锁阳和肉苁蓉寄生方式的区别
2016-12-22陈金元陈学林郭楠楠马文兵达清璟张亥贤
陈金元, 陈学林, 郭楠楠, 马文兵, 达清璟, 张亥贤
( 西北师范大学 生命科学学院, 兰州 730070 )
锁阳和肉苁蓉寄生方式的区别
陈金元, 陈学林*, 郭楠楠, 马文兵, 达清璟, 张亥贤
( 西北师范大学 生命科学学院, 兰州 730070 )
锁阳和肉苁蓉都是中医药里重要的补益类药材,但由于过度采挖和采挖方式不当,目前它们的野生资源已濒临枯竭。肉苁蓉和锁阳分别是我国濒危和易危珍稀植物,研究二者寄生方式的特点与区别不仅可以促进锁阳和肉苁蓉的人工栽培,从而使野生药材得到一定的保护,而且对了解寄生植物在荒漠地区等极端严酷环境中的适应机制具有重要的生态学意义。该研究采用形态学观察结合常规石蜡切片法,对锁阳和肉苁蓉分别在各自寄主植物上的寄生方式进行了研究。结果表明:(1)锁阳的营养繁殖体在寄主植物根部呈串状分布,与寄主植物的连接方式属于非末端寄生;锁阳的吸器侵入寄主根系韧皮部和木质部的一部分区域,但是韧皮部和木质部大部分区域未被锁阳吸器占据,即有部分营养物质被锁阳“截取”。(2)肉苁蓉在其肉质茎基部长出新的芽体,与寄主植物的连接方式属于末端寄生;肉苁蓉的吸器侵入寄主根韧皮部和木质部全部区域。因此,锁阳寄生后,被寄生的寄主根依然能够向前生长,具有正常的功能;肉苁蓉寄生后,寄生点的寄主根失去根系的正常功能,成为一个为肉苁蓉生长发育提供营养物质的“输送通道(Transport channel)”。
寄生植物, 锁阳, 肉苁蓉, 寄生方式
锁阳(Cynomoriumsongaricum)为锁阳科(Cynomoriaceae)锁阳属植物,肉苁蓉(Cistanchedeserticola)为列当科(Orobanchaceae)肉苁蓉属植物。文献记载均为全寄生植物,锁阳的寄主植物为白刺属(NitrariaL.)、红砂属(ReaumuriaL.)、猪毛菜属(SalsolaL.)、柽柳属(TamarixL.)(Chen & Michele,2007)以及霸王(Sarcozygiumxanthoxylon)(陈叶等,2011)、多裂骆驼蓬(Peganummultisectum)(王进等,2011),肉苁蓉的寄主植物为梭梭(Haloxylonammodendron)及白梭梭(Haloxylonpersicum)(Zhang & Nikolai,1998)。锁阳分布于我国甘肃、内蒙古、宁夏、青海、陕西和新疆(Chen & Michele,2007);肉苁蓉分布于甘肃、内蒙古、宁夏和新疆(Zhang & Nikolai,1998)。二者均分布于荒漠地区。寄生植物可以加速有机物的分解,促进养分物质循环(Quested et al,2002)。因此,锁阳和肉苁蓉的寄生对荒漠地区生态系统具有一定的积极作用。
锁阳和肉苁蓉均为补益类药物,具有补肾阳、益精血、润肠通便的作用(国家药典委员会,2010)。肉苁蓉为濒危物种(傅立国,1992),锁阳为易危物种(中华人民共和国环境保护部和中国科学院,2013),肉苁蓉为国家二级重点保护野生植物(第二批),锁阳为新疆维吾尔自治区、内蒙古自治区、青海省重点保护野生植物。锁阳和肉苁蓉作为两种补益类中药,由于其不断的深入研究及临床应用,其药用价值和经济价值越来越受到人们的关注。但是肉苁蓉用药来源大部分来自野生药材的采挖,锁阳用药则完全依赖于野生药材的采挖,在经济利益的驱动下,人们盲目采挖,致使锁阳和肉苁蓉产地生态环境受到严重破坏。虽然目前已有关于锁阳和肉苁蓉人工栽培(刘桂荣等,2014;孙永强等,2008;屠鹏飞和郭玉海,2015)的报道,但是锁阳还未得到大规模的种植,肉苁蓉种植则存在很多盲目性。为拯救和保护锁阳和肉苁蓉资源,急需在两者的生态适宜生长区(陈君等,2007;黄林芳等,2010)进行锁阳和肉苁蓉的人工种植。
李天然等(1994)发现锁阳存在特殊的“类生殖根状物组织(Stolon-like-organ)”,并认为其实质是寄生于寄主根内的具有分生和繁殖发育成锁阳新个体能力的细胞组织;苏格尔和包玉英(1999)又将这些组织称为“类生殖根状细胞”,它可在寄主根韧皮部内向四周分裂扩张,形成“类生殖根状细胞群”,在寄主根部一定部位膨大突出根表皮外形成新的锁阳芽体。但是锁阳的“类生殖根状细胞”与寄主植物根的维管组织的连接方式还未见相关报道。了解锁阳和肉苁蓉分别与各自的寄主植物的寄生方式可以在采挖药材时采用适当的方法,这对保护锁阳和肉苁蓉野生资源具有重要的意义。
1 材料与方法
1.1 研究地概况
采样地马鬃山地区、肃州区和明花乡均为戈壁荒漠地区,气候干旱,植被稀少。马鬃山地区年均降水为85.2 mm,年均蒸发量为3 072.9 mm,年均气温为3.9 ℃(汪之波等,2009)。植物主要有梭梭、红砂(Reaumuriasongarica)、泡泡刺(Nitrariasphaerocarpa)、多种柽柳(Tamarixspp.)、芦苇(Phragmitesaustralis)、膜果麻黄(Ephedraprzewalskii)等。
1.2 研究材料
锁阳于2014年5月和2015年5月采自甘肃省肃北蒙古族自治县马鬃山地区(41°54′ N,97°7′ E)、酒泉市肃州区(39°31′ N,98°48′ E)、张掖市肃南裕固族自治县明花乡(39°30′ N,99°6′ E)和张掖市甘州区(38°59′ N,100°25′ E),锁阳均为野生状态,共观察52例,寄主植物均为泡泡刺,采样时均处于出土或开花阶段,采样时均在5月,各地取样时间间隔不超过10 d。
肉苁蓉于2014年5月和2015年5月采自甘肃省肃北蒙古族自治县马鬃山地区(41°54′ N,97°7′ E)、酒泉市肃州区(39°40′ N,98°29′ E,栽培肉苁蓉)和张掖市甘州区(38°54′ N,100°40′ E),人工种植肉苁蓉共观察23例,采样时间均为种植后第三年,野生肉苁蓉共观察65例,寄主植物均为梭梭。野生和种植药材采样时均处于出土或开花阶段。取样时间均在5月,各地取样时间间隔不超过10 d。
取样地相关信息如表1所示。采用整体挖掘法进行观察和拍照(Nikon D700),相关材料用70% F.A.A固定液进行固定。
1.3 研究方法
采用常规石蜡切片法对固定好的材料进行切片,用番红-固绿复染法进行染色,用奥林巴斯研究级倒置荧光显微镜(Olympus IX53)进行组织切片观察与照相。
表 1 取样地位置信息
Table 1 Location information of sampling sites
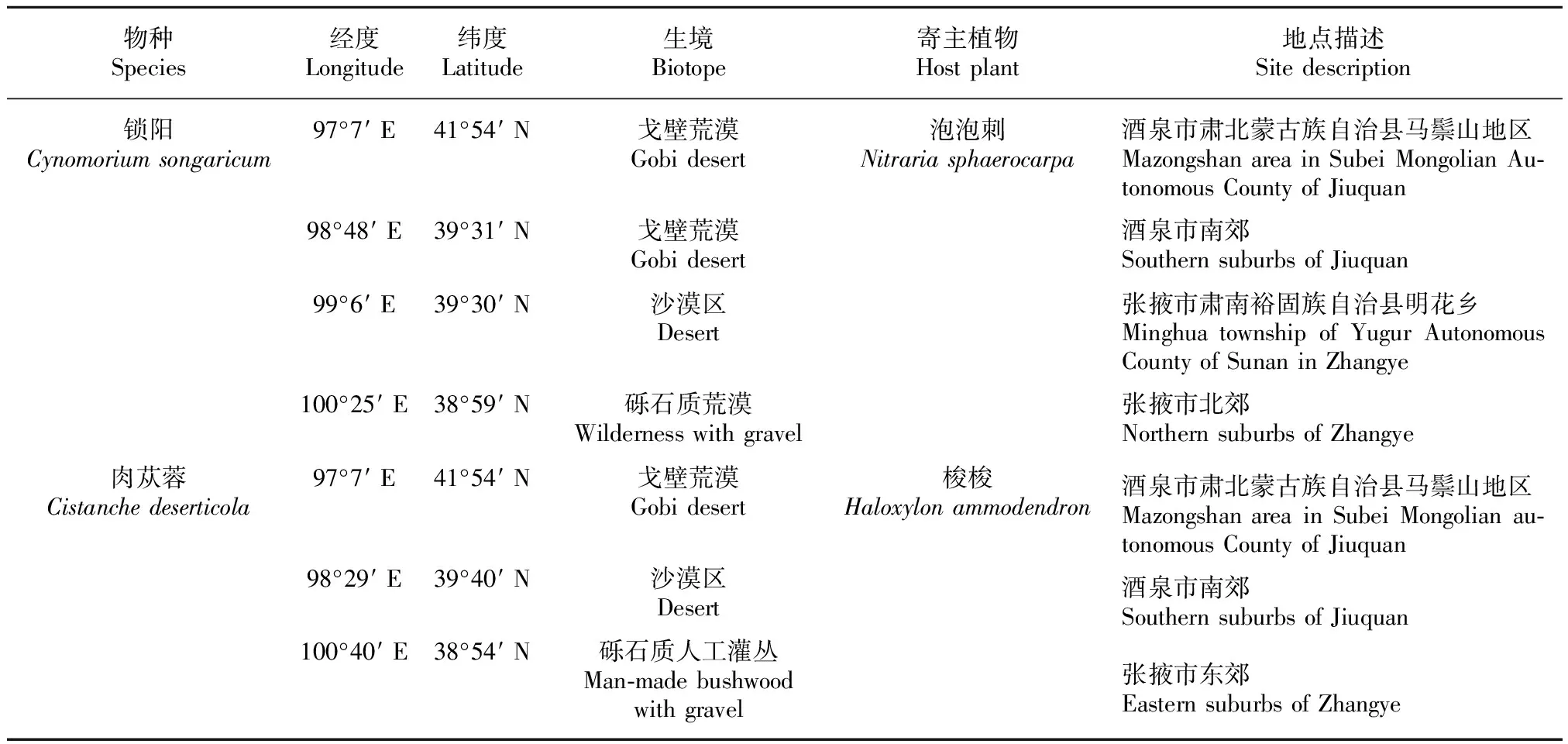
物种Species经度Longitude纬度Latitude生境Biotope寄主植物Hostplant地点描述Sitedescription锁阳Cynomoriumsongaricum97°7′E41°54′N戈壁荒漠Gobidesert98°48′E39°31′N戈壁荒漠Gobidesert99°6′E39°30′N沙漠区Desert100°25′E38°59′N砾石质荒漠Wildernesswithgravel泡泡刺Nitrariasphaerocarpa酒泉市肃北蒙古族自治县马鬃山地区MazongshanareainSubeiMongolianAu⁃tonomousCountyofJiuquan酒泉市南郊SouthernsuburbsofJiuquan张掖市肃南裕固族自治县明花乡MinghuatownshipofYugurAutonomousCountyofSunaninZhangye张掖市北郊NorthernsuburbsofZhangye肉苁蓉Cistanchedeserticola97°7′E41°54′N戈壁荒漠Gobidesert98°29′E39°40′N沙漠区Desert100°40′E38°54′N砾石质人工灌丛Man⁃madebushwoodwithgravel梭梭Haloxylonammodendron酒泉市肃北蒙古族自治县马鬃山地区MazongshanareainSubeiMongolianau⁃tonomousCountyofJiuquan酒泉市南郊SouthernsuburbsofJiuquan张掖市东郊EasternsuburbsofZhangye
2 结果与分析
2.1 形态学观察
对52例采自不同地点的锁阳进行形态学观察后均发现:锁阳在寄主的根上呈串状分布,有连续的许多芽体,锁阳与寄主的寄生方式属于非末端寄生(图版Ⅰ:A)。对65例采自不同地点的野生肉苁蓉进行观察后均发现:肉苁蓉与寄主单点寄生,寄生点的寄主根系呈膨大状,肉苁蓉的肉质茎基部完全包围寄主根末端,寄生方式属于末端寄生(图版Ⅰ:B),23例人工种植肉苁蓉的寄生方式与野生肉苁蓉完全相同(图版Ⅰ:C)。
2.2 解剖学观察
锁阳与寄主植物根系的解剖学结果显示:锁阳的吸器侵入了寄主植物的韧皮部和木质部(图版Ⅱ)。图版Ⅱ:A为未寄生锁阳的寄主根,显示锁阳的寄主根系的木质部有呈放射状的髓射线;图版Ⅱ:B为寄生了锁阳的寄主根(箭头所指为锁阳的吸器),锁阳的吸器侵入寄主的韧皮部和木质部,但韧皮部和木质部的大部分区域未被侵占;图版Ⅱ:C为吸器的局部放大图,显示锁阳的吸器侵入寄主植物根的木质部。由锁阳的吸器只侵占寄主根韧皮部和木质部局部区域的现象来看,锁阳只从寄主根系局部区域吸收有机营养物质、水分及矿质养分,即寄主根的营养物质在输送过程中有部分被锁阳“截取”。
肉苁蓉的吸盘将寄主植物的根部完全包围,吸器侵入了寄主植物的韧皮部和木质部(图版Ⅲ)。连续切片结果显示,肉苁蓉将寄主的木质部包围后逐渐解离,连接部位的寄主根木质部充满肉苁蓉的环纹导管(图版Ⅲ)。肉苁蓉寄生后,其肉质茎完全包围寄主根末端, 寄生点的寄主根丧失吸收水分等正常功能,而只是供给肉苁蓉生长发育所需的营养物质,变为肉苁蓉吸收水分、无机盐和光合作用产物的一个“输送通道(Transport channel)”。

图版 Ⅱ 锁阳与寄主连接部位的切片观察 A. 未寄生锁阳的寄主根; B. 寄生了锁阳的 寄主根 (箭头所指为锁阳的吸器); C. 吸器的局部放大图。 Plate Ⅱ Relationship of Cynomorium songaricum and its host A. Root of host which is not parasitic of C. songaricum; B. Root of host which is parasitic the C. songaricum (Arrow point is haustorium of C. songaricum); C. Partial enlarged view of haustoria.

图版 Ⅲ 肉苁蓉与寄主的寄生关系 虚线框内为肉苁蓉寄主植物的木质部; A2,B2,C2,D2,E2,F2分别为A1,B1,C1,D1,E1,F1的局部放大图。 Plate Ⅲ Relationship of Cistanche deserticola and its host The dotted box is the xylem of host plant of C.deserticola; A2, B2, C2, D2, E2 and F2 are the enlarged section of A1, B1, C1, D1, E1 and F1 respectively.
3 讨论
在荒漠等严酷恶劣的环境(王洪义等,2005)和限制性资源条件(Verburg & Grava,1998)下营养繁殖往往占据优势,营养繁殖对种苗更新困难时维持种群的存活有重要的作用,克隆植物须生长到一定大小才可能开花,因此有性生殖比营养繁殖需要更高的投资(王洪义等,2005)。荒漠地区环境条件恶劣,锁阳和肉苁蓉的种子必须从地面到达地下才能萌发与寄主植物建立寄生关系,因此营养繁殖对锁阳和肉苁蓉维持种群的生存具有重要的意义。
锁阳的寄生方式属于非末端寄生,被寄生后锁阳的吸器侵入寄主根韧皮部和木质部部分区域,但韧皮部和木质部的大部分区域未被吸器占据,因此锁阳寄生后,研究寄主根系依然能向前生长,具有正常功能。肉苁蓉寄生后,被寄生的寄主根失去正常功能,变为一个为肉苁蓉生长发育提供营养物质的“输送通道”,因此若寄生发生在寄主幼苗时期,很容易由于肉苁蓉过度吸收寄主的营养物质而造成寄主死亡。锁阳和肉苁蓉虽然都同属寄生植物,又都分布于荒漠中,但是却具有不同的两种寄生方式,这种不同一方面反映了植物在进化过程中的复杂性,另一方面也说明了植物在生长繁衍过程中为了获得物种存活度最大化而采取了最适合自身的繁殖方式。
锁阳的寄生对寄主的伤害是非致命的,寄生关系发生后寄主根系可以继续向前生长,这尤其对处于幼苗期的寄主植物的存活较为有利;肉苁蓉则不同,末端寄生方式完全吸收了寄生点的寄主根系所有营养物质,此时如果寄主植物尚处于幼苗阶段,则有可能造成寄主植物的营养物质来源中断而死亡,这对寄主梭梭或肉苁蓉都不利。一般情况下,肉苁蓉只在梭梭的一个根系上寄生,表明寄生肉苁蓉后的梭梭其株高、地径和冠幅等均普遍小于未寄生肉苁蓉的梭梭,总生物量比未寄生肉苁蓉的梭梭减少了49%,同时枯枝生物量比未寄生肉苁蓉的梭梭高出一倍,而且在寄生肉苁蓉后梭梭根系中细根的比例增高(谭德远等,2004)。这说明肉苁蓉将梭梭产生的营养物质大量吸收,导致梭梭采取了维持生存的生态对策;在栽培肉苁蓉的考察中作者也发现:(1)一株梭梭上寄生了由营养繁殖形成的12个肉苁蓉,均即将开花,但其寄主梭梭已经枯死;(2)另一株梭梭上发现有4个由营养繁殖形成的肉苁蓉寄生,同一个栽植坑内生长的另一株梭梭没有被寄生肉苁蓉,导致两株梭梭外表呈现出明显的区别:寄生肉苁蓉的梭梭全株颜色枯黄,叶短小,而没有寄生肉苁蓉的梭梭颜色鲜绿,叶较长,这两个现象均说明肉苁蓉的寄生对梭梭本身的生长有极大损害(郑国琦,2005)。因此,肉苁蓉寄生对梭梭来说属于有害寄生。
锁阳寄生后寄主根系可以继续向前生长,根尖可以继续进行营养物质的吸收,水分、矿质养分和光合作用产物的运输过程并没有因为锁阳的寄生而中断,只是在运输过程中有部分被锁阳所吸收。因而锁阳的这种非致死性寄生方式比肉苁蓉的寄生方式更“温和”,因此这种寄生方式可以防止寄主营养物质被过度消耗而死亡。锁阳具有两种克隆繁殖方式:(1)锁阳种子萌发后产生初生吸器,初生吸器侵入白刺根部,一部分细胞分化为维管组织并与白刺根的维管束连接后发生寄生关系,在吸器远离寄主根系的一端发育形成锁阳芽体;另一部分细胞继续保持分生能力,在白刺根韧皮部内向四周生长,分裂繁殖形成带状:“类生殖根状组织”,这种组织在一定部位激烈分化后突出寄主根系形成锁阳的营养繁殖体(苏格尔和包玉英,1999)。(2)锁阳肉质茎基部产生白色的不定根,这些不定根向四周生长,遇到寄主根系后粘附于其上,并将具有繁殖功能的细胞植入寄主根系并形成“类生殖根状组织”,这些组织分化后形成锁阳的营养繁殖体(李天然等,1994)。锁阳的营养繁殖现象可以在不同的白刺植株之间发生,这与肉苁蓉只能在一株梭梭上寄生有所不同。因此可以合理推断:锁阳的寄生方式比肉苁蓉的寄生方式更为进化。
锁阳与肉苁蓉的肉质茎中贮存的营养物质主要用于供给生殖器官开花时所需,因此锁阳和肉苁蓉的最佳采收时期均在出土前期或出土期。野生状态下经常发现在往年挖过锁阳药材的坑中又生长出新的锁阳,而肉苁蓉由于只与寄主植物肉质茎基部连接,在采挖时特别容易与连接部位一起被挖下而造成单株植物灭绝的现象。因此在采挖时需特别注意不要挖断寄主根系,在距离肉质茎基部以上5~10 cm处使用工具(木质或塑料)水平切断,然后及时填埋采挖坑。这种采挖方式既可以保证肉苁蓉和锁阳药材的可持续利用,又可以最大限度地实现肉苁蓉和锁阳的物种保护。
CHEN J,XIE CX,CHEN SL,et al,2007. Suitability evaluation ofCistanchedeserticolabased on TCMGIS-Ⅰ [J]. Chin J Chin Mat Med,32(14):1396-1401. [陈君,谢彩香,陈士林,等,2007. 濒危药材肉苁蓉产地适宜性数值分析 [J]. 中国中药杂志,32(14):1396-1401.]
CHEN JR,MICHELE FA,2007. Flora of China:Vol.13 [M]. Beijing:Science Press ; St. Louis:Missouri Botanical Garden:434.
CHEN Y,LUO GH,WANG J,et al,2011. New host plant ofCynomoriumsongaricum[J]. Chin Trad Herb Drugs,42(5):1007-1008. [陈叶,罗光宏,王进,等,2011. 锁阳的一个新寄主植物 [J]. 中草药,42(5):1007-1008.]CHINESE PHARMACOPOEIA COMMISSION, 2010. Pharmacopoeia of China: Vol. 1 [M]. Beijing: China Medical Science Press:126,325. [国家药典委员会,2010. 中华人民共和国药典(一部) [M]. 北京:中国医药科技出版社:126,325.]
FU LG,1992. China plant red data book [M]. Beijing:Sciense Press:502-503. [傅立国,1992. 中国植物红皮书 [M]. 北京:科学出版社,502-503.]
HUANG LF,XIE CX,CHEN SL,et al,2010. Regional suitability evaluation on desert herbaCynomorii[M]. Chin Bull Bot,45(2):205-211. [黄林芳,谢彩香,陈士林,等,2010. 沙生药用植物锁阳产地适宜性的定量评价 [J]. 植物学报,45(2): 205-211.]LI TR,SU GE,LIU JH,et al,1994. Reproduction of parasitic drug flowering plantCynomoriumsongaricumin host plants [J]. J Inner Mongolia Univ (Nat Sci Ed),25(6):673-679. [李天然,苏格尔,刘基焕,等,1994. 寄生药用有花植物锁阳在寄主体内的繁殖 [J]. 内蒙古大学学报(自然科学版),25(6):673-679.]
LIU GR,MENG F,HE L,et al,2014. The cultivate techniques ofCynomoriumsongaricumparasited toNitrariaspp. in Hexi corridor [M]. Inform Agric Sci Technol,13:34-35. [刘桂荣,孟飞,何亮,等,2014. 河西走廊白刺接种锁阳栽培技术 [J]. 农业科技与信息,13:34-35.]
MINIETRY OF ENVIRONMENTAL PROTECTION OF THE PEOPLE’S REPUBLIC OF CHINA,THE CHINESE ACADEMY OF SCIENCES,2013. The Announcement of “China Biodiversity Red List on the International Day for Biological Diversity-Volume of Higher Plants” [EB]. [中华人民共和国环境保护部,中国科学院,2013. 关于发布《中国生物多样性红色名录-高等植物卷》的公告 [EB].]
QUESEDE HM,PRESS MC,CALLAGHAN TV,et al,2002. The hemiparasitic angiospermBartsiaalpinahas the potential to accelerate decomposition in sub-arctic communities [J]. Oecologia,130(1):88-95.
SU GE,BAO YY,1999. On the parasitic characteristics and artificial propagation ofCynomoriumsongaricumRupr. [J]. J Inner Mongolia Univ (Nat Sci Ed),30(2):83-87. [苏格尔,包玉英,1999. 锁阳(CynomoriumsongaricumRupr.)的寄生生物学特性及其人工繁殖 [J]. 内蒙古大学学报(自然科学版),
(Continueonpage1343)(Continuefrompage1317)
30(2):83-87.]
SUN YQ, TIAN YZ, SHENG JH, et al, 2008. Study on manual inoculation ofCistanchedeserticolaY. C. Ma in desert [M]. J Arid Land Resour Environ,22(9):167-171. [孙永强,田永祯,盛晋华,等,2008. 干旱荒漠区肉苁蓉人工接种技术研究 [J]. 干旱区资源与环境,22(9):167-171.]
TAN DY,GUO QS,WANG CL,et al,2004. Effects of the parasite plant (Cistanchedeserticola) on growth and biomass of the host plant (Haloxylonammodendron) [J]. For Res,17(4):472-478. [谭德远,郭泉水,王春玲,等,2004. 寄生植物肉苁蓉对寄主梭梭生长及生物量的影响研究 [J]. 林业科学研究,17(4):472-478.]
TU PF,GUO YH,2015. Cultivation techniques ofCistanchedeserticolaand its hostHaloxylonplants [M]. Beijing: Sciense Press: 5-7. [屠鹏飞,郭玉海,2015. 荒漠肉苁蓉及其寄主梭梭栽培技术 [M]. 北京:科学出版社:5-7.]
VERBURG R,GRAVA D,1998. Differences in allocation patterns in clonal and sexual offspring in a woodland pseudo-annual [J]. Oecologia,4:472-477.
WANG HY,WANG ZW,LI LH,et al,2005. Reproductive tendency of clonal plants in various habitats [J]. Chin J Ecol,24(6):670-676. [王洪义,王正文,李凌浩,等,2005. 不同生境中克隆植物的繁殖倾向 [J]. 生态学杂志,24(6):670-676.]
WANG J,LUO GH,CHEN Y,et al,2011. New record for China of host plant ofCynomoriumsongaricum-Peganummultisectum[J]. Chin J Chin Mat Med,36(23):3 244-3 246. [王进,罗光宏,陈叶,等,2011. 锁阳寄主植物的一个国内新记录——多裂骆驼篷 [J]. 中国中药杂志,36(23):3 244-3 246.]
WANG ZB,SUN JZ,MA QL,2009. Studies on seed plant diversity of Mazong Mountain in Gansu province [J]. J Arid Land Resour Environm,23(7):188-190. [汪之波,孙继周,马全林,2009. 甘肃马鬃山地区种子植物多样性研究 [J]. 干旱区资源与环境,23(7):188-190.]ZHANG ZY,NIKOLAI NT,1998. Flora of China:Vol.18 [M]. Beijing:Science Press and St. Louis:Missouri Botanical Garden:229-231.
ZHENG GQ,SONG YX,GUO SH,et al,2005. Study on variations of nitrogen nutrient substances ofCistanchedeserticolaandHaloxylonammodendronin differdent growth stages [J]. J Agric Sci,26(4):16-18. [郑国琦,宋玉霞,郭生虎,等,2005. 不同发育时期肉苁蓉和梭梭体内氮营养物质变化研究 [J]. 农业科学研究,26(4):16-18.]
Difference of parasitic form ofCynomoriumsongaricumandCistanchedeserticola
CHEN Jin-Yuan, CHEN Xue-Lin*, GUO Nan-Nan, MA Wen-Bing,DA Qing-Jing, ZHANG Hai-Xian
(CollegeofLifeSciences,NorthwestNormalUniversity, Lanzhou 730070, China )
CistanchedeserticolaandCynomoriumsongaricumare important tonic herbs of traditional Chinese medicine, but currently their wild resource is in danger of drying up in China because of excavation excessively or incorrectly method.Cistanchedeserticolais the endangered plant andCynomoriumsongaricumis a rare vulnerable plant of extinction in China. The study on the parasitic forms and characters of them can promote the cultivation of them, at the same time, protect the wild medicinal herbs resource, and it has the important ecological significance on the adapted mechanism of parasitic plants to desert environment. We compared the parasitic forms ofC.songaricumandCistanchedeserticolain their host plants respectively through morphology observation and routine paraffin section. The results were as follow: (1) The vegetative propagation buds ofCynomoriumsongaricumgrew on the root of host plant one by one and the parasitic form was Non-End parasitic. The haustorium ofC.songaricuminvaded into phloem and partial part of xylem of host plant, but most areas of phloem and xylem were not been occupied by haustoria, namely some nutrients of host were “interception” byC.songaricumon the process of transportation. (2)Cistanchedeserticolagrew new buds in the base of fleshy stem, the parasitic form was End parasitic. The haustorium ofC.deserticolainvaded into all the xylem and phloem of host plant. After been parasitized byCynomoriumsongaricum, the root of its host plant could grow continuely and have the regularly function. After been parasitized byCistanchedeserticola, the root of host plant losed normally function, and became a “Transport channel” to supply nutrition forC.deserticola.
parasitic plant,Cynomoriumsongaricum,Cistanchedeserticola, parasitic form
10.11931/guihaia.gxzw201512023
2015-12-23
:2016-05-10
中央财政中医药公共卫生专项(财社[2011]76号);国家中医药行业科研专项(201207002)[Supported by Specially Item of Chinese Medicine Public Health of Central Finance ([2011]76); Specially Item of National Traditional Chinese Medicine Industry for Scientific Research(201207002)]。
陈金元(1990-),男,甘肃永登人,在读硕士研究生,主要从事系统与进化植物学研究,(E-mail)18993176162@189.cn。
*通讯作者: 陈学林,博士,教授,主要从事系统与进化植物学和种子生态学研究,(E-mail)chenxuelin63@163.com。
Q948.9
A
1000-3142(2016)11-1312-07
陈金元, 陈学林, 郭楠楠, 等. 锁阳和肉苁蓉寄生方式的区别 [J]. 广西植物, 2016, 36(11):1312-1317CHEN JY, CHEN XL, GUO NN, et al. Difference of parasitic form ofCynomoriumsongaricumandCistanchedeserticola[J]. Guihaia, 2016, 36(11):1312-1317
