前庭毛细胞的反相激活模式
2017-03-07丁大连亓卫东RichardSalvi
丁大连 亓卫东 Richard Salvi
1 Center for Hearing and Deafness,State University of New York at Buffalo 2复旦大学附属华山医院耳鼻咽喉头颈外科
1 前庭终器的功能与毛细胞分型
位于内耳腔内的前庭外周感受器具备感受重力和机体加速运动的功能,通过前庭神经纤维将前庭感觉上皮接收到的平衡改变信息传入到脑干的前庭神经核,前庭神经核再通过前庭眼束、前庭小脑束、前庭脊髓束、脑干网状结构、及大脑皮层的前庭中枢等投射神经网络把来自前庭周边感受器的平衡改变信息传递到机体其它参与平衡感知和平衡调控的器官或系统,从而使机体通过一系列的神经反射或主动调节来共同维持身体的平衡。前庭的感觉上皮分布在球囊斑(macula of saccule)、椭圆囊斑(macula of utricle)和三个半规管壶腹腔内的壶腹嵴(ampullary crista)(图1)。这些感觉毛细胞虽然在前庭池内分布在不同的区域,但其基本组织结构都是由传入和传出神经支配的前庭毛细胞及位于毛细胞下层的支持细胞所组成。前庭感觉毛细胞分为两种,它们分别是I型毛细胞和II型毛细胞,这两种毛细胞表面的纤毛束中都有一根排列在表皮板一侧的动纤毛(kinocilium)和沿着动纤毛排列的50-110根静纤毛(stereocilia),其中I型毛细胞的静纤毛束沿着动纤毛由长变短(图2A),而II型毛细胞的静纤毛束很短仅为一小撮(图2B)。I型毛细胞的细胞体呈烧瓶状被来自前庭大神经元的“古希腊葡萄酒瓶”(ancient Greek wine bottles)样或“酒杯型”(chalice-type)样的I型传入神经末梢所包饶,支配I型毛细胞的前庭传出神经纤维末梢则附着在前庭大神经元传入神经杯底部的传入神经纤维;II型毛细胞的胞体呈柱状,来自前庭小神经元的“郁金香”样的传入神经末梢和前庭传出神经末梢在II型毛细胞的底部与之建立突触联系。简单地说,两种不同类型毛细胞的不同之处表现在:1.静纤毛的长度不同。2.细胞的形状不同。3.神经末梢的连接方式不同。4.与之相连接的前庭神经元不同(图2C)。

图1 摘除镫骨并打开前庭池的外壁以充分暴露前庭池内的球囊、椭圆囊、三个半规管壶腹、外半规管非壶腹端单脚及总脚。Fig.1 The stapes and the lateral wall of the vestibular cavity were completely removed to fully expose the saccule,utricle,the ampulla of the three semicircular canals,non-ampulla ends of lateral semicircular canal and common crus in the vestibular cavity.
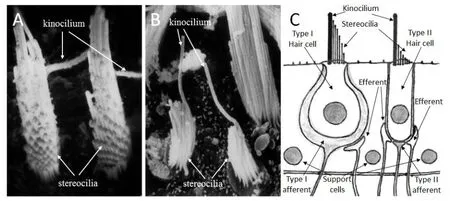
图2 I型毛细胞和二型毛细胞的形态学差别。A.应用扫描电镜显示I型毛细胞表面的长静纤毛特征。B.应用扫描电镜显示II型毛细胞表面的短静纤毛特征。C.I型毛细胞和II型毛细胞的模式图显示两种毛细胞从静纤毛长度到细胞形态以及传入神经末梢和传出神经的连接方式都完全不一样。Fig.2 Morphological differences between type I and type II hair cells.(A)Note the characteristic,long stereocilia on type I hair cells as shown by scanning electron microscopy.(B)Note characteristic short stereocilia on type II hair cells as shown by scanning electron microscopy.(C)Schematic illustrating major differences of type I and type II hair cells in terms of length of stereocilia,shape of the cell body and afferent and efferent synapses and nerve endings.
2 前庭毛细胞接受刺激的被动模式
球囊斑和椭圆囊斑毛细胞的动纤毛和静纤毛插入到其上方覆盖的辅助结构-凝胶状的耳石膜(gelatinous otolithic membrane),壶腹嵴毛细胞的纤毛插入到覆盖在壶腹嵴表面的终帽(gelatinous cu⁃pula)。球囊斑和椭圆囊斑毛细胞通过与其表面覆盖的耳石膜之间的相对位置移动感受因体位上下左右前后平移所引起的直线加速度(linear acceler⁃ation)刺激,而壶腹嵴上的毛细胞则通过与其表面覆盖的终帽之间的相对位置移动感受因体位旋转而引起的角加速度(angular acceleration)运动的刺激。在静止状态下,前庭各终器可以感知地球引力的作用并通过反射或调解机制与机体其它参与感知平衡和调控平衡的器官及系统共同使机体保持在静止状态下的平衡体位,但是当身体开始一个加速移动或者终止一个匀速移动时,由于前庭终器表面覆盖的辅助装置的惯性作用,使之与前庭毛细胞之间发生了相对位移,从而导致前庭毛细胞纤毛的被动弯曲而对感觉毛细胞构成实质性刺激。位于前庭池内的球囊斑和椭圆囊斑分别处于两个相互垂直的平面,因此它们可以分别感受体位朝着不同方向的上下或左右或前后的平面直线加速运动。连接到椭圆囊的三个半规管则分别位于三个不同的相互垂直的弧形平面,当体位沿着某一角度发生旋转而使与旋转平面相对应半规管内的内淋巴液发生向壶腹或逆壶腹流动时,该壶腹嵴上的毛细胞则感受到沿着该弧形半规管旋转所发生的最大角加速运动的刺激。
3 前庭各终器毛细胞纤毛的排列及其可变电阻式膜电位调节方式
无论是哪个前庭终器,前庭毛细胞表面纤毛的排列位置都遵循着一个统一的规则,就是那根决定毛细胞极性的动纤毛总是排列在毛细胞表皮板的一端(图3)。在球囊斑和椭圆囊斑的中心地带各有一个狭窄的毛细胞区域被称之为微纹区(striola zone),在球囊斑微纹区和椭圆囊斑微纹区的两侧分别是两个囊斑的周边区(marginal zone)(图3A,B)。我们发现南美栗鼠球囊斑和椭圆囊斑微纹区的I型毛细胞约占该区域毛细胞总数的90%,然而周边区的I型毛细胞所占比例大约在65%左右[1],这一观察结果与以往其它实验动物种类囊斑上两种毛细胞在不同区域所占比例的报道基本一致[2-5]。球囊斑周边区前庭毛细胞的动纤毛都是排列在背离微纹区的一侧(图3D),而位于椭圆囊斑周边区的毛细胞动纤毛都是排列在靠近微纹区的一侧(图3E)。外半规管壶腹嵴毛细胞的动纤毛统一排列在朝着椭圆囊的一侧,但上半规管和后半规管壶腹嵴毛细胞的动纤毛则都是排列在背离椭圆囊的一侧(图3F)。
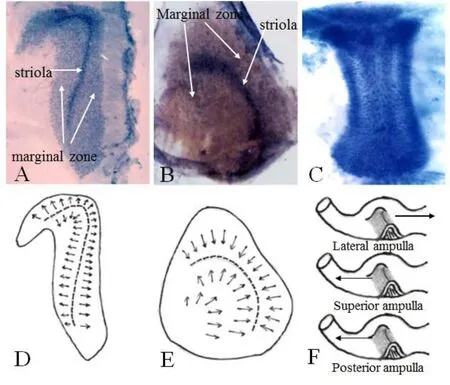
图3 球囊斑和椭圆囊斑的微纹区和周边区划分以及壶腹嵴毛细胞的极性。A.球囊斑铺片显示微纹区和周边区。B.椭圆囊斑铺片显示微纹区和周边区。C.壶腹嵴铺片。D.球囊斑周边区毛细胞的动纤毛都是排列在离开微纹区的一侧。E.椭圆囊斑周边区毛细胞的动纤毛都是排列在朝着微纹区的一侧。F.外半规管壶腹嵴毛细胞的动纤毛都是排列在朝着椭圆囊的一侧;上半规管和后半规管壶腹嵴毛细胞的动纤毛都是排列在朝着半规管的一侧。Fig.3 The striola zone and marginal region of the macula of the saccule and the utricle as well as the polarity of hair cells in cristae.(A)Striola and marginal regions as seen from surface preparation of macula of saccule.(B)Striola and marginal region as seen from surface preparation of macula of utricle.(C)Surface preparation of crista.(D)Kinocilium of hair cells in marginal regions of macula of saccule face away(arrow)from the striola.(E)Kinocilium of hair cells in marginal regions on macula of utricle face(arrow)the striola.(F)In the crista of lateral semicircular canal,the kinocilium of hair cells face(arrow)the utricle whereas the kinocilium of hair cells in the crista of the superior and posterior canals are face away(arrow)from the utricle.
前庭毛细胞的动纤毛位于表皮板的一侧,其位置决定了毛细胞的极性,前庭毛细胞静纤毛的弯曲方向则决定了毛细胞表面膜电阻的变化和神经冲动的发放。因此,前庭毛细胞顶部的静纤毛运动可以被理解为是一个可变电阻,这个可变电阻具有十分重要的调节毛细胞内电流的作用。当静纤毛朝向动纤毛弯曲时引起毛细胞膜电位的去极化,但是,当静纤毛朝着背离动纤毛的方向弯曲时则引起毛细胞膜电位的超极化。毛细胞的去极化使膜电位减弱而使传入神经冲动信号的发放加强,从而使毛细胞处于兴奋状态;毛细胞的超极化则使膜电位加强使传入神经冲动信号的发放减弱,从而使毛细胞处于抑制状态[6](图4)。因此,正是由于前庭毛细胞静纤毛与其上方覆盖的辅助结构(耳石膜或者终帽)的相对往复运动才使毛细胞发放出脉冲式的神经冲动信号[7-13]。
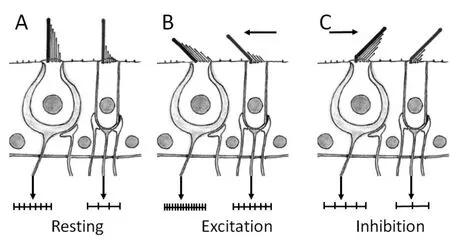
图4 毛细胞纤毛弯曲方向的不同导致了毛细胞的去极化或超极化。A.I型毛细胞和II型毛细胞在静止状态下的静息电位。B.当静纤毛朝着动纤毛方向弯曲时造成毛细胞膜的去极化改变从而使细胞兴奋。C.当静纤毛朝着背离动纤毛方向弯曲时造成毛细胞膜的超极化改变从而使细胞抑制。Fig.4 Schematic showing the direction of stereocilia bending that leads to depolarization or hyperpolarization of hair cells.(A)Spike discharges of type I and type II afferent fibers in the unstimulated,resting state.(B)Note increase in spike discharge rate(excitation)in type I and type II afferent fibers when stereocilia of hair cells bend toward the kinocilium causing hair cell depolarization.(C)Note decrease in spike discharge rate(inhibition)when the stereocilia of hair cells bend away from the kinocilium that causes the hair cell hyperpolarization.
4 发生在球囊斑和椭圆囊斑毛细胞的不平衡刺激模式
球囊斑和椭圆囊斑微纹区两侧周边区毛细胞的动纤毛分别排列在背离微纹区或朝着微纹区的相反位置,说明两个囊斑周边区毛细胞的极性完全相反。当机体沿着一个囊斑的平面从垂直于微纹区的方向产生一个加速运动时,该囊斑微纹区一侧周边区的毛细胞会因为静纤毛朝着动纤毛的方向弯曲使该侧毛细胞发生去极化而处于兴奋状态,而另一侧周边区的毛细胞却必定因静纤毛朝着离开动纤毛的方向弯曲使该侧毛细胞发生超极化而处于抑制状态,这意味着同样的直线加速运动方向对同一个囊斑微纹区两侧感觉上皮产生了截然相反的刺激,从而造成该囊斑微纹区两侧的输入信号不平衡(图5)。这种囊斑两侧不平衡的刺激信号传入到与之相联系的前庭周边神经元,必然会造成一部分前庭神经元兴奋而另一部分前庭神经元抑制,接受兴奋刺激或抑制刺激的两组不同反应的前庭周边神经元再分别把这两种同时发生的相反刺激信号继续上传到中枢的前庭神经核,则必然使前庭核神经元也同时呈现出部分兴奋和部分抑制的相同刺激模式。显然,正是由于前庭感受器的这种不平衡输入模式才使直线加速度刺激被前庭中枢以同样不平衡兴奋状态有效感知。
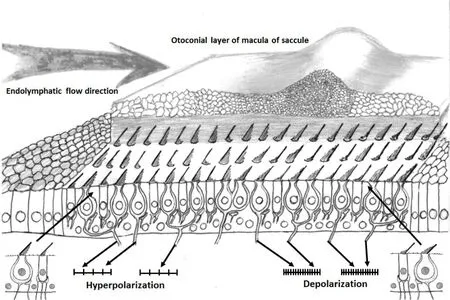
图5 垂直于微纹区的耳石膜位置移动造成球囊斑两侧周边区的毛细胞分别呈现去极化和超极化的相反刺激。Fig.5 Movement of the otolithic membrane perpendicular to the striola causes the hair cells in the periphery on opposite sides of the striola to respond in the opposite manner depolarization on the right side and hyperpolarization on the left side.
5 发生在壶腹嵴毛细胞的不平衡刺激模式
既然囊斑微纹区两侧毛细胞是接受完全相反的刺激模式,那么壶腹嵴毛细胞是不是也以这种相同的刺激模式使壶腹嵴一侧毛细胞兴奋而另一侧毛细胞抑制呢?可以确定的是,壶腹嵴毛细胞的动纤毛都是朝着一个统一的方向,即外半规管壶腹嵴上毛细胞的动纤毛都是朝着椭圆囊的方向,而上半规管和后半规管壶腹嵴上毛细胞的动纤毛都是朝着半规管的方向。如此看来,壶腹嵴毛细胞似乎需要的只是一个统一行动而并不需要像球囊斑和椭圆囊斑微纹区两侧的毛细胞那样对同一刺激表现出截然不同的反应。然而,壶腹嵴在壶腹腔内是一个横位的如同“水坝”样的马鞍形隆起,其上方覆盖的胶状质终帽可随内淋巴液的流动发生摇摆,从而使插入到终帽内的毛细胞纤毛发生弯曲以构成对毛细胞的刺激。我们不妨在此基础上再做一个更深入一步的假设性思考,这个思考的前提是根据壶腹嵴的不平坦堤坝状“地势”。以外半规管为例,当外半规管内的内淋巴液因角加速旋转或者温度刺激朝着壶腹嵴方向流动时,壶腹嵴面对外半规管侧的毛细胞纤毛显然会随着终帽朝着椭圆囊方向的偏移发生方向一致的弯曲,从而使外壶腹嵴半规管侧毛细胞的静纤毛朝着动纤毛的方向弯曲而产生出同步的去极化兴奋反应,然而外壶腹嵴椭圆囊侧毛细胞的纤毛却很可能因为外壶腹嵴“堤坝”上方终帽朝着椭圆囊侧的偏转使被“堤坝”遮挡的外壶腹嵴底部形成一个朝着壶腹嵴方向流动的漩涡式回流,这股从椭圆囊朝着外壶腹嵴底部流动的液体使壶腹嵴椭圆囊侧的毛细胞纤毛因终帽被回流推向壶腹嵴顶部方向而发生了朝着壶腹嵴顶部方向的弯曲,这就可能会使外壶腹嵴椭圆囊侧毛细胞的静纤毛朝着背离动纤毛方向发生偏移而产生同步的超极化抑制反应。这样的话,意味着壶腹嵴两侧毛细胞的极性虽然完全相同,但是由于壶腹嵴的隆起造成了壶腹嵴另一侧的液体回流,使壶腹嵴两侧的内淋巴液在同一时间都朝着壶腹嵴流动,从而使壶腹嵴两侧毛细胞分别接受到截然相反的兴奋或抑制刺激信号,由此便可能发生一侧壶腹嵴毛细胞呈现兴奋而另一侧壶腹嵴毛细胞反而呈现抑制的不平衡刺激模式(图6)。这个假设使我们不由得联想到沧州那对从下游掉进河水却在上游被找到的石狮子,虽然那个已被人们接受的标准解释是因为流水冲刷掉石狮子上游方向的泥沙从而促使石狮子在水中逆流朝着上游翻滚行进,但是那个所谓的标准解释显然忽略了另外一个更重要的事实,那就是在石狮子背后形成的漩涡式回流及其形成的泥沙堆积才是推动石狮子逆流行进的真正动力。我们提出的这个假设虽然可以根据暗礁背面的水流方向相反或者风在障碍物背面形成旋转回风使落叶聚集在障碍物背面等自然现象进行初步推测,但是还是有待于通过实际测试壶腹嵴两侧液体流变学的改变予以进一步证实。很早以前的实验研究就已经证实,从半规管注入的印度墨水无法通过终帽进入椭圆囊,由此得出终帽是从壶腹嵴的顶部延伸到对侧壶腹膜的顶部并将膜壶腹腔完全封闭的结论[8,14]。如果内淋巴液确实完全不能从终帽与膜壶腹之间的缝隙穿过,那么座落在壶腹嵴顶部的终帽就相当于一道屏障,而位于壶腹嵴两侧的毛细胞在内淋巴液流动时可能会因此而存在一个位相差,其作用方式也许就会像镫骨底板向前庭池内踏动时圆窗膜向外膨出那样形成外淋巴液在前庭阶是从蜗底朝着蜗尖流动而在鼓阶却是从蜗尖朝着蜗底流动。液体流动被终帽阻挡在壶腹嵴两侧形成的不同位相差同样可能造成壶腹嵴两侧毛细胞接受到不同的刺激。另外一种情况就是万一内淋巴液可以从终帽与壶腹嵴感觉上皮的细小间隙或终帽与膜壶腹之间的间隙流过,那么从壶腹嵴上方流过的液体也必然会在壶腹嵴的另一侧造成漩涡式的回流,同样会造成壶腹嵴两侧液体的流动方向不一致。为了观察当内淋巴液朝着壶腹嵴流动时壶腹嵴另一侧的液体流向,我们在解剖出来的南美栗鼠的椭圆囊内滴入1微升蓝色的ToPro-3染料,在没有液体流动的状态下,那滴染料会停留在椭圆囊缓慢地逐渐向四周扩散,但是当我们从外半规管用微量注射泵朝着壶腹嵴的方向以每分钟5微升的速度泵入透明液体的时候,我们发现随着半规管内透明液体朝着壶腹嵴的流动,椭圆囊内的染料在半规管灌流后三分钟左右并没有向椭圆囊内扩散而是流向椭圆囊侧的外半规管壶腹嵴,然而流向壶腹嵴的染料未能穿越终帽进入到半规管(图7)。这个现象说明,内淋巴液穿越终帽的可能性确实不大,但是当半规管内液体流向壶腹嵴的时候,壶腹嵴椭圆囊侧的液体也在朝着壶腹嵴的方向流动,因此壶腹嵴底部两侧的液体流动方向在同一时间可能呈现出相反方向的流动态势。根据这个简单的预初实验结果,我们初步推测壶腹嵴两侧的毛细胞在内淋巴液沿着半规管流动时可能会分别接受到截然相反的刺激模式。显然,这个推测还有待于更深入细致的半规管内液体流变学实验和前庭神经电生理学实验来予以进一步的证实。
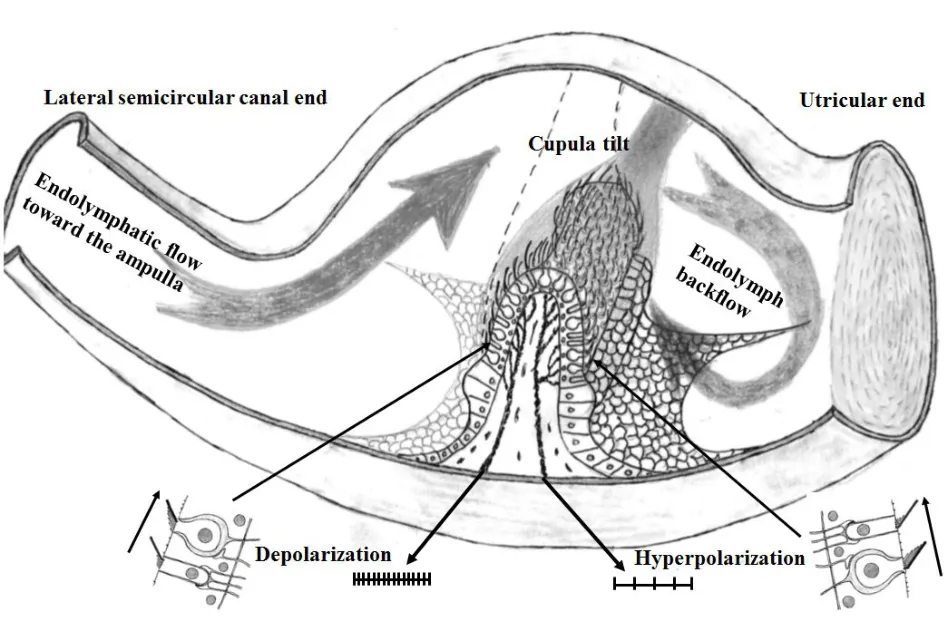
图6 朝着外半规管壶腹嵴的内淋巴流动造成半规管侧毛细胞去极化和椭圆囊侧毛细胞超极化的相反刺激。Fig.6 Schematic model of hypothesized flow of endolymphatic toward the crest of the ampulla of the lateral semicircular canal.Direct endolymphatic flow causes the depolarization on the left side and the proposed backflow or undercurrent(turbulence)on the right side causes hyperpolarization.

图7 从外半规管朝着壶腹嵴注入液体可见滴在椭圆囊的蓝色染料朝着壶腹嵴扩散。A.将蓝色染料滴在椭圆囊。B.向半规管内注入透明液体3分钟,可见蓝色染料朝着壶腹嵴流动。Fig.7 Experiment showing blue dye spread toward the ampulla when the liquid is perfused into the semicircular canal.(A)Blue dye was placed in the utricle before perfusion of the the semicircular canal.(B)Three minutes after perfusion toward the semicircular canal,the visible blue dye flows from the utricle to the crista of ampulla.
6 结论
综上所述,无论是在球囊斑或椭圆囊斑,还是在壶腹嵴,每个前庭感受区的毛细胞都对同一方向的加速运动刺激产生出一侧感受区前庭毛细胞兴奋而另一侧感受区前庭毛细胞抑制的不平衡刺激模式,由此必将造成与之相连的前庭周边神经元也同时呈现一部分神经元兴奋而另一部分神经元抑制的不平衡活动状态。这种发自前庭感受器并沿着前庭神经通路一路上传到中枢的不平衡刺激模式,将使我们对前庭各个终器在毛细胞水平上发生的同步但不平衡刺激方式和与之相联系的前庭神经元交替兴奋模式有一个全新的思考和认识。
1 丁大连,蒋海燕,王家东,等。卡铂损害灰鼠前庭神经元和I型毛细胞的定量观察 听力学与言语疾病杂志,2002.10(3):p.170-183.
2 Engstrom,H.,Wersall,J.Structure and innervation og the inner ear sensory epithelia.Int.Rev.Cytol.,1958.7:p.535.
3 Engstrom,H.,Wersall,J.The ultrastructural organization of the organ of Corti and of the vestibular sensory epithelia.Exp Cell Res,1958.14(Suppl 5):p.460-92.
4 Lindeman,H.H.,Studies on the morphology of the sensory regions of the vestibular apparatus with 45 figures.Ergeb Anat Entwick⁃lungsgesch,1969.42(1):p.1-113.
5 Lindeman,H.H.,Regional differences in structure of the vestibu⁃lar sensory regions.J Laryngol Otol,1969.83(1):p.1-17.
6 Hudspeth,A.J.,Corey,D.P.Sensitivity,polarity,and conductance change in the response of vertebrate hair cells to controlled me⁃chanical stimuli.Proc Natl Acad Sci U S A,1977.74(6):p.2407-11.
7 Engstrom,H.,Bergstrom,B.,Rosenhall,U.Vestibular sensory epi⁃thelia.Arch Otolaryngol,1974.100(6):p.411-418.
8 Schucnect,H.F.The vestibular system.Pathology of the ear.Har⁃vard University Press,Cambridge,Massachustts.,1974:p.129-131.
9 丁大连,亓卫东,曲雁,等。内耳解剖。丁大连主编内耳科学.中国科学技术出版社,2010:p.1-8.
10 丁大连,姜泗长。内耳的组织与解剖。丁大连姜泗长.豚鼠内耳解剖检验技术手册.学林出版社,1989.上海:p.24-35.
11 丁大连,李明,姜泗长,等。颞骨及内耳解剖。丁大连主编.内耳形态学.黑龙江科学技术出版社,2001.哈尔滨:p.10-17.
12 曲雁,丁大连,亓卫东,等。前庭生理。丁大连主编内耳科学.中国科学技术出版社,2010:p.26-33.
13 Schucnect,H.F.The vestibular sense organs.Pathology of the ear.Harvard University Press,Cambridge,Massachustts.1974:p.52-59.
14 Ramprashad,F.,Landolt,J.P.,Money,K.E.,et al.,Dimensional anal⁃ysis and dynamic response characterization of mammalian periph⁃eral vestibular structures.Am J Anat,1984.169(3):p.295-313.
