模拟氮沉降对亚热带栎属树种幼苗生长、生物量累积及光合特性的影响
2016-12-20王晓荣庞宏东郑京津王瑞文
王晓荣,潘 磊,庞宏东,郑京津,王瑞文
(湖北省林业科学研究院,湖北 武汉 430075)
模拟氮沉降对亚热带栎属树种幼苗生长、生物量累积及光合特性的影响
王晓荣,潘 磊,庞宏东,郑京津,王瑞文
(湖北省林业科学研究院,湖北 武汉 430075)
设置模拟氮沉降的控制试验,以NH4NO3作为外加氮源,模拟氮沉降变化,设置CK、N5、N15和N30(分别相当于氮沉降0、5、15和30 g·m-2a-1)4个处理,历时2 a,研究氮沉降对小叶栎、麻栎、栓皮栎、白栎和短柄枹栎幼苗生长、光合特性以及生物量累积和分配的影响。结果表明:(1)随着氮沉降浓度的逐渐增加,麻栎、栓皮栎和短柄枹栎的株高显著增高,中高浓度氮显著促进了栓皮栎和麻栎地径的增加。(2)栎类不同树种利用光能的程度存在较大差异,除麻栎外,小叶栎、栓皮栎、白栎和短柄枹栎幼苗的最大净光合速率、表观量子速率和光饱和点随氮处理水平增加呈先增加后降低的趋势,在N15水平达到最高。小叶栎和麻栎的光补偿点和暗呼吸速率随氮沉降浓度增加呈逐渐增加,栓皮栎和短柄枹栎呈先增加后降低,而白栎则为逐渐降低的趋势。除栓皮栎和麻栎外,氮沉降对白栎、短柄枹栎和小叶栎叶绿素α、叶绿素b和叶绿素α+b含量均表现出显著的促进作用,但不同处理间差异不显著。(3)栎属5个树种生物量均表现随氮沉降浓度增加而增加的趋势,不同栎类树种生物量分配格局受氮沉降影响存在差异。叶重比受影响均不显著,中高氮水平促进麻栎和小叶栎的枝重比以及短柄枹栎和栓皮栎的干重比明显增加。5种树种幼苗根冠比显著降低,且氮沉降浓度越高越明显。
栎属树种;亚热带;模拟氮沉降;幼苗生长;生物量累积;光合参数
近年来,氮沉降增加已经成为全球化的生态环境问题[1-2],我国已成为全球三大氮沉降集中区之一(分别为欧洲、美国和中国),国内许多地区存在高氮沉降现象[3]。因此,在氮沉降全球化的环境背景下,研究和预测氮沉降对森林生态系统的影响及其反馈就显得尤为重要,已成为许多研究学者关注的热点[4-5]。
氮素是植物体的基本构成元素之一,在植物生理过程中有着重要作用[6]。已有研究表明,大气氮沉降对植物生长、光合作用、光合产物分配、有机质分解、营养结构状况、抗逆性、生物多样性等诸多方面具有影响[2,5,7-8]。氮沉降在一定范围内能够刺激植物生长,提高植物生产力[3-4],但当氮素在植物体内积累到一定程度出现氮饱和现象时,氮的输入反而会抑制植物的生长,削弱树木对环境胁迫的抗逆性,甚至导致生态系统衰退[9-10],且氮输入量是否过量由树种因素决定[11]。一些学者关于亚热带氮沉降对木荷Schima superba、锥栗Castanea henryi、黄果厚壳桂Cryptocarya concinna、马尾松Pinus massoniana、青冈Cyclobalanopsis glauca、枫香Liquidambar formosana和秃瓣杜英Elaeocarpus glabripetalus等树种的生长、光合生理以及生物量累积特征进行了研究[2,12-13]。然而,关于亚热带地区植物在氮沉降升高趋势下的生理生态响应还较缺乏数据支持[13],亚热带森林植物对氮沉降的响应机制还存在很多不确定性,特别是针对栎属不同树种幼苗光合生理及生物量累积对氮沉降的响应还未见报道。
壳斗科栎属Quercus植物是我国亚热带最重要的地带性森林树种,在天然林和人工林中往往成为优势种和建群种,广泛分布于南方大部分省份,在维护区域生态安全起到了重要作用。本研究选取亚热带栎属中小叶栎Q. chenii、麻栎Q. acutissima、栓皮栎Q. variabilis、短柄枹栎Q.glandulifera var. brevipetiolata、白栎Q. fabri作为实验材料,研究模拟氮沉降的发生对亚热带栎类树种的生长、光合生理和生物量分配等过程的影响,为评价和预测氮沉降对亚热带森林影响程度提供科学依据,进一步为探讨全球变化背景下亚热带森林的管理和适应过程提供参考。
1 材料与方法
1.1 研究区概况
研究地点位于湖北省武汉市郊区的九峰国家森林公园,地理中心位置为东经114°29′50″,北纬30°31′4″。气候属亚热带季风性湿润气候,具有雨热同季、四季分明的特点。全年平均气温16.7 ℃,极端最高气温为41 ℃,极端最低气温-17.6 ℃,年日照时数1 600 h左右,无霜期235 d,年降水量1 200~1 400 mm。由于工农业和城市化的发展,本区域大气污染相当严重,武汉市郊区2005年降雨氮沉降总量为26.07 kg·hm-2a-1,2006年则上升为33.09 kg·hm-2a-1,与全国湿沉降最高值相近,远高于同期欧洲和美国的氮沉降量[8],已经成为我国典型的高氮沉降区域。
1.2 研究方法
1.2.1 实验材料
2011年9月中旬在九峰山国家森林公园周围采集栎属5个试验树种的种子,分别是小叶栎(XYL)、麻栎(ML)、栓皮栎(SPL)、短柄枹栎(DBBL)、白栎(BL)。采集的种子经过筛选、消毒,然后经沙藏处理后播于湖北省林业科学研究院温室内进行育苗。于2012年4月份选择株高与地径大小及生长状况基本一致的苗木,每个物种各选择28株,共计140株,移栽于直径17.2 cm、高15 m的生长盆内,用九峰山内土壤和草炭土按2∶1的体积比混合的土壤进行栽植,且放置在自然环境下生长3个月。
1.2.2 试验设计
待实验材料生长稳定后,设置4个氮沉降处理水平:CK(对照)、N5(低氮)、N15(中氮)、N30(高氮),每个处理7盆。于7月初开始,每月初和月中按处理分别向幼苗土壤浇灌NH4NO3溶液,溶液浓度为0、0.013 75、0.027 50、0.041 25 mol·L-1,每盆每次50 mL,保证所浇灌溶液不会造成烧苗现象发生,每年施氮6次,连续处理2 a,累计施氮量折算为氮沉降率分别是0、5、15、30 g·m-2a-1,不包括大气沉降的氮含量,这些施氮量是基于本地区的氮沉降量和国际上同类研究情况而确定[2,12]。
1.2.3 测定方法
(1 ) 株高、地径的测定
从2013年5月至2013年10月,每月月初对所有试验植株用直尺和游标卡尺分别测定株高和地径。
(2) 生物量的测定
于2013年10月底,应用全收割法,将幼苗整棵挖出清洗干净后,按叶、枝、干、根进行称量,之后放置在80℃烘箱中烘干至恒质量,分别称取其干质量,计算叶生物量、枝生物量、干生物量、根生物量以及叶重比、枝重比、干重比、根重比和根冠比。
(3) 叶绿素含量的测定
于2013年8月底,采用乙醇浸泡法进行叶绿素测定。取0.2 g新鲜叶片剪碎,长度2 mm左右,然后放入50 mL试管容量瓶中,用95%乙醇定容至刻度,避光浸提24 h,且每隔2 h震荡1次直至叶片完全发白为止,然后利用分光光度计在波长665和649测定吸光度,重复3次,计算出相应叶绿素的含量[14]。
1.2.4 光合作用的测定
于2013年9月初,利用LI-6400XT 便携式光合测定系统,选择晴天9:00~12:00进行植株的光响应测定。每个树种随机选3株幼苗,选择中上部第6~10片叶片,参照CO2浓度设定为400 μmol·m-2s-1,将温度控制在 25~ 30 ℃、湿度控制在55%,采用Li-6400红蓝光源做内置光源,设定光合有效辐射(PAR)为: 2 000、1 500、1 200、1 000、800、600、400、200、150、100、75、50、25、10 和 0 μmol·m-2s-1,测定其净光合速率。
光响应曲线的相关参数利用叶子飘等[15-16]的直角双曲线修正模型求得,模型函数表达式为:
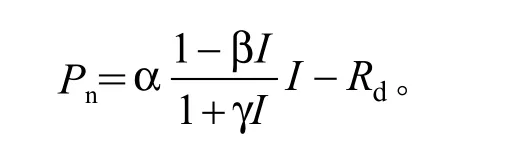
式中:Pn为净光合速率;α是光响应曲线的初始斜率,即表观光合量子效率;β和γ为系数;I为光合有效辐射;Rd为暗呼吸速率。植物的光饱和点(L)和最大净光合速率(Pnmax)分别由和求得。
1.3 数据处理
数据统计均采用SPSS 16.0和Excel 2007软件进行处理和分析,根据各项的测定指标,将氮处理水平作为独立因素,以采用单因素方差分析(one-way ANOVA),且进行多重LSD比较,以检验数据之间的差异显著性(P=0.05);采用SigmaPlot 10.0作图。
2 结果与分析
2.1 氮沉降对栎属不同树种株高和地径的影响
由图1所示,经过连续2 a氮沉降处理,栎属不同树种株高和地径对氮沉降都发生了显著变化。麻栎、栓皮栎和短柄枹栎的株高随氮沉降浓度逐渐增加,其中麻栎分别增加了6.53%、6.75%、39.29%,栓皮栎增加了3.24%、29.04%、50.72%,短柄枹栎增加了28.39%、58.03%、94.80%。氮沉降处理对短柄枹栎和白栎地径生长未造成显著性影响,小叶栎在N5水平较对照增加了66.51%,麻栎在中高氮水平分别增加了23.65%、28.08%,栓皮栎在N30水平增加了89.91%。

图1 不同氮浓度处理下栎属不同树种形态特征Fig. 1 Morphological characteristics of different tree species of Quercusunder different N treatments
2.2 氮沉降下栎属不同树种光响应参数特征
已有研究表明,最大净光合速率、表观量子效率、光饱和点、光补偿点等光响应参数是研究植物光合生理特性以及适应环境变化能力的重要指标[17]。如表1所示,各氮处理水平对不同物种光能利用能力产生了不同程度的影响。随着氮沉降浓度的增加,最大净光合速率表现为先增加后降低的趋势。其中,栓皮栎最大净光合速率在N5和N15水平较对照增加了108.37%和141.24%,短柄枹栎在N5、N15、N30水平较对照分别增加了37.68%、61.69%、58.21%,小叶栎在N15水平较对照提高了18.62%。表观量子效率反映了叶片对光能的利用情况,尤其是对弱光的利用能力,其值越大表明植物吸收与转换光能的色素蛋白复合体可能越多,利用弱光的能力越强[18]。除小叶栎在N30达到最高外,其他树种均在N5和N15达到最高值,然后出现一定程度下降;氮沉降未对麻栎和小叶栎的光饱和点造成显著影响,白栎、短柄枹栎、栓皮栎则呈现在N15达到最大后出现降低的趋势,其中白栎和栓皮栎在N5和N15水平分别较对照63.44%、25.58%和69.31%、61.74%,说明白栎、短柄枹栎、栓皮栎在高浓度氮沉降会发生一定光抑制。光补偿点和暗呼吸速率随着氮沉降浓度的增加,麻栎和小叶栎表现为逐渐上升的趋势,其中麻栎未到达显著水平(P≥0.05),而小叶栎光补偿点和暗呼吸速率在N30较对照增加了112.14%和141.52%,说明高氮沉降致使其消耗物质和能量较大,不利于生物量的明显积累。栓皮栎和短柄枹栎则表现为先升高后降低的趋势,光补偿点均在N15达到最高值,较对照分别增加了43.91%和76.53%,暗呼吸速率则在N5达到最高值后出现降低,栓皮栎在N5和N15分别较对照增加了76.1%和61.69%。白栎光补偿点和暗呼吸速率随氮浓度的增加表现为逐渐降低的趋势,但均未达到显著水平(P≥0.05)。

表1 不同氮浓度处理下栎属不同树种光合参数†Table 1 Photosynthetic parameters of different tree species of Quercusunder different N treatments
2.3 氮沉降对栎属不同树种光合色素含量的影响
光合色素尤其是叶绿素在植物光合作用中对光能的吸收、传递和转化起着极为重要的作用,叶绿素含量的多少在一定程度上影响植物的光合速率[18]。除栓皮栎和麻栎的叶绿素含量未对氮沉降表现出明显变化外,白栎、短柄枹栎和小叶栎叶绿素α、叶绿素b和叶绿素α+b均表现出显著的促进作用(P≤0.05),但不同氮处理间未表现出显著性差异(P≥0.05)。

图2 不同氮处理下栎属不同树种光合色素含量Fig. 2 Photosynthetic pigment contents of different tree species of Quercus under different N treatments
2.4 氮沉降对栎属不同树种生物量累积和分配的影响
由表2可以看出,随氮沉降浓度的增加,栎属5个树种生物量均表现出了增加的趋势,且氮沉降浓度越高生物量累积较对照增加的越多。其中,麻栎为19.28%、49.46%、93.51%,栓皮栎为17.82%、48.53%、90.67%,短柄枹栎为91.94%、59.65%、117.05%, 白 栎 为 6.99%、47.11%和121.96%,仅有小叶栎未达到显著水平。从各树种生长器官而言,高氮沉降浓度促进麻栎、栓皮栎和白栎的叶、枝、茎和根分别增加了108.21%、210.27%、131.04%、61.29%;139.80%、45.61%、236.97%、48.47%和140.98%、38.74%、41.80%、163.49%。可见,大气氮沉降可以明显促进植物生物量的累积,但不同树种幼苗利用氮素进行生产力转化的能力存在差异。
不同器官的有机物分配比重是植物对外界环境响应和自身调节的结果,往往会根据环境改变处于动态变化之中,以使自身处于最佳的生理生命状态,提高适应能力[19]。如表2所示,不同氮沉降处理对各树种生物量分配比例产生了影响,叶重比具有一定程度的增加,但均未达到显著水平;N15促进麻栎和小叶栎枝重比的显著增加98.77%和72.933%(P≤0.05);N30促进短柄枹栎、栓皮栎干重比显著增加(P≤0.05),增加了124.8%、54.41%,而短柄枹栎、麻栎、栓皮栎根重比显著降低,降低了44.11%、19.73%、24.11%。 同时,植物根冠比反映了生物量在地上和地下部分之间的分配情况,经常作为衡量植物生长状态的重要指标。本结果表明,氮沉降均在一定程度上导致根冠比的降低,但不同树种对不同浓度氮沉降反映不一致,除小叶栎和白栎对氮处理水平不存在差异外,N30促使麻栎和短柄枹栎根冠比分别降低42.87%和118.8%;栓皮栎根冠比在N15和N30分别降低32.39%和74.07%,说明高氮沉降对根的生长不利,明显抑制了地下生物量的积累。
3 结论与讨论
氮素是植物生长过程中最重要的养分限制因子,直接影响植物体内叶绿素和可溶性蛋白水平及光合酶类的合成与活性,从而调节光合作用,影响了植物的形态建成、生长发育和生产力变化[12,18]。本研究结果表明,外源氮素增加显著促进了栎属不同树种株高、地径的增加,且随供氮量的增加表现为一定程度的逐渐增加。但是,栎属不同树种生长对氮沉降响应存在明显差异,比如白栎株高和地径均未受到显著影响,而其他树种则变化明显,这可能是因为不同物种对所处氮环境的响应能力有所差异,过量的氮引起了植物营养失衡,或者是自我隐蔽产生的负效应抵消了氮含量增加的效应[20]。同时,氮沉降显著提高小叶栎、白栎和短柄枹栎的叶绿素含量,栓皮栎和麻栎则可能由于氮沉降下较大的生长量而发生的稀释效应致使叶绿素含量与对照无差异,说明氮沉降提高了栎类树种捕获光能的能力,有利于光合作用的高效进行。随着氮沉降浓度的增加,小叶栎、栓皮栎和白栎最大光合速率表现为先增加后降低,以N15水平最高,而麻栎和短柄枹栎则逐渐增加,可见氮沉降可以促进栎属树种净光合速率的增加,与叶绿素含量的增加有关。植物叶片的光饱和点、光补偿点反映了植物对光照条件的需求,植物的光补偿点越低,越能在弱光条件下顺利进行光合作用,较高的光饱和点说明植物具有利用较高光强的能力[3,20,21]。本研究中,除白栎幼苗外,不同氮沉降水平致使栎属不同树种光饱和点、光补偿点和暗呼吸速率均表现为增加的趋势,可见氮沉降促使其他栎类树种光合能力的加强,也降低了其对弱光环境的适应性,易发生光抑制,物质能量消耗也同样有所增加。
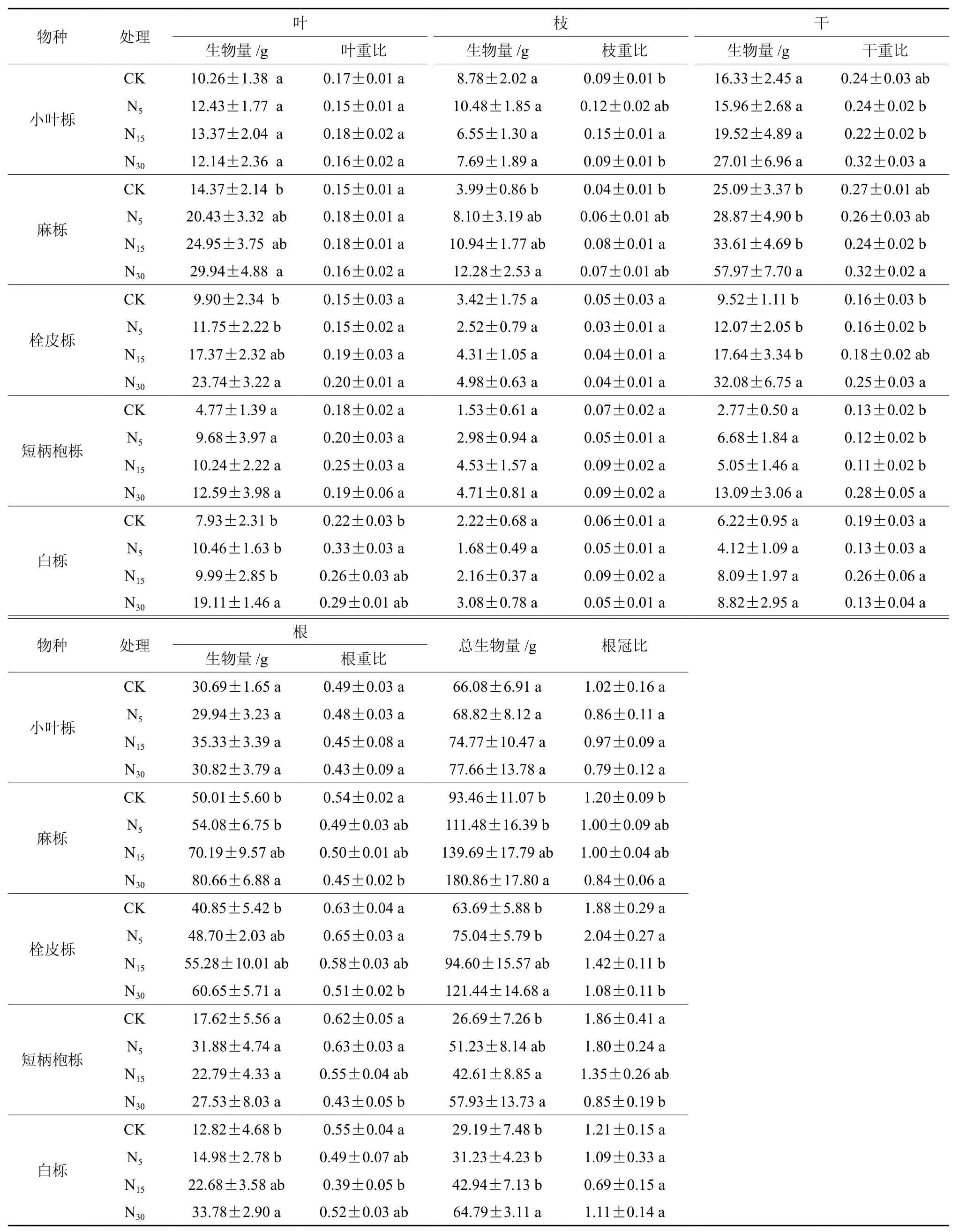
表2 不同氮浓度处理下栎属不同树种生物量累积及分配格局Table 2 Biomass accumulation and allocation of different tree species of Quercus under different N treatments
大量研究表明,氮输入的增加会明显促进亚热带森林植物的生长,导致生物量累积的增加[2,12,13,21],而Matson等[4]认为绝大部分亚热带森林植物的生长不受氮的限制,氮沉降的增加可能不会促进植物生长,本研究结果与前者结论相一致,氮沉降显著促进栎类树种全株生物量和各部分生物量的增加。随着氮沉降浓度的增加,各树种生物量均表现为逐渐增加,说明该区域栎类树种生长还未受到氮限制,高氮浓度仍可促进其生物量的累积,也可能是实验时间较短的缘故。从氮沉降对各树种体内生物量分配比例而言,各物种在不同氮沉降下对光资源的竞争策略和竞争力间存在着显著差异[12],且对氮沉降的响应程度具有很大变异性[13]。氮沉降增加均一定程度上促使叶重比的增加,虽然均未到达显著水平,但仍可看出,叶片作为植物的光合同化器官,是进行光合作用获取资源的主要构件,氮素累积后首先要供应到叶片,这有利于植物光合产物的累积,枝重比则由于所占地上生物量不高而变化不明显。高氮沉降明显促进了各树种茎重比的增加,而低中氮沉降则不明显,说明氮素在植物体内积累足够多后,解除了植物的营养限制,加速了植物代谢,使同化产物更多地向碳“汇”器官(茎和根)运输[22]。另外,地上生物量的分配比例的增加是以降低地下生物量的分配比例为代价的[12]。本研究中,不同栎属树种的根重比和根冠比随氮浓度增加而逐渐降低,表明氮沉降对植物根系生长不利,导致幼苗生物量向根部分的比例增加,这与李德军等[2]和闫慧等[21]对黄果厚壳桂和荷木幼苗的研究结果相一致,这是因为根冠比的下降是由于氮增加的同时增加了地上和地下的生物量,但是地下生物量增加的量少于地上生物量[12]。
综上分析可知,氮沉降对栎属不同树种生长和生理生态等方面造成较大的影响。中高氮水平显著促进了麻栎、栓皮栎、小叶栎和短柄枹栎株高和地径的增长,且高浓度氮处理更显著,但白栎则不明显。中低氮浓度处理促进了栓皮栎和短柄枹栎最大光合速率、表观量子效率、光饱和点、光补偿点和暗呼吸速率显著增加,高氮浓度处理促进了小叶栎光补偿点和暗呼吸速率的增加,对麻栎和白栎光合参数没有显著作用。然而,氮沉降处理使得所有实验树种生物量累积不断提高,且随氮浓度增加而增加,麻栎为19.28%、49.46%、93.51%,栓皮栎为17.82%、48.53%、90.67%,短柄枹栎为91.94%、59.65%、117.05%,白栎为6.99%、47.11%和121.96%,仅有小叶栎未达到显著水平。高氮沉降浓度促进了栓皮栎、麻栎和白栎叶、枝、茎和根各生长器官的显著增加,同时也显著降低了栓皮栎、麻栎和短柄枹栎根冠比的显著降低。整体而言,栎类不同树种对氮沉降响应存在差异,栓皮栎、麻栎和短柄枹栎更敏感,而小叶栎和白栎则相对不敏感,可能是因为不同物种在适应环境变化的策略不同,是何种因素造成这种差异还需要进一步的研究。另外,已有大多数研究均认为,氮输入的增加会明显促进亚热带森林植物的生长,导致树种相对生长速率、总生物量、株高、地径、最大净光合速率和光合色素含量的增加,且以中等程度氮处理(10 g·m-2a-1NH4NO3)的效果最佳,高氮沉降浓度则会抑制植物的生长[2,21],而本研究均以高氮沉降更显著。主要原因可能是本实验采用盆栽幼苗进行控制实验,根系生长会受到明显的限制,且试验区域降水较充沛,雨水往往会冲刷外施氮素,造成土壤氮沉降浓度的稀释,影响了土壤养分的可利用性,这在一定程度上影响了实验结果的精确性。因此,研究大气环境变化下植物的响应机制,应该选择自然生长条件下栽植进行模拟,减少水分和生长空间的干扰因素,结合当地实际降水量和CO2浓度变化耦合研究是下一步准备进行的工作。
[1]Galloway J N,Dentener F J,Capone D G,et al. Nitrogen cycles:past,present,and future[J]. Biogeochemistry,2004,70(2): 153-226.
[2]李德军,莫江明,方运霆,等. 模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响[J]. 植物生态学报,2005,29(4): 543-549.
[3]张 蕊,王 艺,金国庆,等. 氮沉降模拟对不同种源木荷幼苗叶片生理及光合特性的影响[J].林业科学研究,2013,26(2): 207-213.
[4]Matson P A ,Lohse K A ,Hall S J. The globalization of nitrogen deposition: consequences for terrestrial ecosystems[J]. Ambio.,2002,31(2): 113-119.
[5]Reay D S,Dentener F,Smith P,et al. Global nitrogen deposition and carbon sinks[J]. Nature Geoscience,2008,1(7): 430-437.
[6]郑 威,闫文德,梁小翠,等. 氮添加对樟树林生长的影响[J].中南林业科技大学学报,2013,33(4): 34-37.
[7]吕超群,田汉勤,黄 耀. 陆地生态系统氮沉降增加的生态效应[J]. 植物生态学报,2007,31(2): 205-218.
[8]Zhao C S,Hu C X,Huang W,et al. A lysimeter study of nitrate leaching and optimurn nitrogen application rates for intensively irrigated vegetable production systems in Central China[J].Journal of soil and sediments,2010,10(1): 9-17.
[9]Neff J C,Townsend A R,Gleixner G,et al. Variable effects of nitrogen additions on the stability and turnover of organic carbon[J]. Nature,2002,419: 915-917.
[10]Nordin A,Strengbom J,Witzell J,et al. Nitrogen deposition and the biodiversity of boreal forests : implications for the nitrogen critical load[J]. Ambio.,2005,34(1): 20-24.
[11]赵 亮,周国逸,张德强,等. CO2浓度升高和氮沉降对南亚热带主要乡土树种及群落生物量的影响[J].应用生态学报,2011,22(8): 1949-1954.
[12]吴 茜,丁 佳,闫 慧,等. 模拟降水变化和土壤施氮对浙江古田山5个树种幼苗生长和生物量的影响[J]. 植物生态学报,2011,35(3): 256-267.
[13]李明月,王 健,王振兴,等. 模拟氮沉降条件下木荷幼苗光合特性、生物量与C、N、P分配格局[J]. 生态学报,2013,33(5):1569-1572.
[14]李志丹,韩瑞宏,廖桂兰,等. 植物叶片中叶绿素提取方法的比较研究[J]. 广东第二师范学院学报,2011,31(3): 80-83.
[15]YE Z P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica,2007,45(4): 637-640.
[16]叶子飘,赵则海. 遮光对三叶鬼针草光合作用和叶绿素含量的影响[J]. 生态学杂志,2009,28(1):19-22.
[17]王明怀,许秀玉,魏 龙,等. 木麻黄无性系光响应特性研究[J]. 华南农业大学学报,2011,32(2): 76-79.
[18]王建波,钟海秀,付小玲,等.氮沉降对小叶章光合生理特性的影响[J]. 中国农学通报,2013,29(7): 45-49.
[19]Magnani F,Mencuccini M,Grace J. Age related decline in stand productivity: The role of structural acclimation under hydraulic constraints[J]. Plant,Cell and Environment,2000,23(3):252-263.
[20]王 强,金则新,彭礼琼.氮沉降对乌药幼苗生理生态特性的影响[J]. 应用生态学报,2012,23(10):2766-2772.
[21]闫 慧,吴 茜,丁 佳,等. 不同降水及氮添加对浙江古田山4种树木幼苗光合生理生态特征与生物量的影响[J]. 生态学报,2013,33(14):4226-4236.
[22]杜启然,雷静品,刘建锋,等. CO2浓度增加和施氮对栓皮栎幼苗生理生态特征的影响[J]. 应用生态学报,2014,25(1):24-30.
Effects of simulated nitrogen deposition on growth,biomass accumulation and photosynthetic characteristics responses of Quercus seedlings in Mid-subtropics of China
WANG Xiao-rong,PAN Lei,PANG Hong-dong,ZHENG Jing-jin,WANG Rui-wen
(Forestry Institute of Hubei Province,Wuhan 430075,Hubei,China)
To clarify the effects of nitrogen deposition on the growth,photosynthetic characteristics and biomass accumulation and distribution of Quercus chenii,Q. acutissima,Q. variabilis,Q. fabri and Q. glandulifera var. brevipetiolata seedlings,a two-year term control experiment were conducted,in which NH4NO3was added as the external nitrogen sources (four nitrogen depositions (0(CK),5 g·m-2a-1(N5),15 g·m-2a-1(N15) and 30 g·m-2a-1(N30)). The results show that (1) With increased nitrogen deposition levels,plants heights of Q. acutissima,Q. variabilis and Q. glandulifera var. brevipetiolata increased signi fi cantly,and medium and high concentrations of nitrogen deposition signi fi cantly promotes the diameter growth of Q. acutissima and Q. variabilis. (2) Light utilization efficiency of Quercus differed from each other species. Except Q. acutissima,maximum net photosynthetic rate,apparent quantum rate and light saturation point of Q. chenii,Q. variabilis,Q. fabri and Q. glandulifera var. brevipetiolata fi rstly increased,the peak was at the N15level,then decreased; Meanwhile,light compensation point and respiration rate of Q. chenii and Q. acutissima increased gradually; in contrast,light compensation point and respiration rate of Q. fabri decreased,while on Q. variabilis and Q. glandulifera var. brevipetiolata increased at fi rst and then decreased; Nitrogen deposition signi fi cantly favor the content increase of chlorophyll a,chlorophyll b and chlorophyll a+b in Q. fabri,Q. glandulifera var. brevipetiolata and Q. chenii,regardless of the nitrogen deposition levels. (3) As for the biomass accumulation in Quercus,regardless of the species,they gradually increased with the increase of nitrogen deposition level; however,biomass distribution in different species of Quercus signi fi cantly differed from nitrogen deposition levels,while the effects of nitrogen deposition levels on leaf weight ratio were insignificant; medium and high nitrogen levels increased the branches weight ratio of Q. acutissima and Q. chenii and significantly lead to higher stem weight ratio of Q. variabilis and Q.glandulifera var. brevipetiolata; root shoot ratio of the fi ve Quercus tree seedlings were signi fi cantly reduced,and a higher nitrogen deposition level accelerated the reduction.
Quercus; subtropics; simulation of nitrogen deposition; seedlings growth; biomass accumulation; photosynthetic parameters
S792.18
A
1673-923X(2016)01-0078-08
10.14067/j.cnki.1673-923x.2016.01.014
2014-04-02
国家林业公益性行业项目(201304308)
王晓荣,助理研究员,硕士 通讯作者:潘 磊,研究员;E-mail:panlei2008@126.com
王晓荣,潘 磊,庞宏东,等. 模拟氮沉降对亚热带栎属树种幼苗生长、生物量累积及光合特性的影响[J].中南林业科技大学学报,2016,36(1): 78-85.
[本文编校:谢荣秀]
