蝶形花植物猪屎豆自花传粉机制研究
2016-12-22郭艳峰蒋谦才孙红梅
郭艳峰, 刘 妍*, 蒋谦才, 孙红梅
( 1. 中山火炬职业技术学院 生物医药系, 广东 中山 528436; 2. 中山市国有森林资源保护中心, 广东 中山 528400 )
蝶形花植物猪屎豆自花传粉机制研究
郭艳峰1, 刘 妍1*, 蒋谦才2, 孙红梅2
( 1. 中山火炬职业技术学院 生物医药系, 广东 中山 528436; 2. 中山市国有森林资源保护中心, 广东 中山 528400 )
猪屎豆(Crotalariapallida)为典型的蝶形花植物,分布极广,是路边或遭破坏生境中最常见的先锋种之一,野外观察未发现其有营养生殖的现象,主要为种子繁殖。该研究通过对自然生境中猪屎豆开花物候、访花昆虫及繁育系统的研究,旨在从繁殖的角度阐述其快速扩张的能力。结果表明:尽管猪屎豆的主要访花昆虫是蜜蜂,但蜜蜂的访花频率极低 [(1.73±1.30)次/花序·h-1],且在整个花期内花药和柱头均被龙骨瓣包裹,蜜蜂访花时未成功接触柱头和花粉,不能实现传粉,因此蜜蜂不是猪屎豆有效的传粉昆虫,这与假说“蝶形花普遍是对膜翅目昆虫,尤其是对蜜蜂传粉的适应”不一致。人工授粉结果显示,猪屎豆为自交亲和种,不存在无融合生殖现象,其繁殖主要通过主动自交生产种子来实现,且在自交过程中长短花药都参与主动自交。这种自交方式不同于其他蝶形花植物的主动自交仅由短花药实现。对猪屎豆而言,长短花药均参与自交能够增加柱头的授粉几率,保证其在不利的生境中成功结籽,是其成功扩张的关键因素之一。
猪屎豆, 蝶形花, 主动自交, 先锋植物, 快速扩张
自交和异交是繁育系统研究的中心问题。自从Darwin(1876)起,人们认为异交是一种进化的传粉机制,是形成繁育系统中传粉多样性的主要动力。自花授粉因会引起自交衰退而被认为是被子植物繁育系统进化的绊脚石(Darwin,1876;Kalisz et al,2004)。然而,自然界的开花植物中有20%是高度自交,33%介于自交和异交之间(Kalisz et al,2004),故自交在交配系统中占很大比重。但随着对自然界多种传粉机制及其生态意义的不断揭示,人们对自交与异交的利弊有了新的认识。在缺乏传粉昆虫或花粉受限制时,异交种的结籽会受到限制(Burd,1994;Ashman et al,2004),而自交种能通过自交保障结籽(Larson & Barrett,2000;Kalisz et al,2004)。对处于开拓新生境或传粉条件不利的植物而言,自交能保证植物繁殖而使其避免灭绝消失(Stebbins,1957),即具主动自交的种在建立新居群或快速扩张方面比异交种更具优势(Baker,1974;Cruden & Lyon,1989;Kalisz et al,2004;Randle et al,2009;Shivanna,2014)。在扩张迅速及广布种中,自交比异交更普遍(Baker,1974;Shivanna,2014)。但目前大多关于植物快速扩张的研究主要在植物的种子产量、种子萌发优势及无性繁殖能力方面(Kraichak,2012;Berkovic et al,2014)。尽管开花物候、传粉机制和繁育系统是种子生产的基础,是植物居群结构及对变化环境适应能力的决定因素,但相关的详细研究却很不足(Shivanna,2014)。
猪屎豆属(Crotalaria)植物隶属于豆科(Leguminosae)蝶形花亚科(Papilionoideae),属内种普遍为热带杂草,极好地生长于路边、沙地及遭人类改变的地方(Moore,1978;Lewis,1987;Jacobi et al,2005)。我国约有40种,其中6种从印度、非洲、南美、美洲逃逸而来。不少种类可供药用,是药用植物开发的重点类群。猪屎豆(Crotalariapallida)植物全草有散结、清湿热等作用,近年来试用于抗肿瘤效果较好,主要对鳞状上皮癌,基底细胞癌有疗效(崔鸿宾等,1998);在药用、防沙固土、园林绿化等方面也应用较广(赵琼玲等,2008)。课题组近年来在对华南地区猪屎豆属药用植物进行野外调查过程中发现,猪屎豆是该属中分布最广,且在路边或遭破坏生境中最常见的先锋种之一,即使在烈日当空、多砂多砾的环境中仍然能生长。在野外未发现营养生殖现象,主要通过种子繁殖,其种子耐盐能力强、萌发率高,对环境适应力强(廖富林等,2011;严琳玲等,2012;罗天琼等,2013),是其快速建立新居群的关键因素。然而,有关该种是如何传粉来生产种子的报道却极为匮乏。本研究通过对猪屎豆进行传粉生物学研究,以明确以下研究目的:(1)开花物候及开花生物学特性;(2)访花昆虫的行为及传粉机制;(3)繁育系统。进而明确猪屎豆在新生境中成功生存的原因,同时为探讨蝶形花亚科的花形态与传粉关系的协同提供重要依据。
1 材料与方法
1.1 研究植物与观察地点
猪屎豆为猪屎豆属多年生草本植物(图版Ⅰ:A)。本种是世界广布种,原产于热带美洲,分布于大西洋海岸线、巴西、美洲、非洲及中国大部分地区,常见于路边、河边、沙地还有贫瘠的山坡。本种为典型的蝶形花植物,果实为豆荚,一般长为3.4 cm,成熟时褐色(图版Ⅰ: C),外界压力易导致开裂。花果期为9-12月。
本研究的野外观察和实验工作分别于2014年9月至2015年5月在广东省中山市陵岗村 (22°53′ N,113°45′ E,海拔715 m)完成。
1.2 开花生物学特性观察
在所研究植物的自然居群中,对所有植株在2014-2015年间不同月份的开花物候情况进行观察和记录,明确居群的初花期、盛花期、末花期。随机选取20朵花(2 朵/株)进行标记,从6:00-18:00每隔3 h观察记录每一朵花的开放时间、花药开裂时间、花谢时间,花冠脱落标志着单花花期结束。同时测量雌蕊、长雄蕊、短雄蕊的长度。

图版 Ⅰ 猪屎豆的植株、花、访花者及不同发育阶段花的雌雄蕊位置关系 A. 植株; B. 花序; C. 果实; D. 不同角度的花、雄蕊及蜜腺 (a. 花侧面, b.花正面, c. 10个雄蕊基部融合成鞘, d. 蜜腺); E. 访花者; F. 不同发育阶段花的雌雄蕊位置关系 (a. 刚刚开放的花, LA. 长雄蕊, SA. 短雄蕊, b. 长雄蕊与柱头接触, c. 柱头高于雄蕊,长花药开裂,短花药未开裂, d. 短花药开裂且与柱头接触, e-f. 柱头高于雄蕊,花药变干, g. 花冠脱落后的雌蕊)。 Plate Ⅰ Plant, flowers, visitor and the distance between stigma and stamens in different development stages’ flowers of Crotalaria pallida A. Plant growing in the roadsides; B. Flowers growning in the inflorescence; C. Fruits on the plants; D. Flowers (a. Flower side elevation, b. Flower vertical, c. Ten stamens’ fusing by the basal part into a sheath, d. A nectary); E. Visitors visiting flower; F. Flowers in different development stages (a. Flower just opening, LA. Long-anthers, SA. Short-anthers, b. Long-anthers contacting with the stigma, c. Stigma protrusion from ten stamens, d. Short-anthers contacting with the stigma, e-f. Stigma protrusion from ten stamens and ten anthers dried-up, g. Corolla and ten anthers dropped off).
1.3 柱头可受性与花粉活性的测定
在单花花期内每天于10:00-12:00之间,随机采取不同发育时期的新鲜花10 朵,用MTT法测定花粉活性(Dafni,1992)和柱头可受性(Rodriguez-Riao & Dafni,2000),并在显微镜(40×)下观察并统计着色和未着色花粉粒数目。
1.4 花粉组织化学的检测
随机采集所研究植物花药已经开裂的花,将新鲜的花粉刮到干净的载玻片上,用I2-KI或苏丹Ⅲ-Ⅳ染液进行染色,在显微镜下进行观察,每个样品重复3次并记录反应的颜色。若I2-KI显示蓝色或黑色则表明花粉有淀粉存在,若苏丹Ⅲ-Ⅳ显示红色则说明花粉有脂质存在(Dafni,1992)。
1.5 花蜜分泌量、含糖量的测定
在单花花期内每天于10:00-12:00,用1 μL的毛细管(RS-Sigma,USA)吸取不同发育时期的新鲜花10朵进行吸蜜,检测其花蜜分泌量,并用手持折光仪(ATAGO,Brix 0%~32%,Japan)直接测定花蜜含糖量(%)。处理中的花均在花开放前进行套袋。
1.6 访花昆虫种类及访花行为的观察
于2014年11月22-24日和2015年1月13-15日内,在居群中随机标记 5~10 株已开放的花序进行访花昆虫的观察,具体是从7:00-18:00每隔2 h观察、统计0.5 h内访花昆虫的种类、数量,对其进行拍照、描述,并捕捉昆虫作凭证标本。
1.7 繁育系统检测
(1)自然对照:不做任何处理,挂牌直至结实。(2)人工自交:随机选择即将开放的花,套袋,花开后授同株花药花粉。(3)人工异交:随机选择即将开放的花,去掉花药,套袋,花开后授异株花药花粉。(4)主动自交: ①完全套袋,随机选择即将要开放的花,套袋,不做其他处理,挂牌直至结实; ②去长花药套袋,随机选择即将开放的花,去掉长花药,套袋,挂牌直至结实; ③去短花药套袋,随机选择即将要开放的花,去掉短花药,套袋,挂牌直至结实。(5)去全部花药套袋: 随机选择即将要开放的花,去掉花药,套袋。(6)去全部花药不套袋: 随机选择即将要开放的花,去掉花药,不套袋。1个月后收取所有处理的果实,统计各个处理的结实率。
所有数据统计分析均采用SPSS 15.0软件,统计数据用平均值 ± 标准差,n表示样本量。
2 结果与分析
2.1 开花物候及花的形态特征
在所观察的居群中,猪屎豆的初花期为9月上旬,盛花期从10月中下旬开始,持续到第2年的5月,末花期为5月中下旬。盛花期期间,平均每个植株有(10.27 ± 4.34)个花序(n= 30),每个花序开花(15.63 ± 5.86)朵(n=30)。猪屎豆的开花时间不一致,一天当中都有花开,大多集中在下午4:00-6:00。单花序花期约40 d(n= 20),单花花期10 d(n= 20)。
猪屎豆为蝶形花,总状花序顶生,花由下向上开(图版Ⅰ:B)。花冠由旗瓣、翼瓣和龙骨瓣组成,旗瓣狭细,翼瓣着生在旗瓣的内侧并紧贴龙骨瓣,下部包裹龙骨瓣,两片龙骨瓣合生(图版Ⅰ:D-a, b),10枚雄蕊呈两轮,5枚长雄蕊,5枚短雄蕊,长短雄蕊交替排列,10枚雄蕊基部融合成鞘(图版Ⅰ:D-c),子房成线状。龙骨瓣包裹10枚雄蕊和雌蕊(图版Ⅰ:D-a),雌蕊位于正中央,蜜腺位于融合花丝的基部(图版Ⅰ:D-d)。即将开放的花,旗瓣紧抱着翼瓣,二者与龙骨瓣紧靠。第1天时,旗瓣翼瓣与龙骨瓣开始分离,第2天龙骨瓣与旗瓣翼瓣约成90°角,旗瓣翼瓣依然未分离。第3天开始,旗瓣翼瓣分离,旗瓣与翼瓣约成60°夹角,但未完全展开。第5天旗瓣翼瓣完全展开。长花药在花开的第3天开裂,短花药在第4天开裂,第7天长短花药均干裂。
2.2 雌雄蕊位置关系
在整个开花进程中,雌雄蕊的位置关系如图版Ⅰ:F和图1所示,花刚开时,雌蕊最长,长短雄蕊均低于柱头,短雄蕊低于长雄蕊(图版Ⅰ:F-a),随着开花进程,长短雄蕊都逐渐升高,到第3天,长花药与柱头接触,短花药仍低于柱头(图版Ⅰ:F-b)。到第5天,短花药长至柱头高度,与柱头接触,长雄蕊低于短雄蕊与柱头(图版Ⅰ:F-d)。从第6天开始,柱头超过长短花药(图版Ⅰ:F-e,f)。
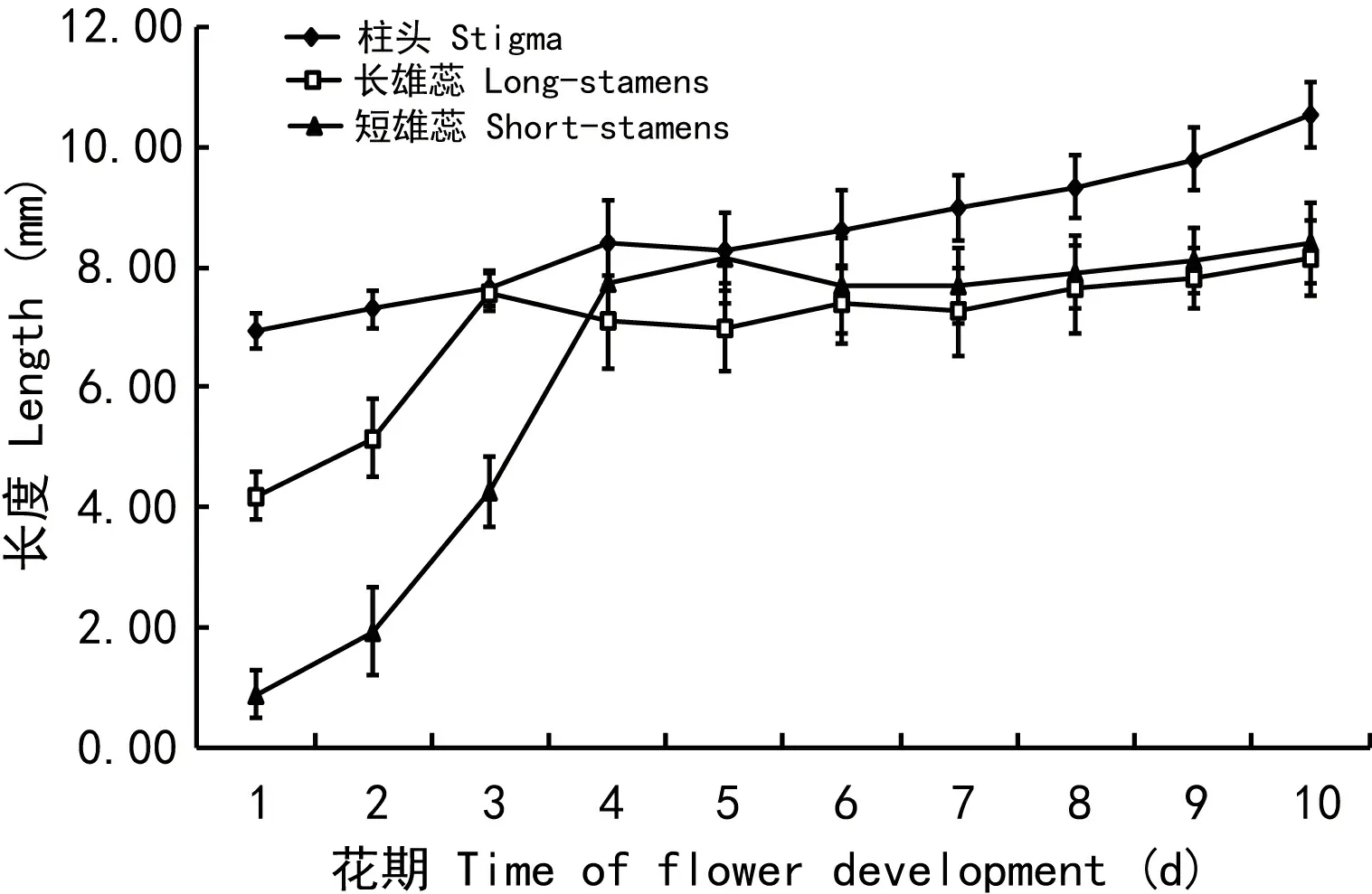
图 1 花期内猪屎豆雌雄蕊长度变化Fig. 1 Distance between stigma and stamens during flowering

图 2 猪屎豆花粉、柱头活性变化Fig. 2 Pollen viability and stigma receptivity of flowers
2.3 花粉活性和柱头可受性、花蜜量和含糖量、花粉组织化学特性
如图2所示,在整个花期内,柱头都具有活性。长短花药花粉活性在花药刚开裂是较高,均在90%以上,随着开花进程,花粉活性略微降低,但在花谢时,长短花药的花粉活性仍高于75%。花蜜检测结果显示(图3),刚开的花,花蜜量仅(0.09 ± 0.05) μL,随着开花时间的延长,从第3天开始,花蜜量显著升高(t= 9.732,df= 18,P= 0.001),花谢时花蜜量为(2.55 ± 0.72)μL。花刚开时,含糖量为(3.92 ± 2.07)%,从第3天开始,含糖量显著升高(t= 14.805,df= 18,P= 0.000),花谢时含糖量为(16.75 ± 2.14)%。猪屎豆的花粉经I2-KI染色后显示蓝色,表明花粉有淀粉存在,苏丹Ⅲ-Ⅳ染色后显示红色,说明花粉有脂质存在。
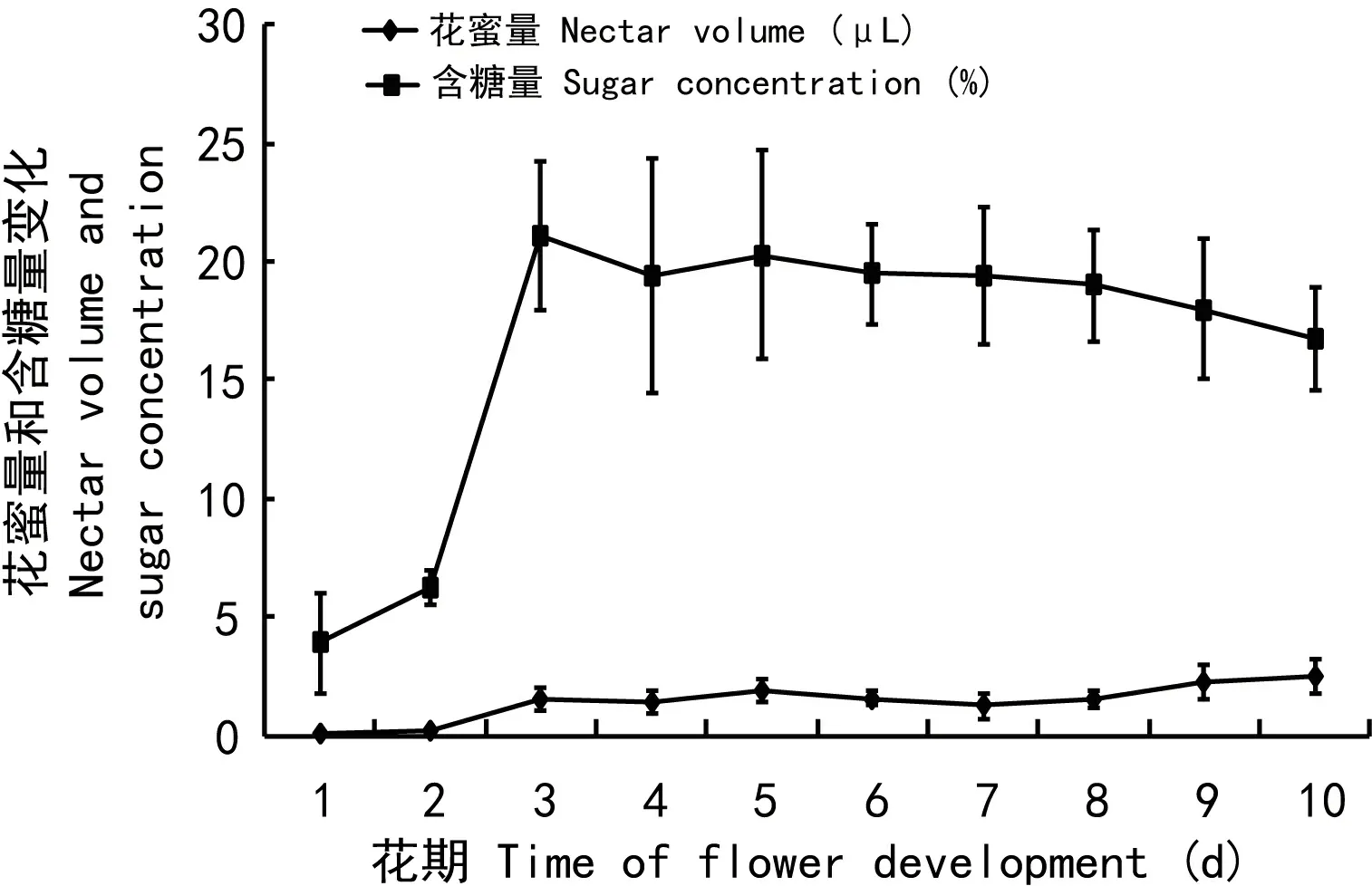
图 3 猪屎豆花期内花蜜量、含糖量变化Fig. 3 Nectar production and sugar concentration of
2.4 访花昆虫种类及访花行为
访花昆虫主要是蜜蜂(Apidaesp.),偶尔见到蝴蝶。蜜蜂平均访花频率为4.3(0.06~1.40)次/花序·30 min-1,访花高峰期为9:00-9:30、14:30-15:00。蜜蜂沿着花序从下往上或从上往下访花,访花时蜜蜂先在花附近盘旋,然后降落在花上,用足抱住翼瓣,头探入花冠基部的蜜腺吸蜜。吸蜜过程中,蜜蜂不是正面访花,其腹部未压在龙骨瓣上,而是避开龙骨瓣,腹部位于花梗的上方(图版Ⅰ:E)。由于蜜蜂未压在龙骨瓣上,被龙骨瓣包裹的柱头不能探出,花粉也不能通过挤压龙骨瓣释放,因此蜜蜂不能实现传粉,不是有效的传粉昆虫。
2.5 繁育系统
繁育系统检测结果显示(表1),人工自花授粉和人工异花授粉的结实率分别为12.38%、13.68%),差异不显著(t= 0.078,P= 0.802),说明猪屎豆不存在自交不亲和。去全部花药套袋结实率为0,表明猪屎豆不存在无融合生殖。完全套袋结实率较高(49.26%),说明猪屎豆能够发生主动自交。去长花药套袋和去短花药套袋有结实率,表明长花药和短花药都能实现主动自交。
3 讨论
豆科植物中, 蝶形花亚科的大多植物具两侧对称的蝶形花(Tucker,2003),花由旗瓣、翼瓣和龙骨瓣组成,雄蕊和雌蕊被龙骨瓣紧包,这种花型被认为是对膜翅目昆虫,尤其是蜜蜂传粉的适应 (Faegri &Van der Pijl,1979)。旗瓣一般较大,色彩艳丽,起吸引传粉昆虫的作用;翼瓣为访花者提供降落平台,传粉昆虫停在翼瓣的边缘,将吻通过缺口伸到小花的基部取食花蜜;访花者吸取花蜜时挤压龙骨瓣,龙骨瓣受外力后使包裹的成熟花粉借助张力释放(黄利春等,2014)。蝶形花植物花粉的释放必须在昆虫的参与下才能完成,且这种花结构与其传粉功能群协同进化形成的花粉呈现机制有以下4种:弹花机制、活塞机制、瓣膜机制和毛刷机制(Westerkamp,1997;Westerkamp & Weber,1999;Etcheverry et al,2003;Galloni et al,2007;黄利春等,2014)。在4种机制中,访花者挤压旗瓣和翼瓣进行吸蜜时,柱头会伸出龙骨瓣,花粉会通过龙骨瓣尖端的小孔释放在访花者的腹部,这样反复访花过程完成柱头的授粉。
表 1 不同授粉处理的结实率 (%)
Table 1 Fruit set in the pollination experiments

处理Treatment花数(株)Numberofflowers(Stems)结实率Fruitset(%)标准差s变异系数CV自然对照Openpollination220(10)34.0921.740.64完全套袋Autonomousautogamy136(10)49.263.980.08人工自交Experimentaltestforautogamy105(10)12.3810.120.82人工异交Experimentaltestforexnogamy117(10)13.689.870.72去全部花药套袋Testforagamospermy148(10)0.000.000.00长花药自交(花药开裂前移除短花药),套袋Testforlong⁃anthersselfing(short⁃anthersremovedbeforeantherdehiscence),bagged51(10)17.656.060.34短花药自交(花药开裂前移除长花药),套袋Testforshort⁃anthersselfing(long⁃anthersremovedbeforeantherdehiscence),bagged67(10)34.3321.330.62去雄,不套袋Allstamensremovedbeforean⁃therdehiscence,unbagged118(10)1.693.311.96
猪屎豆也具有蝶形花的特征,是蝶形花植物中为数不多的二体雄蕊。猪屎豆的花具有与其他蝶形花类似的吸引传粉者的特征:鲜艳的花冠、花序、分泌花蜜,且花蜜在花药开裂后显著增加;野外观察显示其访花者也是蜜蜂为主。然而猪屎豆的花部特征及访花者的访花行为与其他蝶形花植物有很大区别:大多数蝶形花植物如小花冠属(Coronillaemerus)、紫穗槐属(Amorphacanescens)的旗瓣成心形或卵形,旗瓣大,颜色鲜艳,部分还有红色的纹理起蜜导作用;猪屎豆花的旗瓣狭细,缺乏蜜导。同时,猪屎豆的访花昆虫蜜蜂的访花行为与当前蝶形花亚科中常见的4种机制不一样。蜜蜂降落时,用足抱住翼瓣,头伸进花冠底部吸花蜜,然而蜜蜂并没有正面降落着陆在翼瓣上,未对龙骨瓣产生压力。因此当蜜蜂吸蜜时,柱头未伸出龙骨瓣,花粉也未得到释放。故猪屎豆虽具蝶形花特征且有报酬物花蜜,访花者也是膜翅目昆虫,然而访花者数量少且访花行为的不同,导致猪屎豆不能像其他蝶形花科植物一样完成传粉过程。繁育系统处理中,去全部雄蕊不套袋处理的结实率低(1.69%)也表明猪屎豆在自然状态下缺乏有效传粉者。
自交能保障植物在不良环境中成功繁殖。猪屎豆属中已有主动自交现象报道,且被认为是该属植物在不良生境中生存及快速扩张的主要决定因素(Etcheverry,2001;Etcheverry et al,2003;Jacobi et al,2005;Shivanna,2014)。猪屎豆属植物的主动自交主要通过短花药与柱头接触实现(Etcheverry et al,2003;Jacobi,2005;Shivanna,2014)。本研究对猪屎豆整个开花过程花部特征、雌雄蕊位置关系的观察及繁育系统研究结果显示,猪屎豆主要通过自交实现结实,但猪屎豆的长短花药开裂时间和功能与同属报道的其他种不同。其他种长花药通常在花开前24 h开裂,花开后第2天长花药和花丝干枯,长花药未与柱头接触;短花药在开花后24 h开裂,此后花丝不断伸长将花粉送至柱头上实现自交(Etcheverry et al,2003;Shivanna,2014)。本研究中,猪屎豆的长花药在花开后第3天开裂,且开裂时花药即接触到柱头实现自交;短花药在花开后第4天开裂,并在第5天与柱头接触实现自交。与其他种相比,长短花药共同参与自交能提高授粉率,且在整个花期中,长短花药的花粉活性及柱头可受性均保持在较高水平,也是提高成功自交的保障之一。
对于传粉者不稳定或效率不高的植物而言,通过主动自交实现生殖保障能避免植物种的消失(Stebbins,1957;Rodriguez-Riano et al,2007;Freitas & Sazima,2009),同时能促进植物扩张及建立新的居群(Randle et al,2009;Shivanna,2014)。在快速扩张的类群中,繁育系统由异交向自交进化已经出现了很多次(Shivanna,2014)。猪屎豆虽具有蝶形花的特征、有花蜜,但生长环境缺乏传粉者或传粉者不稳定或者效率不高,使其逐渐向自交进化。主动自交能够保证其成功授粉结籽,是一种进化的行为,同时旗瓣变小,花蜜分泌量减少等,可认为是异交向自交进化过程中节约资源的表现。
ASHMAN TL, KNIGHT TM, STEETS JA, 2004. Pollen limitation of plant reproduction: ecological and evolutionary causes and consequences [J]. Ecology,85(9):2408-2421.
BAKER HG, 1974. The evolution of weeds [J]. Ann Rev Ecol Syst,5(1):1-24. BERKOVIC B, SUSANA C, BARRIO JM, 2014. Extending the life history of a clonal aquatic plant: dispersal potential of sexual and asexual propagules ofZosteranoltii[J]. Aquat Bot,113(2):123-129.
BURD M, 1994. Bateman’s principle and plant reproduction: the role of pollen limitation in fruit and seed set [J]. Bot Rev,60(1):83-139. CRUDEN RW, LYON DL, 1989. Facultative xenogamy: examination of a mixed-mating system [M]//BOCK J,LINHART YB. The evo-lutionary ecology of plants. Colorado: Westview Press:171-208.
CUI HB, ZHANG ZW, ZU LR, et al, 1998. Flora Republicae Popularis Sinicae [M]. Beijing:Science Press:344-379. [崔鸿宾,张振万,徐郎然,等,1998. 中国植物志 [M]. 北京:科学出版社:344-379.]
DAFNI A, 1992. Pollination ecology: a practical approach [M]. Oxford:Oxford Univ Press:1-57.
DARWIN C, 1876. The effects of cross and self fertilization in the vegetable kingdom [M]. London:Murray:1-50.
ETCHEVERRY AV, 2001. Role of staminal growth in delayed self-pollination ofCrotalariastipularia(Fabaceae: Papilionoideae) [J]. Beitr Biol Pflanzen,72(2):215-228.
ETCHEVERRY AV,PROTOMASTRO JJ, WESTERKAMP C, 2003. Delayed autonomous self-pollination in the colonizerCrotalariamicans(Fabaceae: Papilionoideae): structural and functional aspects [J]. Plant Syst Evol,239(1):15-28.
FAEGRI K, VAN DER PIJL L, 1979. The principles of pollination ecology [M]. Oxford: Pergamon Press:1-292.
FREITAS L, SAZIMA M, 2009. Floral biology and mechanisms of spontaneous self-pollination in five neotropical species of Gentianaceae [J]. Bot J Linn Soc,160(4):357-368.
GALLONI M, PODDA L, VIVARELLI D, et al, 2007. Pollen presentation, pollen-ovule ratios, and other reproductive traits in Mediterranean legumes (Fam. Fabaceae-Subfam. Faboideae) [J]. Plant Syst Evol,266(3):147-164.
JACOBI CM, 2005. Pollination biology of the exotic rattleweedCrotalariaretusaL. (Fabaceae) in NE Brazil [J]. Biotropica,37(3):357-363. KALISZ S, VOGLER DW, HANLEY KM, 2004. Context-dependent autonomous self-fertilization yields reproductive assurance and mixed mating [J]. Nature,430(7002):884-887.
KRAICHAK E, 2012. Asexual propagules as an adaptive trait for epiphylly in tropical leafy liverworts (Lejeuneaceae) [J]. Am J Bot,99(9):1436-1444.
LARSON BMH, BARRETT SCH, 2000. A comparative analysis of pollen limitation in flowering plants [J]. Biol J Linn Soc,69(4):503-520.
LEWIS GP, 1987. Legumes of Bahia [M]. Kew:Roy Bot Gard:369. LIAO FL, ZHANG J, LIU HN, et al, 2011. Study on hard seed percentage and seed germination characteristics ofCrotalariapallida-a plant for revegetation of deserted mines [J]. Seed, 30(8):34-38. [廖富林,张镜,刘惠娜,等,2011. 矿区复绿植物猪屎豆种子硬实率与萌发特性的研究 [J]. 种子,30(8):34-38.]LLOYD DG, SCHOEN DJ, 1992. Self-and cross-fertilization in plants. I. Functional dimensions [J]. Int J Pl Sci,153(3):358-369. LUO TQ, TAN JY, MO BT, et al, 2013. Study on the seed germination characteristics ofCrotalariapallida[J]. Hubei Agric Sci,52(16):3878-3883. [罗天琼,谭金玉,莫本田,等,2013. 猪屎豆种子萌发特性研究 [J]. 湖北农业科学,52(16):3878-3883.]
MOORE LR, 1978. Seed predation in the legumeCrotalaria[J]. Oecologia,34(2):185-202.
RANDLE AM, SLYDER JB, KALISZ S, 2009. Can differences in autonomous selfing ability explain differences in range size among sister taxa pairs ofCollinsia(Plantaginaceae)? An extension of Baker’s law [J]. New Phytol,183(3):618-629.
SHIVANNA KR, 2014. Reproductive assurance through autogamy in some annual weed species [J]. Proc Natl Acad Sci Ind Sect B Biol Sci,84(3):681-687.
STEBBINS GL, 1957. Self-fertilization and population variability in the higher plants [J]. Am Nat,91(861):337-354.
TUCKER SC, 2003. Floral development in legumes [J]. Plant Physiol,131(3):911-926.
WANG LC, JIN L, LI J, et al, 2014. Floral morphology and its relationship with pollination systems in Papilionoideae [J]. Acta Ecol Sin, 34(19):5360-5368. [黄利春,金樑,李晶,等, 2014. 蝶形花亚科植物花部适应机制与传粉系统 [J]. 生态学报,34(19):5360-5368.]
WESTERKAMP C, 1997. Keel blossoms: bee flowers with adaptations against bees [J]. Flora,192:125-132.
WESTERKAMP C,WEBER A, 1999. Keel flowers of the Polygalaceae and Fabaceae: a functional comparison [J]. Bot J Linn Soc,129(3):207-221.
YAN LL, ZHANG L, BAI CJ, et al, 2012. Evaluation of salt tolerance of differentCrotalariacultivars in germination stage [J]. Chin J Trop Agric, 32(11):13-17. [严琳玲,张龙,白昌军,等,2012. 不同猪屎豆品系种子萌发耐盐能力评价 [J]. 热带农业科学,32(11):13-17.]
ZHAO QL, BAI CJ, YU DG, 2008. Prospects for development and utilization ofCrotalaria[J]. Chin J Trop Agric, 28(4):71-74. [赵琼玲,白昌军,虞道耿, 2008. 猪屎豆属植物的利用价值及开发前景 [J]. 热带农业科学, 28(4):71-74.]
Autogamy mechanism ofCrotalariapallida(Fabaceae: Papilionoideae)
GUO Yan-Feng1, LIU Yan1*, JIANG Qian-Cai2, SUN Hong-Mei2
( 1.DepartmentofBiomedicine,ZhongshanTorchPolytechnic, Zhongshan 528436, Guangdong, China;2.ForestryScienceInstituteofZhongshanCity, Zhongshan 528400, Guangdong, China )
Crotalariapallidais a typical plant of Papilionoideae. Althought it has evolved various adaptations to establish and spread rapidly through prolific seed production and/or asexual propagules, self-fertilization is always an important aspect to decide its colonization ability. It is widely distributed and is the most common pioneer plant on the road sides or destroyed envioronment. In our observation, it lacks pollinators in nature and also lacks vegetative reproduction. How does this species survive in such unpredictable habitats? We focus on its floral morphology, reproductive biology and pollinator behavior to explain its reproductive success. We observed flower development, detected pollen viability and stigma receptivity, performed manual pollination and monitored the behavior of main pollinators inC.pallida. Papilionaceous flowers are reported to show adaptation for bees which are their pollinators. Few insects visited the flowers. Several bees were observed visiting flowers but visiting frequency was low (1.73 ± 1.30 visits·inflorescence-1·h-1). Because the stigma and stamens were enclosed in the petal of the keel in the flower development, the visitors were not contacting any reproductive organs, and they could be considered as visitors not effecting pollination. The results of pollination experiments showed thatC.pallidawas self-compatible and was capable of autonomous selfing. In our study, both long-and short-anthers’ descended time and function in fruit set were different from those autogamous species in the genus. ForC.pallida, both long-anthers and short-anthers participated into selfing and there was no difference in fruit sets between long-and short-anther selfing (t=1.786,P=0.060). While for other species in the genus, autogamy was realized by short anthers. ForC.pallida, therefore, automatic self-pollination increases the chances of seed production and protect it against extinction. Compared with other species, only short-anthers participate into autogamous pollination, both long- and short-anthers providing pollen grains to the stigma increases the success of pollination and fruit set, which is an evolutionary mechanism for the spread ofC.pallida. This results are consistent with the hypothesis that species proficient at autogamy are better adapted to establish new populations and thus can more readily expand their range.
Crotalariapallida, Papilionoideae, autogamouy, pioneer plant, rapidly spread
10.11931/guihaia.gxzw201508006
2015-11-21
2016-01-09
中山市科技计划项目(2013A3FC0352) [Supported by the Scientific and Technological Plan of Zhongshan City (2013A3FC0352)]。
郭艳峰(1984-),女,山西临汾人,博士,从事植物功能食品开发研究,(E-mail)guoyanfeng1984@sina.com。
*通讯作者: 刘妍,硕士,讲师,从事植物化学成分研究,(E-mail)583429336@qq.com。
Q948.1
A
1000-3142(2016)11-1318-07
郭艳峰,刘妍,蒋谦才,等. 蝶形花植物猪屎豆自花传粉机制研究 [J]. 广西植物, 2016, 36(11):1318-1324
GUO YF,LIU Y,JIANG QC,et al. Autogamy mechanism ofCrotalariapallida(Fabaceae: Papilionoideae) [J]. Guihaia, 2016, 36(11):1318-1324
