基于nrITS、psbA-trnH和DMC1序列中国短柄草族的系统进化和生物地理学研究
2018-07-24武泼泼张学杰张洛艳樊守金
武泼泼 张学杰 张洛艳 樊守金
(山东师范大学生命科学学院逆境植物重点实验室,济南 250014)
禾本科(Poaceae)植物种类众多,全世界约有700个属、10 000多种,起源时间最早可以追溯到约7 000年前的白垩纪(Cretaceous)[1],从温带气候为主的环境逐渐适应如今多种多样的生态环境,分布区覆盖地球陆地面积的五分之一[2]。短柄草族隶属于禾本科早熟禾亚科(Pooideae),全球约16种,主要分布在欧洲和亚洲大陆的温带地区,亚洲的高山以及从墨西哥到玻利维亚的美洲地区亦有分布[3],我国分布有5种,其中两种是中国特有。其总状花序和雀麦族的圆锥花序共同演化形成小麦族的穗状花序[4],和小麦族有着较近的亲缘关系。
该族植物具有重要经济价值、生态价值和科研价值。Bossolini等提出将短柄草和二穗短柄草作为模式植物,作为小麦(Triticumaestivum)和水稻(Oryzasativa)研究的桥梁[5]。短柄草是短柄草族多年生草本植物,优良牧草,分蘖能力强,水土保持能力良好。Steinwand等将短柄草开发为研究多年生禾草的模式植物[6]。二穗短柄草是短柄草族一年生草本植物,具有植株小,生长周期短,且染色体基因组小,结构也较为简单,较易遗传转化等特点,被开发为研究小麦、大麦(Hordeumvulgare)等麦类作物的模式植物。二穗短柄草基因组测序已经完成,基因组共5条染色体,测序获得拼接总长为272 MB的序列框架图,约包含25 532个编码基因[7]。根据测序结果,二穗短柄草与小麦基因组相似度很高,亲缘关系很近。以二穗短柄草为模式材料研究麦类作物,具有较高的准确性和效率,为解析小麦抗逆、高产、优质等重要性状提供了新的突破口。但是小麦族、雀麦族和短柄草族的系统学位置尚不明确。观点主要分为3类:一类认为短柄草属(Brachypodium)和雀麦族有较近的亲缘关系[8~9];一类支持短柄草属归属于小麦族[10~13];一类支持短柄草属独立成族[14~19]。短柄草族植物在我国主要分布于喜马拉雅山脉和横断山脉交界处,对揭示青藏高原的隆起和阐明植物的分布原因具有重要研究价值。因此,对短柄草族植物展开系统学和生物地理学研究是很有必要的。
核糖体rDNA片段ITS位于核糖体DNA内转录基因间隔区,是目前生物系统发育进化分析和DNA Barcoding研究中广泛应用的核基因片段。Alvarez&Wendel对发表在在几个著名的系统学和进化学期刊中的植物系统发育文章进行调查统计[20],共计调查244篇文献,在属及属下水平研究中涉及nrITS序列的文献占三分之二(66%),超过三分之一(34%)的系统发育树是只依靠nrITS序列进行构建的。
叶绿体基因psbA-trnH是一段非编码序列,长度约为450 bp,进化速率较快,且序列两端保守,有利于通用引物的设计,是目前常用的DNA Barcoding候选片段之一[21]。Li等人对植物条形码的研究综述中也表明psbA-trnH是目前应用最广泛的质体DNA条形码[22]。
单拷贝基因由于含有更多的遗传信息,逐渐受到分类学家的广泛重视[23~24]。DMC1是减数分裂相关的单拷贝核基因,在多倍体中独立遗传,在小麦族的多倍体体形成、属的分类研究中得到广泛应用。Tang等利用DMC1片段对四倍体小麦A基因组的起源进行研究。表明小麦A基因组来自于Triticumurartu,而不是一粒小麦(T.monococcum)[25]。
因此,本研究用叶绿体基因片段psbA-trnH,核糖体基因转录间隔区ITS,单拷贝核基因片段DMC1以及psbA-trnH+nrITS+DMC1联合序列对短柄草族及其相关属进行研究,以期为其系统学位置、属下关系和分歧时间提供翔实的证据。
1 材料和方法
1.1 样品采集
本研究共选取36份材料,其中短柄草属3种7个个体、雀麦属5种、小麦族7属23种,并选稻亚科(Oryzoideae)的稻为外类群。凭证标本均保存于山东师范大学标本馆(SDNU),相关信息见表1。基于ArcGIS软件制作的采集地点分布图见图1。

表1 实验材料及其来源Table 1 List of plant materials used in this study and their original resources

图1 标本采集地点分布图Fig.1 The distribution of sampling locations
1.2 DNA提取与PCR扩增
DNA材料来自于硅胶快速干燥保存的叶片,用改良的CTAB法[26]和DNA试剂盒提取DNA。扩增引物序列见表2。nrITS反应程序:95℃ 4 min;35个循环,分别为94℃ 30 s,52℃ 30 s,72℃ 90 s;最后72℃延伸10 min。psbA-trnH反应程序:94℃ 3 min;30个循环,分别为94℃ 30 s,55℃ 30 s,72℃ 45 s;最后72℃延伸10 min。DMC1反应程序:94℃ 5 min;35个循环,分别为94℃ 60 s,52℃ 60 s,72℃ 60 s;最后72℃延伸8 min。扩增反应在Bio-Rad T100 PCR仪上进行,PCR反应体系为20 μL:2*Taq PCR Master Mix(莱枫,上海)10 μL,上游和下游引物(华大,北京)各1 μL,DNA模板1 μL,7 μL ddH2O。扩增产物经琼脂糖凝胶电泳检验后交于北京华大基因公司测序。

表2 PCR引物名称及序列Table 2 The names and sequences of Primers
1.3 系统发育分析
使用Clustal X 1.8.3对目的序列进行比对分析,空位(Gap)做缺失处理,利用Gblocks 0.91b对基因序列进行剪切,手工调整完成校对。利用SquenceMatrix将Gblock得到的基因序列联合。通过Shortcut to DAMBE进行核酸饱和性检验,检验序列是否适合构建系统发育树;使用软件MrModeltest选择最佳核苷酸替换模型,ITS最佳模型为GTR+G,psbA-trnH最佳模型为HKY+I+G,DMC1最佳模型为HKY+G,联合序列在nex文件分区,模型对应如上。分别用MEGA6、PAUP 4.0、MrBayes 3.2.6进行最大简约法(MP)分析、最大似然法(ML)分析和贝叶斯法(BI)分析,构建3种进化树。采用启发式搜索对四组数据进行1 000次随机重复检验,TBR置换算法,得到的自展支持率用于评价最大似然树和最大简约树的拓扑结构的可靠性。使用MrBayes 3.2.6进行贝叶斯推理法分析,替换模型采用MrModeltest的结果,位点间差异比率采用γ分布比率差异,其中部分为不变位点,其余参数为默认值,采用马尔科夫—蒙特卡罗链数据模拟估算后验概率,数据运行1 000 000代,取样频率为1 000,总结树前舍弃25%老化样本,计算最优分支格局获得发育树的拓扑结构,以后验概率作为评估参数。
1.4 分化时间计算
用BEAST v1.8.4[27]进行分化时间计算。首先对联合序列进行似然比检验LRT(likelihood ratio test)确定该片段在各进化枝中是符合严格分子钟还是宽松分子钟。利用BEAST1.8.4中的宽松分子钟模型估算系统发育树上的每个节点的分歧时间。依据已有的化石记录和分子钟估计的时间选择两个节点进行标定:(1)选用稻亚科和早熟禾亚科的分歧时间49.6 Ma;(2)短柄草族和小麦族的分歧时间33 Ma。选择“Yule”进化模型和GTR+G模型进行运算。四条马尔科夫链(Markov Chain Monte Carlo)迭代计算100 000 000代,每1 000代进行一次取样,以保证参数posterior,prior和likelihood的有效取样量(ESS)大于200。用软件Tracer v1.6检查收敛程度,最初的10 000代做为老化样本舍去,用剩余样本构建时间尺度下的一致树。用FigTree v1.4.2进行处理,得到各分支的分化时间。
1.5 生物地理学分析
植物区系主要根据区系组成和植被类型进行划分,是一定区域内所有植物种类的总和,是在一定的自然地理条件下发展演化的结果。根据吴征镒的中国植物区系划分方案,本研究所用材料可划分为三个区域:A西北地区、B华北地区、C西南地区。基于BEAST1.8.4的trees文件,TreeAnnotator v1.8.4生成的tree文件以及统计的分布区信息,利用RASP(Reconstruct Ancestral State In Phylogenies)[28]软件对短柄草族、雀麦族和小麦族进行祖先分布区重建。
2 结果与分析
2.1 系统发育分析
基于nrITS、DMC1和联合序列构建的ML和BI系统发育树具有一致的拓扑结构,稻为外类群,小麦族(Ⅰ)和雀麦族(Ⅱ)聚为一支互为姐妹群,短柄草族(Ⅲ)和这一支互为姐妹群。基于psbA-trnH构建的ML和BI系统发育树,短柄草族单独聚为一支,雀麦族散布镶嵌在小麦族内,共同聚为一支。系统发育图如图2,图中支持率格式分别代表靴代值/后验概率。在nrITS、DMC1、psbA-trnH和联合序列分别构建的四个系统发育树中,短柄草族的支持率都是100/100,高度支持该属独立成族。雀麦族在nrITS、DMC1和联合序列分别构建的系统发育树中的支持率都是100/100,小麦族在nrITS、DMC1和联合序列分别构建的系统发育树中的支持率分别是37/97,58/94,78/100,支持小麦族和雀麦族的成立,但是基于psbA-trnH构建的系统发育树中,雀麦族镶嵌在小麦族内,支持率为100/100,暗示小麦族和雀麦族有着多种进化历史,可能发生过叶绿体基因的渐渗或转移等事件。
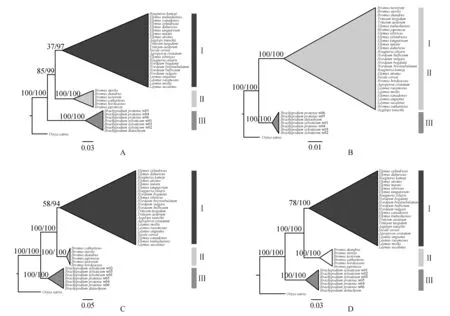
图2 小麦族、雀麦族和短柄草族系统发育图 A.nrITS;B.psbA-trnH;C.DMC1;D.nrITS+psbA-trnH+DMC1 Ⅰ(小麦族):节节麦、冰草、黑紫披碱草、加拿大披碱草、圆柱披碱草、披碱草、垂穗披碱草、老芒麦、麦薲草、Elymus tsukushiensis、布顿大麦草、短芒大麦草、球茎大麦、大麦、窄颖赖草、滨麦、大赖草、赖草、纤毛鹅观草、黑麦、普通小麦、圆锥小麦;Ⅱ(雀麦族):扁穗雀麦、双雄雀麦、大麦状雀麦、雀麦、旱雀麦;Ⅲ(短柄草族):二穗短柄草、草地短柄草、短柄草Fig.2 The Phylogenetic tree of Triticeae, Bromeae and Brachypodieae A.nrITS;B.psbA-trnH;C.DMC1;D.nrITS+psbA-trnH+DMC1 Ⅰ(Triticeae):Aegilops tauschii,Agropyron cristatum,Elymus atratus,Elymus canadensis,Elymus cylindricus,Elymus dahuricus,Elymus nutans,Elymus sibiricus,Elymus tangutorum,Elymus tsukushiensis,Hordeum bogdanii,Hordeum brevisubulatum,Hordeum bulbosum,Hordeum vulgare,Leymus angustus,Leymus mollis,Leymus racemosus,Leymus secalinus,Roegneria ciliaris,Secale cereale,Triticum aestivum,Triticum turgidum;Ⅱ(Bromeae): Bromus catharticus,Bromus diandrus,Bromus hordeaceus,Bromus japonicus,Bromus tectorum;Ⅲ(Brachypodieae):Brachypodium distachyum,Brachypodium pratense,Brachypodium sylvaticum
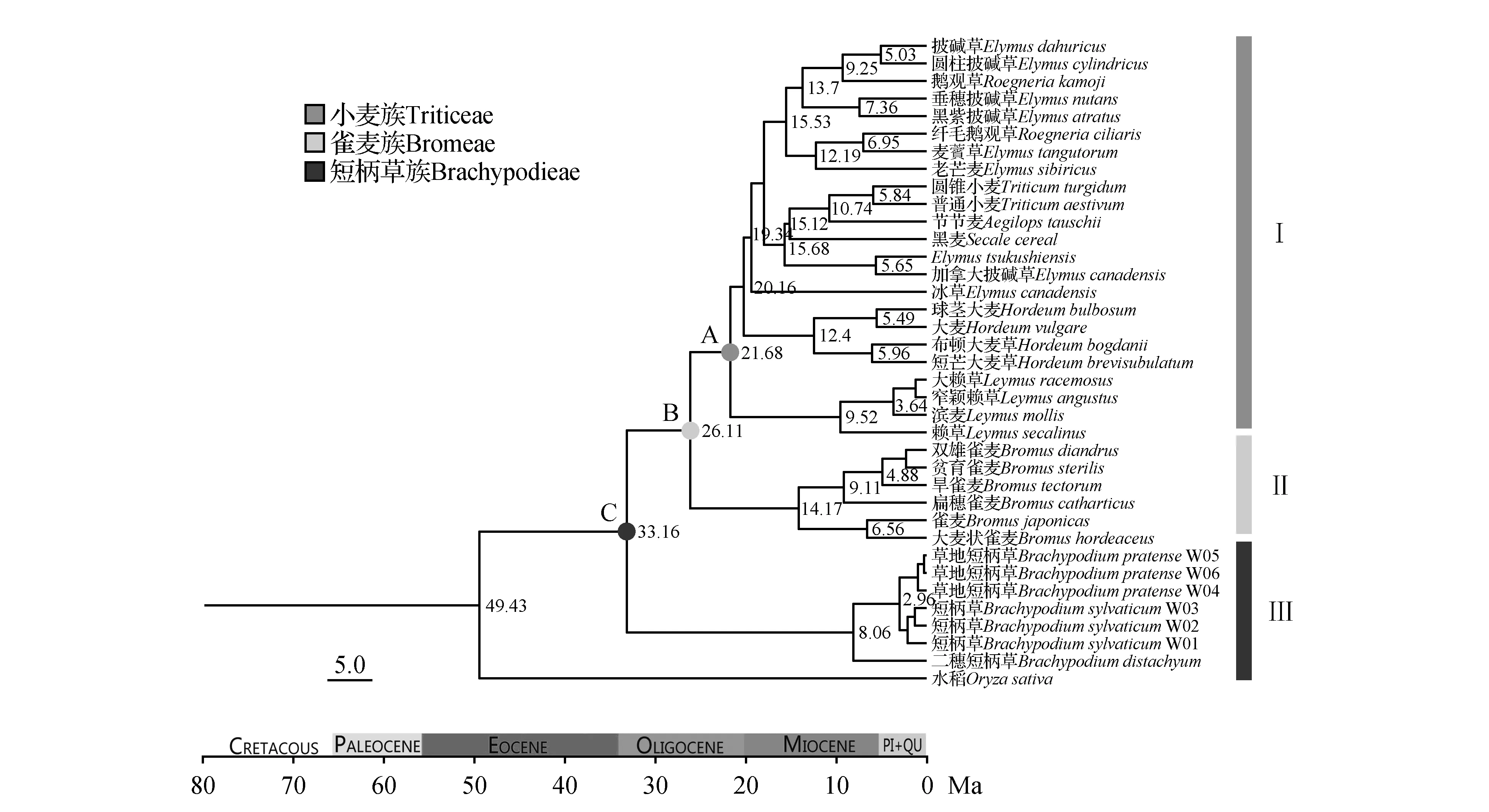
图3 短柄草族、雀麦族和小麦族各分支分歧时间图Fig.3 Divergence time of Brachypodieae,Bromeae and Triticeae
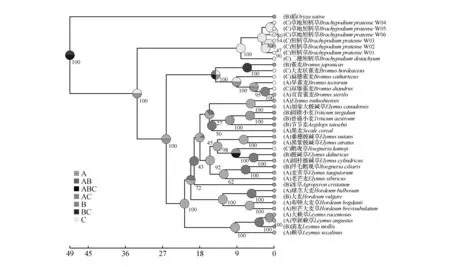
图4 祖先分布区重建 A.西北地区;B.华北地区;C.西南地区Fig.4 The reconstruction of ancestral state A. Northwest region;B. North China;C. Southwest region
2.2 分化时间计算
经LRT检验选用贝叶斯宽松分子钟计算,利用多个标定点对联合基因序列进行分析,这不仅能克服单个基因片段可能对分子钟造成的偏离影响,还可以弥补因为单个基因片段信息量不足所造成的缺陷。短柄草族、雀麦族和小麦族各分支分歧时间结果如图3所示。短柄草族其主要类群起源于33.16 Ma渐新世(Oligocene)的鲁培勒阶(Rupelian),基部的二穗短柄草在8.08 Mya分化形成;雀麦族其主要类群起源于26.11 Mya的渐新世的恰特阶(Chattian),基部类群在14.17 Mya分化形成;小麦族主要类群起源于21.68 Mya的中新世(Miocene)早期,其基部类群在9.52 Mya分化形成。鉴于各地质时间年龄值在不断修订,本研究的地质时期名称及其时间划分来自于国际地层委员会(International Commission on Stratigraphy)在2017年2月更新的数据[29]。
2.3 祖先分布区重建
通过对短柄草族、雀麦族、和小麦族的祖先分布区重建,RASP结果如图4。我国分布的短柄草族植物最近共同祖先分布在我国西南地区,小麦族和雀麦族组成小麦超族,最近共同祖先分布于我国西北地区。
3 讨论
3.1 短柄草族系统学位置分析
本研究基于nrITS、DMC1、psbA-trnH及其联合序列对短柄草族的系统学位置进行研究。基于nrITS、DMC1和联合序列的结果高度支持短柄草属成立短柄草族,这与Soreng[30~31]等人的结果相符合,也与Cheng等人[32]构建的中国维管植物生命之树相一致。基于叶绿体基因psbA-trnH的研究结果与其他基因片段及联合序列结果冲突,表明小麦族与雀麦族有着复杂的进化路线,且二者亲缘关系较近,支持小麦族和雀麦族组成小麦超族。综合以上结果,短柄草属单属成族,且最先分化出来,位于基部;其次,雀麦族和小麦族分化形成,二者相互为姐妹群,二者有着复杂的进化路线。
3.2 短柄草族生物地理学分析
BEAST的分析结果显示短柄草族位于三者的最基部,最先分化形成,分化时间是33.16 Ma,族下属内物种形成时间为二穗短柄草8.06 Ma,2.96 Ma短柄草与草地短柄草分化。在26.11 Ma雀麦族和小麦族开始分化,雀麦属在26.11 Ma分化,赖草属在21.68 Ma分化,大麦属在20.16 Ma分化,冰草属在20.16 Ma分化,披碱草属在19.34 Ma分化,黑麦属和小麦属在15.68 Ma分化。结合地理地质事件事件,青藏高原的隆升过程,可发现小麦族的属下物种的形成大多集中在青藏高原的第三阶段的隆升(7-13 Ma),在此阶段青藏高原在西北地区发生抬升事件[33];短柄草族的物种形成时间在3 Ma之后,而青藏高原在3.6 Ma以来经历了地表强烈提升和高原快速扩张,同时期的东亚季风也在不断加强[34]。中国特有种草地短柄草也是在此时间段分化形成。
祖先分布区重建结果也表明,小麦族与雀麦族祖先同属于北方干旱地区,而短柄草族起源于湿润的西南地区。现如今,短柄草族的分布也主要集中在湿润多雨区域,如喜马拉雅山脉南侧是海洋性气候,分布较多;而山脉北侧是大陆性气候,该族物种分布较少。短柄草族植物的共同祖先是古地中海分布的,随着古地中海的收缩退却和喜马拉雅山脉的隆起,中亚和西亚逐渐干旱[35],部分短柄草属植物退居在我国西南地区,西南地区的横断山脉在第四纪冰川时期也是多种生物的避难所。可以推测在古地中海收缩后,部分短柄草属植物迁移到我国西南地区,在第四纪冰川时期,气候的反复变化和青藏高原的隆起影响下形成现有分布格局。通过野外采集调查和网上资料收集,发现短柄草属植物的分布和我国山脉水系走向密切相关,该属植物主要分布在南北走向的横断山系和长江流域,东西走向的秦岭和华南地区南北走向的山脉限制了该属植物的扩张。
3.3 草地短柄草和短柄草进化关系
草地短柄草是我国特有种,但是与短柄草的关系一直存有疑问。在FOC被收录,但外籍作者认为可能是短柄草的局部变体,The plantlist和GBIF皆未收录。本研究的结果表明3个不同地区的短柄草聚成一支,3个不同地区的草地短柄草聚成一支,支持草地短柄草这一物种的成立。通过查看对比中科院北京植物研究所标本馆和中科院西北高原生物研究所标本馆以及iDigBio[36]公布的英国邱园、法国国家植物标本馆、加利福尼亚科学院标本馆、柏林达勒姆标本馆、美国国家标本馆、犹他州立大学植物标本馆、密歇根大学标本馆、密苏里植物园标本馆、北欧植物标本馆、奥斯陆大学自然历史博物馆、亚利桑那州立大学维管植物标本馆等标本馆的实物标本和高清图片,发现草地短柄草的具有小穗穗柄柄长大于2 mm的特点是该属其他物种所不具有的。
短柄草属植物全球广布,其属下15~22种,起源于33 Ma左右,但在各国植物志和地方志的记载总和远超此数字,其属下物种数目是亟待修订的。短柄草属植物起源于古地中海区域,但是具体的起源时间和地点有待深入考证,其在全球的分布成因也待进一步研究。短柄草属植物在我国西南地区广泛分布,也分布在澜沧江—怒江(湄公河—萨尔温江分水岭Mekong-Salween Divide)分水岭的两侧,其种群结构需要进一步的深入研究。
