饮水正常,反应机警,生长发育良好,体质量增加;慢性肠炎组于第7 d开始饮用1.0%(质量分数)DSS,饮用6 d出现懒动,摄食饮水减少,体质量下降,稀便、肛周潮湿现象,部分小鼠出现肉眼血便,停止饮用1.0%(质量分数)DSS后的第4 d体质量开始回升,在第1个循环周期中平均体质量下降近10%,在第2、3循环中,上述症状反复出现,慢性肠炎组小鼠体质量自第2周起至造模结束均低于对照组 (P<0.05)(图3)。造模结束时,两组小鼠均无死亡。
2.3.2 组织病理学评分
对照组可见小鼠结肠黏膜腺体结构完整,无炎性反应表现;慢性肠炎组小鼠结肠黏膜呈慢性炎性反应改变,腺体结构不完整,隐窝结构遭到破坏,细胞结构排列紊乱,杯状细胞减少或消失,局部伴有炎性反应细胞浸润,并可见隐窝不规则增生;两组相比炎性反应程度差异有统计学意义(P<0.01)(表3)。

图3 两组小鼠体质量变化
*P<0.05vscontrol.
2.3.3 小鼠结肠组织中Nox1、Nox2和Duox2 mRNA的表达
Nox1和Duox2 mRNA在对照组和慢性肠炎组结肠组织中没有差别,而Nox2 mRNA在慢性肠炎组结肠组织的表达较对照组升高2.4倍 (P<0.01)(图4)。
2.3.4 小鼠结肠组织中Nox1、Nox2和Duox2蛋白的表达
Nox1蛋白在正常结肠组织中表达,且主要表达在上皮细胞刷状缘和胞质;Nox2蛋白主要表达在吞噬细胞和中性粒细胞胞质中;Duox2蛋白在正常的上皮细胞刷状缘少量表达。Nox1蛋白在对照组和慢性肠炎组结肠组织中无差别,而Nox2和Duox2在慢性肠炎组结肠组织的表达较对照组均显著升高 (P均<0.01)(表3和图5)。

图4 小鼠结肠组织中Nox1、Nox2和Duox2 mRNA表达
*P<0.05vscontrol;NADPH:nicotinamide adenine dinucleotide phosphate.
3 讨论
IBD是西方国家的常见病,由于环境和饮食习惯的变化,我国IBD的总报道病例数量在近十余年内增加了2.5倍,尤其是CD增加了15.7倍[10-11]。NOX1、NOX2和DUOX2作为ROS的重要来源,正常情况下,NOX家族氧化酶产生的ROS参与维持细胞的正常生理活动,但当机体受到各种因素刺激时,引发NOX家族氧化酶过表达产生大量ROS,过多的ROS可激活核因子介导产生大量的细胞促炎因子,促进炎性反应[12]。但有关NOX1、NOX2和DUOX2在IBD疾病发生和发展过程中的研究仍有限。

表3 两组小鼠组织病理学和免疫组化评分
HE staining showed that mucous membrane structure of control group was normal, the epithelial cells were unbroken (A); Mucous membrane structure of colitis group was broken, and has inflammatory cell infiltration (B); Immumohistochemical staining showed that Nox1 expression was undifferentiated between chronic colitis group (D) and control group (C); Nox2 expression in chronic colitis group (F) was significantly higher than control group (E); Duox2 expression in chronic colitis group (H) was also higher than control group (G).
本研究结果显示,NOX1蛋白和mRNA在正常的结肠黏膜组织上皮细胞中表达,在炎性反应结肠组织中,NOX1表达明显上调,提示NOX1除维持正常生理功能过程外,还在IBD病理过程中发挥重要作用,可能参与早期阶段的炎症性肠病的发生。生理状态下NOX1源性ROS参与调节结肠上皮细胞生长和转化,并维持杯状细胞和吸收细胞功能的稳态[13]。当NOX1过度表达,增高的NOX1产生大量的ROS可以激活多种转录因子,活化的转录因子又能促进细胞因子类基因的转录,而细胞因子的转录又会进一步加剧炎性反应[14-15]。研究[16-18]表明Nox1参与细胞的增生、迁移和凋亡,并调控结肠上皮细胞的修复和宿主免疫防御功能,且与早期发生的炎症性肠病有关。本研究中NOX1在人类炎症性肠病的肠炎上皮细胞中表达,这与Szanto等[19]的研究结果一致;而慢性肠炎动物模型结果显示,Nox1 mRNA和蛋白在慢性肠炎和对照组结肠组织中的表达差异无统计学意义,可能是由于Nox1主要在肠黏膜上皮表达,慢性炎性反应时肠黏膜上皮结构破坏严重,上皮细胞大量丢失所致。
本研究结果显示NOX2 mRNA和蛋白在人和小鼠结肠黏膜炎性反应组织中的表达量较对照组明显增高,这与炎性反应组织中炎性细胞增多有关,提示在IBD疾病的发生发展中,NOX2可能在宿主免疫中发挥重要作用。静息状态时,NOX2定位在细胞内,不产生ROS,当受到外界刺激,如:病原微生物和细胞因子等,NOX2可易位到细胞表面并被迅速激活,产生大量ROS;过量的ROS可与细胞大分子发生反应,使DNA、蛋白质和脂质受到严重的氧化损伤,从而使肠上皮细胞结构与功能受损最终可导致细胞坏死,甚至死亡;此外,细胞内产生的ROS还可以通过信号转导通路激活NOX2[20]。NOX2不仅在抵御细菌入侵中起重要作用,而且还参与炎性反应的许多信号通路,如炎性反应因子反应、中性粒细胞的聚集和转录[21]。研究[22-23]显示NOX2功能受损与小于6岁的IBD的发生发展有关。
DUOX2是肠道黏膜H2O2的主要产生者,在整个消化系均有表达[24-25],生理情况下,H2O2可抵御细菌侵入和调控肠道菌群,参与肠道黏膜固有免疫应答[26]。但H2O2产生过量时可导致氧化应激,引起组织损伤[3,27-28]。对GPX1/2双敲小鼠的研究[22,29]显示Duox2可能是结肠炎的候选易感基因,在小鼠和人炎性反应性肠病中均发现DUOX2和IBD有密切的联系,并参与初期炎性反应性肠病的发生。本临床研究结果显示DUOX2 mRNA和蛋白在正常结肠组织中有一定量的表达,在炎性反应性结肠黏膜组织中的表达量明显升高;慢性肠炎动物模型结果显示,慢性肠炎时Duox2蛋白表达水平均上调,mRNA上调不明显,提示DUOX2除维持结肠组织正常生理功能外,可能在炎性反应过程中也发挥作用。
总之,本研究结果显示IBD病人病变组织中NOX1、NOX2和DUOX2基因或/和表达明显高于对照组;并首次在慢性结肠炎动物模型中得到相似的结论,提示这些NADPH氧化酶不但在维持机体正常生理功能过程中发挥重要作用,而且在炎症性肠病初期阶段的发病中也起一定的作用,其具体机制有待进一步研究。
[1] Kaser A, Zeissig S, Blumberg R S. Inflammatory bowel disease[J]. Ann Rev Immunol,2010,28: 573-621.
[2] Maloy K J, Powrie F. Intestinal homeostasis and its breakdown in inflammatory bowel disease[J]. Nature,2011, 474(7351): 298-306.
[3] Biasi F, Leonarduzzi G, Oteiza P I, et al. Inflammatory bowel disease: mechanisms, redox considerations, and therapeutic targets[J]. Antioxid Redox Signal,2013, 19(14): 1711-1747.
[4] Circu M L, Aw T Y. Intestinal redox biology and oxidative stress[J]. Semin Cell Dev Biol, 2012, 23(7): 729-737.
[5] Rada B, Leto T L. Oxidative innate immune defenses by Nox/Duox family NADPH oxidases[J]. Contrib Microbiol,2008, 15: 164-187.
[6] Bhattacharyya A, Chattopadhyay R, Mitra S, et al. Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases[J]. Physiol Rev,2014, 94(2): 329-354.
[7] Cifuentes-Pagano E, Csanyi G, Pagano P J. NADPH oxidase inhibitors: a decade of discovery from Nox2ds to HTS[J]. Cell Mol Life Sci: CMLS,2012, 69(14): 2315-2325.
[8] Katsuyama M, Matsuno K, Yabe-Nishimura C. Physiological roles of NOX/NADPH oxidase, the superoxide-generating enzyme[J]. J Clin Biochem Nutr, 2012, 50(1): 9-22.
[9] Lam G, Apostolopoulos V, Zulli A, et al. NADPH oxidases and inflammatory bowel disease[J]. Curr Med Chem,2015, 22(17): 2100-2109.
[10]Ouyang Q, Xue L Y. Inflammatory bowel disease in the 21(st) century in China: turning challenges into opportunities[J]. J Dig Dis,2012,13(4): 195-199.
[11]Jemal A, Bray F, Center M M, et al. Global cancer statistics[J]. CA: Cancer J Clin, 2011, 61(2): 69-90.
[12]Bedard K, Krause K H. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology[J]. Physiol Rev,2007, 87(1): 245-313.
[13]韩晓燕, 高丽萍, 刘箐. NOX家族蛋白的研究进展[J]. 生命科学, 2012,24(6): 568-577.
[14]Reuter S, Gupta S C, Chaturvedi M M, et al. Oxidative stress, inflammation, and cancer: how are they linked? [J]. Free Radic Biol Med,2010, 49(11): 1603-1616.
[15]Jones R M, Luo L, Ardita C S, et al. Symbiotic lactobacilli stimulate gut epithelial proliferation via Nox-mediated generation of reactive oxygen species[J]. EMBO J,2013, 32(23): 3017-3028.
[16]杨茉莉, 李宏谦, 李小珍, 等. NADPH氧化酶Nox1和Duox2在小鼠肠炎中的表达和意义[J]. 世界华人消化杂志,2015,23(10): 1560-1567.
[17]Kato M, Marumo M, Nakayama J, et al. The ROS-generating oxidase Nox1 is required for epithelial restitution following colitis[J]. Exp Anim,2016,65(3): 197-205.
[18]Yokota H, Tsuzuki A, Shimada Y, et al. NOX1/NADPH oxidase expressed in colonic macrophages contributes to the pathogenesis of colonic inflammation in trinitrobenzene sulfonic acid-induced murine colitis[J]. J Pharmacol Exp Ther,2017,360(1): 192-200.
[19]Szanto I, Rubbia-Brandt L, Kiss P, et al. Expression of NOX1, a superoxide-generating NADPH oxidase, in colon cancer and inflammatory bowel disease[J]. J Pathol,2005, 207(2): 164-176.
[20]Chen L, Xu B, Liu L, et al. Cadmium induction of reactive oxygen species activates the mTOR pathway, leading to neuronal cell death[J]. Free Radic Biol Med, 2011, 50(5): 624-632.
[21]Singel K L, Segal B H. NOX2-dependent regulation of inflammation[J]. Clin Sci, 2016, 130(7): 479-490.
[22]Hayes P, Dhillon S, O’Neill K, et al. Defects in NADPH oxidase genes NOX1 and DUOX2 in very early onset inflammatory bowel disease[J]. Cell Mol Gastroenterol Hepatol,2015,1(5): 489-502.
[23]Dhillon S S, Fattouh R, Elkadri A, et al. Variants in nicotinamide adenine dinucleotide phosphate oxidase complex components determine susceptibility to very early onset inflammatory bowel disease[J]. Gastroenterol,2014, 147(3): 680-689. e2.
[24]Nauseef W M. Biological roles for the NOX family NADPH oxidases[J]. J Biol Chem, 2008, 283(25): 16961-16965.
[25]Brown D I, Griendling K K. Nox proteins in signal transduction[J]. Free Radic Biol Med, 2009, 47(9): 1239-1253.
[26]Lipinski S, Till A, Sina C, et al. DUOX2-derived reactive oxygen species are effectors of NOD2-mediated antibacterial responses[J]. J Cell Sci,2009, 122(Pt 19): 3522-3530.
[27]Achitei D, Ciobica A, Balan G, et al. Different profile of peripheral antioxidant enzymes and lipid peroxidation in active and non-active inflammatory bowel disease patients[J]. Dig Dis Sci, 2013, 58(5): 1244-1249.
[28]杜莉莉, 吕润潇,杨晓漪,等.胎盘间充质干细胞低氧培养液对肠黏膜上皮细胞氧化应激损伤的保护作用[J]. 中国医科大学学报,2016 ,45(2) :131-135,140.
[29]Chu F F, Esworthy R S, Doroshow J H, et al. Deficiency in Duox2 activity alleviates ileitis in GPx1-and GPx2-knockout mice without affecting apoptosis incidence in the crypt epithelium[J]. Redox Biol,2016(11): 144-156.
编辑 陈瑞芳
Significance of NADPH oxidases NOX1, NOX2 and DUOX2 expression in inflammatory bowel disease
Xuan Qingxia1, Dai Faliang1,Chen Pan1, Feng Dandan1, Jin Jianjun1, Gao Qiang1,2*
(1.DepartmentofGastroenterologyandHepatology,TheFirstAffiliatedHospital,andCollegeofClinicalMedicineofHenanUniversityofScienceandTechnology,Luoyang471003,HenanProvince,China; 2.DepartmentofGastroenterologyandHepatology,BeijingRehabilitationHospital,CapitalMedicalUniversity,Beijing100114,China)
Objective To investigate the expression of NOX1, NOX2 and DUOX2, the members of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase family, in large intestine of patients with inflammatory bowel disease (IBD) and mouse colitis and analyze the relationship between these enzymes and IBD. Methods The samples were collected from large intestine of patients with IBD. Mouse chronic colitis model was established by using eight to ten-week-old C57BL/6 mice treated with three cycles of drinking 1.0% dextran sulfate sodium (DSS) for 7 days plus drinking water for 14 days. Mouse body weight change and hematoxylin-eosin (HE) staining of colon sections were used to evaluate the severity of inflammatory lesions. The expression of NOX1, NOX2 and DUOX2 gene and protein were detected by real-time quantitative reverse transcription polymerase chain reaction (RT-PCR) and immunohistochemistry, respectively. Results The expression of NOX1, NOX2 and DUOX2 mRNA significantly increased in human inflamed tissues compared to paired normal tissues (allP<0.01). NOX2 mRNA higher in male patients than female. Unlike human, only Nox2 mRNA increased by 2.4 times in chronic colitis mice, Nox1 and Duox2 mRNA remained unchanged. These three proteins in IBD patients increased (allP<0.01); in chronic colitis mice Nox2 and Duox2 proteins increased (bothP<0.01), but Nox1 did not. Conclusion NOX1, NOX2 and DUOX2 not only play an important role in maintaining the normal physiological function, but also were associated with early onset IBD.
NADPH oxidases; NOX1; NOX2; DUOX2; inflammatory bowel disease (IBD)
国家自然科学基金(81370487)。This study was supported by National Natural Science Foundation of China (81370487).
时间:2017-04-13 19∶37
http://kns.cnki.net/kcms/detail/11.3662.R.20170413.1937.010.html
10.3969/j.issn.1006-7795.2017.02.014]
R574
2016-12-07)
*Corresponding author, E-mail:gaoq@ccmu.edu.cn
