外源ABA对低温不同光周期下四季秋海棠叶片花色素苷的诱导机理
2016-12-17郭美丽王佳婉靳慧慧张开明
郭美丽,王佳婉,王 珂,靳慧慧,张开明
(河南农业大学林学院,郑州 450002)
外源ABA对低温不同光周期下四季秋海棠叶片花色素苷的诱导机理
郭美丽,王佳婉,王 珂,靳慧慧,张开明*
(河南农业大学林学院,郑州 450002)
以‘超级奥林匹克’四季秋海棠(‘Super Olympia’Begoniasemperflorens)为材料,于常温(25 ℃/15 ℃)等日照条件下用0、5、10、50、100、500 μmol/L脱落酸(ABA)和低温(15 ℃/6 ℃)不同光周期下用10 μmol/L ABA分别喷施各处理植株,对不同处理下植株的色素含量、内源激素含量及其酶活性进行比较分析,探讨ABA对其叶片花色素苷合成的调控作用及其机理。结果显示:(1)常温等日照条件下,四季秋海棠叶片在5和10 μmol/L ABA处理后的第3~5天有明显变红趋势,且花色素苷含量和内源ABA含量显著增加,而内源赤霉素(GA)含量的下降幅度较为显著,相应的花色素苷合成关键酶和运输酶活性也显著提高。(2)低温条件下,四季秋海棠叶片花色素苷积累量与光周期密切相关,短日照处理的花色素苷积累量最大并显著高于等日照和长日照,但等日照与长日照下花色素苷积累量无显著差异;外源10 μmol/L ABA处理均可显著提高低温不同光周期下四季秋海棠叶片的花色素苷含量。(3)外施10 μmol/L ABA增加了低温下长日照和等日照处理中DFR(二氢黄酮醇-4-还原酶)还原反应中H供体NADPH的含量,促使DFR和UFGT(糖苷转移酶)的活性上调进而增加了花色素苷的含量;外施ABA处理均能够提高低温不同光周期处理组的内源ABA含量,降低内源GA的含量,与花色素苷的生成量相一致。研究表明,外源ABA能够通过调节花色素苷合成的关键酶来影响花色素苷的合成,外施适宜浓度ABA能够促进四季秋海棠叶片花色素苷的积累,可用于实际生产中的叶片着色调控管理。
四季秋海棠;脱落酸;花色素苷;低温;光周期
花色素苷(anthocyanin)是一种黄酮类次生代谢物,是一类从红到紫到蓝的植物色素,一般产生于植物叶片。当外界环境改变使得叶绿素遭到破坏,其含量明显下降时,色素也就随之而产生了[1]。影响花色素苷生成的环境因素有很多[2],其中光照、温度、旱涝、营养缺乏、激素、衰老等条件下都能影响花色素苷的形成。温度一般通过影响色素合成关键酶的活性,从而调节花色素苷的合成以及色素之间的组成和比例[3]。温度是影响叶片中花色素苷含量的重要因素。它不仅从基因转录水平上影响花色素苷合成,同时影响花色素苷的稳定[4]。低温可以诱导一些植物体内花色素苷合成的相关基因的表达,使花色素苷含量升高,如使PAL(苯丙氨酸解氨酶)、CHS(查尔酮合成酶)、CHI(查尔酮异构酶)和DFR(二氢黄酮醇-4-还原酶)基因的转录水平升高几倍;而高温却能抑制这些基因的表达,使花色素苷含量降低。光照也是影响彩叶植物叶色变化最重要的环境因素。它从强度、光质和光照时间等方面影响花色素苷的合成及调节相关酶的活性[5]。紫叶矮樱、紫叶小檗等必须在全光照下才能发挥其彩叶的最佳色彩[6]。彩叶秋海棠的叶色在遮阴处呈暗绿色,在全光下则呈红褐色[7]。光照是影响花青苷合成的最重要的外部因子,合成花青苷的4个关键酶,PAL、CHS、DFR、UFGT(糖苷转移酶)都是光调节酶,都以光敏色素为受体[8]。已有研究表明,外源ABA处理不仅可以提高果实的成熟度,促进果实软化和后熟衰老进程,而且在改善果实质量、促进果实着色方面也有明显的效果。
四季秋海棠(Begoniasemperflorens)是城市绿化当中应用较广的一种地被和花坛花卉,绿叶系列的‘超级奥林匹克’品种,能够在外界环境(如强光、低温等)的影响下积累花色素苷,而使叶片变红。低温对花色苷合成的促进是通过提高花色苷合成相关酶的活性, 也可通过强化结构基因的表达水平、 增加酶的含量来实现。脱落酸(abscisic acid, ABA)可提高一些植物的叶片或果实中 PAL、CHS、CHI、F3H(黄烷酮-3-羟基化酶)、UFGT 等酶的活性,促进花色苷合成[9]。如刘延吉等[10]发现ABA可提高南果梨(Pyrusussuriensis)PAL、CHI活性,促进果皮花色苷的积累。在低温条件下,不同光周期与外源ABA对花色素苷的积累是否有存在互作关系,目前我们并不清楚。因此,本实验中我们选用绿叶系列的‘超级奥林匹克’四季秋海棠品种作为材料,选用最适花色素苷积累的ABA激素浓度(10 μmol/L),研究外源激素ABA与不同光周期对花色素苷合成的影响。为花色素苷合成过程中的环境因子交叉作用的研究提供理论依据。
1 材料和方法
1.1 材料培养
参试材料‘超级奥林匹克’(‘Super Olympia’)绿叶系列的四季秋海棠品种,种子购于郑州贝利得花卉有限公司。将种子播种于25 cm宽、200个孔的穴盘内,营养土内含有3∶1∶1(v / v / v)泥炭、蛭石和珍珠岩混合而成的基质。生长环境为气温25 ℃/18 ℃(白天/夜晚)、光周期16 h /8 h(白天/夜晚)、相对湿度70%~85%,待植株长至3~4片叶时,挑选形态长势一致的幼苗分装成几组,以备实验处理使用。
1.2 材料处理
1.2.1 常温下不同浓度ABA处理 将分装的幼苗放于光照培养箱内,生长环境为25 ℃/15 ℃(白天/夜晚)、等日照、空气相对湿度70%~85%、光照强度800 lx。试验设置0、5、10、50、100、500 μmol/L ABA 6个浓度处理组,每组进行4次重复,每天于早上9:00和晚上9:00对其叶片正反面进行2次ABA喷施。分别在处理的第0、1、3和5天进行拍照,并取正常功能叶片测定相关生理指标。
1.2.2 低温不同光周期下外施ABA处理 根据以上实验的结论选用10 μmol/L的 ABA浓度,并在白天/夜晚15 ℃/6 ℃低温、光照强度为800 lx环境条件下进行不同光周期处理。试验处理时要把花蕾去除,选择生长基本一致的四季秋海棠绿叶品种植株。外源ABA处理设10 μmol/L ABA(ABA)和不喷施对照(CK)两个水平,光周期设置长日照(L,白天/夜晚16 h/8 h)、等日照(Q,白天/夜晚12 h/12 h)和短日照(S,白天/夜晚8 h/16 h)3个水平,共组成CK+L(对照+长日照)、ABA+L(ABA+长日照)、CK+Q(对照+等日照)、ABA+Q(ABA+等日照)、CK+S(对照+短日照)、ABA+S(ABA+短日照)6个处理组合。每个处理3次重复,每个重复选取5盆植株进行试验,处理时间为15 d。处理结束后取出拍照,并取足够的正常功能叶立即置于液氮中,保存在-80 ℃的冰箱内,用于测定相关生理指标。
1.3 测定指标及方法
1.3.1 花色素苷含量 花色素苷含量测定参照Mita等[11]的方法加以改进。取样0.1 g用液氮研磨,加1% HCl甲醇液放入2 mL离心管于4 ℃下保存24 h待测。13 000 g在 4 ℃离心15 min ,既可测定530 nm和650 nm处的吸光值A530和A650,按公式(A530- 0.25×A650)计算花色苷含量。
1.3.2 叶绿素含量 叶绿体色素含量参照Porra[12]的方法测定。各处理分别取0.2 g冻样用2 mL的80%丙酮在遮光下提取,分光光度计(SHIMADZUUV-2401PC)测量662 nm、644 nm、445 nm处的吸光值A662、A644、A445,通过以下公式计算出叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)的含量。
Chla=9.78×A662-0.99×A644
Chlb=21.4×A644-4.65×A662
Car=4.69×A445-0.268×(Chla+Chlb)
1.3.3 相关酶活性 (1)PAL和CHI活性 取0.3 g 冻样加入5 mL提取液(0.05 mol/L Na2HPO4/KH2PO4pH 7.0,0.05 mol/L抗坏血酸,0.018 mol/L巯基乙醇),冰浴匀浆,4 ℃下15 000 g离心20 min,上清液为酶粗提液用于测定PAL、CHI活性。3次取样重复。
PAL活性测定采用Lister[13]的方法。取0.5 mL粗酶液加入2.5 mL的反应液 (硼酸缓冲液pH 8.8,0.01 mol/L苯丙氨酸),置于34 ℃反应30 min,用三氯乙酸终止反应,在5 000 g离心5 min除去变性的蛋白质。反应生成的肉桂酸含量用在290 nm处的吸光值来表示。每分钟光密度变化0.001所需的酶量为1活性单位。对照实验用不加苯丙氨酸的反应液代替正常的反应液进行。
CHI活性测定采用Lister等[14]的方法进行。取0.75 mL的粗酶提液加入2 mL混合液(50 mmol/L Tris-HCl,pH 7.4,牛血清蛋白7.5 mg/mL,50 mmol/L KCN),然后加50 μL 1 mg/mL的查尔酮溶液置于34 ℃水浴30 min,另取同量的酶提液沸水浴10 min为对照,测定在381 nm吸光值的变化来表示CHI的活性。每分钟光密度变化0.001所需的酶量为1活性单位。
(2)DFR和UFGT活性 粗酶液提取参照Murray[14]报导的方法加以改进。取样品1 g加液氮研磨后,加入5 mL -20 ℃的丙酮混匀离心,弃去上清液,用4 mL -20 ℃丙酮再提取一次,沉淀用4 mL (0.1 mol/L硼酸缓冲液pH 8.8,5 mmol/L抗坏血酸)溶液提取,上清液为DFR和UFGT酶的粗提液。
DFR活性检测实验参照Stafford 和Lester[15]报导的方法。取0.5 mL酶提液加入0.6 mL反应混合液[(0.1 mol/L Tris缓冲液(pH 7.4),1 μmol/L脱氢栎精(Dihydroquercetin),1 mmol/L NADPH,1单位的葡萄糖-6-磷酸脱氢酶以及6 μmol/L的葡萄糖-6-磷酸)]共1.1 mL,在34 ℃水浴1 h后,用1 mL的乙酸乙酯提取3次,合并提液,然后用0.2 mL的蒸馏水提取3次。乙酸乙酯部分用氮气吹干,加入2.5 mL的正丁醇:盐酸(95∶5 V/V)于95 ℃水浴30 min,在550 nm 测定吸光值,用来表示由无色花色素(leuocyanidin)向花色素转变的量,取同量的酶提液沸水浴10 min为对照。每小时光密度变化0.001所需的酶量为1酶活性单位。
UFGT活性测定参照Lister和Lancaster[14]的方法。取酶液0.5 mL加入0.5 mL的反应液 [0.05 mol/L二甘氨酸缓冲液(pH 8.0),1 mmol/L的槲皮素和2.5 mmol/L的UDP-葡萄糖],反应液在34 ℃水浴30 min后,用0.75 mL 20% 三氯乙酸的甲醇溶液终止反应,于5 000 r/min离心5 min,上清液贮藏于-20 ℃待测。用高效液相色谱检测栎精的减少量并用其表示酶活性。
(3)谷胱甘肽-S转移酶(GST)活性 每个处理取0.3 g 待测样品加入1.8 mL的硼酸缓冲液, 离心后取上清液待测,利用南京建成生物工程研究所提供的谷胱甘肽-S转移酶测定试剂盒提供的方法进行测定。
1.3.4 NADP+和NADPH含量 称取约0.1 g组织样品,加入0.9 mL酸性提取液,冰浴研磨,煮沸5 min,冰浴中冷却后,10 000 g 4 ℃离心10 min,取上清液至另一新的离心管中,加入等体积的碱性提取液使之中和,10 000 g 4 ℃离心10 min,取上清液冰上保存供测定NADP+含量。另外,称取约0.1 g组织样品,加入0.9 mL碱性提取液,冰浴研磨,煮沸5 min,冰浴中冷却后,10 000 g 4 ℃离心10 min,取上清液至另一新的离心管中,加入等体积的酸性提取液使之中和,10 000 g 4 ℃离心10 min,取上清液冰上保存供测定NADPH含量。NADP+和NADPH含量测定采用苏州科铭生物技术有限公司试剂盒,按照相关试剂盒说明书进行相关操作。
1.3.5 ABA和GA含量 取样之前,用自来水将叶片冲洗干净之后,称取0.3 g样品(用液氮速冻后保存在-80 ℃的低温冰箱中),加2 mL样品提取液(80%甲醇,内含1 mmol/L BHT),在冰浴下研磨成匀浆,转入10 mL试管,再用2 mL提取液分次将研钵冲洗干净,一并转入试管中,摇匀后放置在4 ℃冰箱中。4 ℃下提取4 h,1 000 g离心15 min(实验中离心机型号LDZ5-2,4 000 r/min), 取上清液。沉淀中加1 mL提取液,搅匀,置4 ℃下再提取1 h,离心,合并上清液并记录体积,残渣弃去。将上清液用氮气吹干,除去提取液中的甲醇,用样品稀释液定容每个处理取0.3 g 样品样进行ABA、GA的酶联免疫检测,利用中国农业大学提供的ELISA植物激素测定试剂盒提供的方法进行。
1.4 数据分析
用Statistic 5.0进行差异显著性分析,用Excel 2007计算平均值和标准差,用OriginPro 7.5软件制图。
2 结果与分析
2.1 常温等日照下不同浓度外源ABA对四季秋海棠叶片内源激素、色素含量及相关酶活性影响
在常温(白天/夜晚:25 ℃/15 ℃)等日照条件下,叶面喷施系列浓度的ABA后,四季秋海棠叶片花色素苷相关合成酶(PAL、CHI、DFR、UFGT)和运输酶(GST)活性以及花色素苷、光合色素和内源激素含量发生了不同程度的变化(表1、2)。首先,外源喷施ABA对四季秋海棠叶片花色素苷的诱导作用因浓度不同而异,即在低浓度下(5~10 μmol/L),随着浓度的增加,外源ABA对花色素苷的诱导作用逐渐增强,叶片花色素苷含量分别比对照显著增加39.32% 和55.06%(P<0.05),且在10 μmol/L ABA处理5 d之后,四季秋海棠的叶片表现出肉眼可见的红色;而当喷施的ABA浓度较高(50、100和500 μmol/L)时,这种对花色素苷的诱导作用则消失,叶片花色素苷含量比对照不同程度降低。其次,花色素苷合成相关的PAL、CHI、DFR、UFGT和运输酶GST活性的变化与花色素苷的含量变化特征相符:在外源喷施ABA 浓度较低(5、10 μmol/L)时,5种酶活性较对照显著增加,且与施用ABA的浓度呈正相关;而当喷施的ABA浓度较高(50、100和500 μmol/L)时,5种酶活性与对照相比大多显著降低,部分没有显著变化。第三,外源喷施不同浓度的ABA之后,对植物的内源激素含量也造成不同的影响,即显著提高了内源ABA的含量,而显著降低了内源激素GA的含量,特别是10 μmol/L外源ABA处理下,内源ABA显著增加了60.93%,而内源GA却显著降低了38.35%。再次,与叶片花色素苷含量变化相对应,叶片叶绿素含量(a和b)和类胡萝卜素含量与外源施用ABA的浓度呈现显著的负相关,即随着ABA 浓度的增加而逐渐降低,如500 μmol/L的ABA 施用5 d后,叶绿素a、b含量分别比对照显著降低32.56%和29.73%,类胡萝卜素则比对照显著降低8.00%,说明喷施ABA能促使叶片中叶绿素降解。因此,我们在后续试验中选取10 μmol/L的ABA为试验浓度。
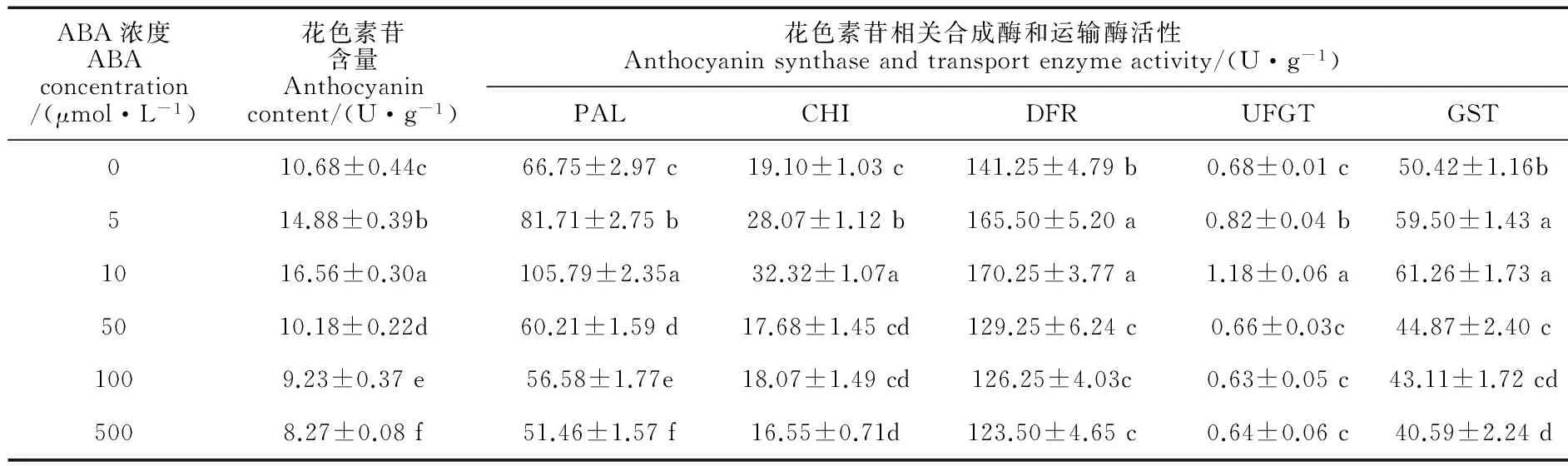
表1 常温等日照下外源ABA处理对四季秋海棠叶片花色素苷含量及相关酶活性的影响
注:同列不同小写字母表示处理间差异达到0.05显著水平;下同
Note: Different letters in the same column indicate significant differences among treatments at 0.05 level; the same as below
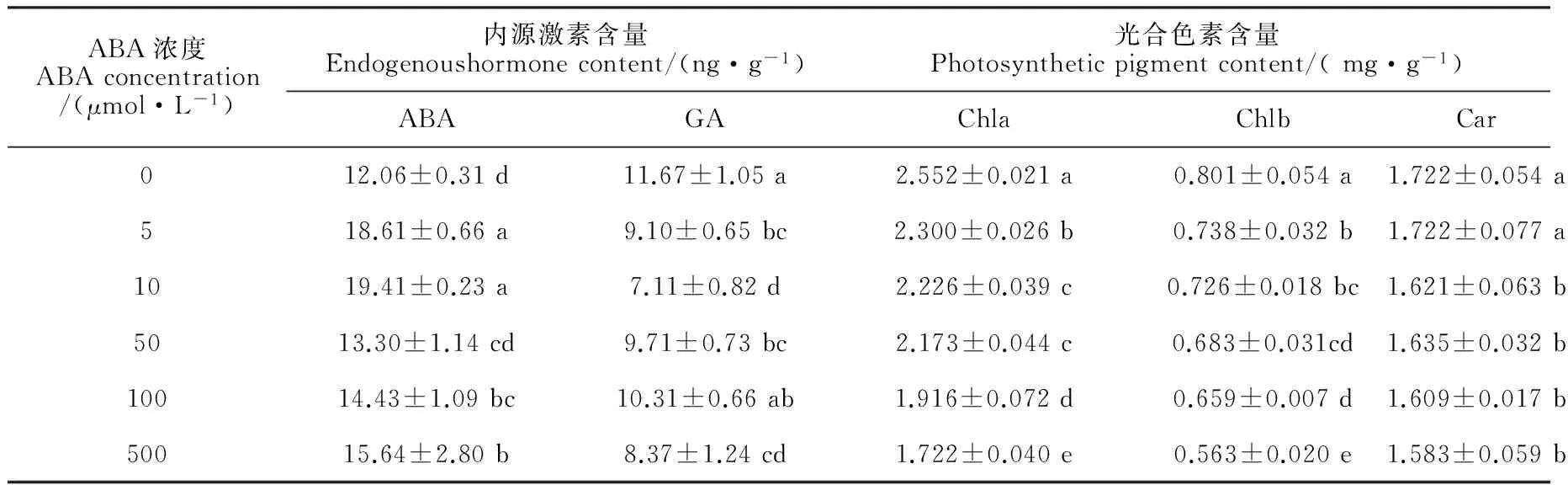
表2 常温等日照下外源ABA处理对四季秋海棠叶片内源激素及色素苷含量的影响
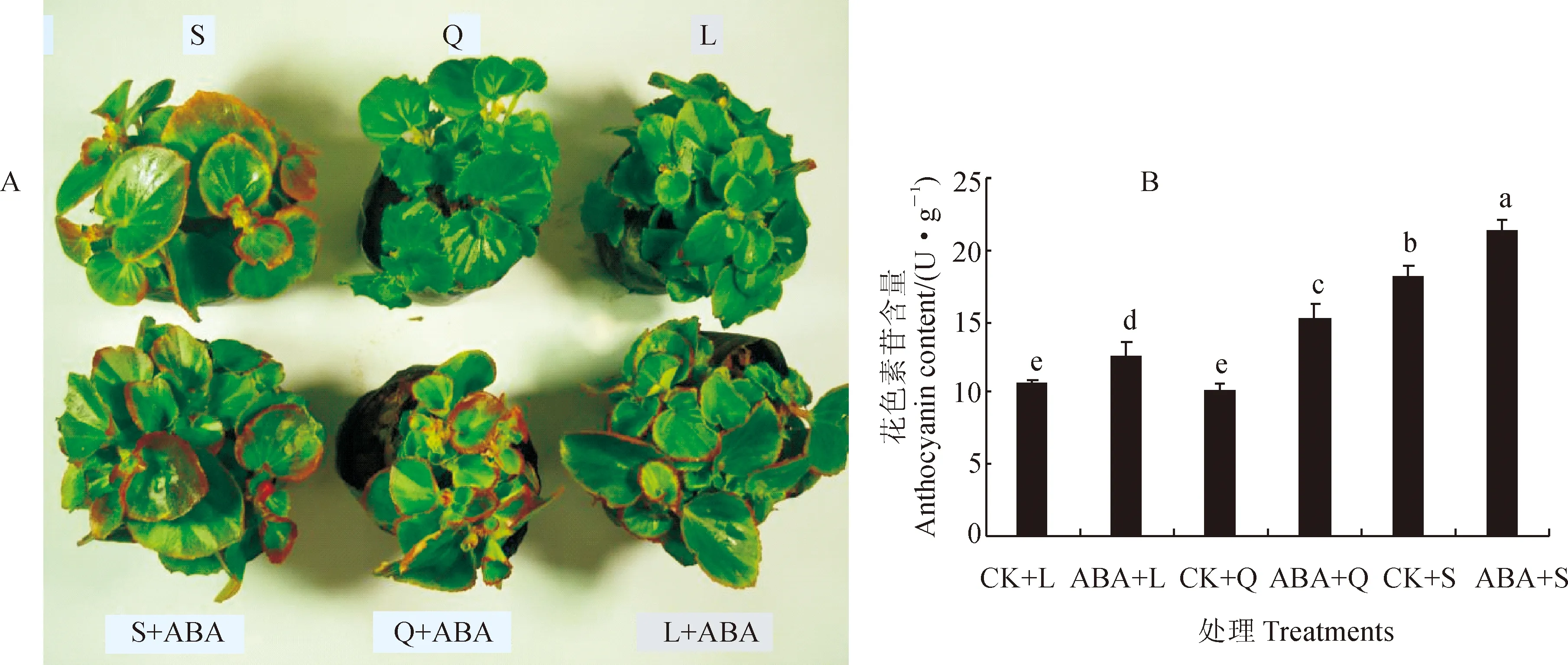
CK.不喷施ABA;L. 长日照(16 h/8 h);Q. 等日照(12 h/12 h);S. 短日照(8 h/16 h)。下同图1 低温不同光周期下外施10 μmol/L ABA对四季秋海棠花色素苷含量的影响CK.Control,without exogenous ABA;L. Long-day(16 h/8 h);Q. Intermediate-day(12 h/12 h);S. Short-day(8 h/16 h).The same as belowFig.1 Effect of 10 μmol/L exogenous ABA on anthocyamin content in leaves of B. semperflorens under low temperature and different photoperiodic treatment
2.2 低温不同光周期下外源ABA对花色素苷含量及相关酶活性的影响
前期研究表明,四季秋海棠的绿叶品种‘超级奥林匹克’在秋季时叶片因积累花色素苷而变红[16],这不仅是低温的作用[17],也是短日照的作用,因为在不适宜光周期(长日照和等日照)下,即使给予低温也不能诱导花色素苷合成[18]。在低温条件下(白天/夜晚:15 ℃/6 ℃),短日照处理(S)促进了四季秋海棠叶片明显变红,而同期的长日照(L)和等日照(Q)处理并未使四季秋海棠叶片变红(图1,A);同时,短日照处理(CK+S)的花色素苷含量显著高于其余处理,分别是长日照(CK+L)和等日照(CK+Q)处理的1.73和1.82倍(图1,B)。在同样的低温下,同时外施10 μmol/L的ABA之后,不论是短日照(ABA+S)、等日照(ABA+Q)还是长日照(ABA+L),四季秋海棠叶片都呈现肉眼可见的红色,同时它们叶片花色素苷含量较相应的对照处理(单独低温处理)显著增加,但仍以短日照处理显著较高(图1,A、B)。相应地,四季秋海棠叶片花色素苷合成酶和运输酶的活性也在喷施ABA之后较对照(单独低温处理)显著增加,尤其是DFR和UFGT活性在长日照条件下喷施ABA之后分别提高了5.20%、7.57%,在短日照条件下分别增加了6.41%、18.71%,在等日照条件下分别提高了10.60%和19.20%;另外,各日照处理间相比较,仍以短日照处理叶片各相关酶活性显著较高(表3)。这说明ABA在低温和不适宜光周期条件下,直接促进花色素苷合成下游的DFR和UFGT酶的增加,进而促进了花色素苷的合成和积累,从而验证了关于“低温短日照条件下诱导四季秋海棠叶片合成花色素苷的关键物质基础可能是ABA积累的推测。

表3 低温不同光周期下外施10 μmol/L ABA对四季秋海棠花色素苷相关合成酶和转移酶活性的影响
2.3 低温不同光周期下外源ABA对四季秋海棠叶片内源ABA和GA含量的影响
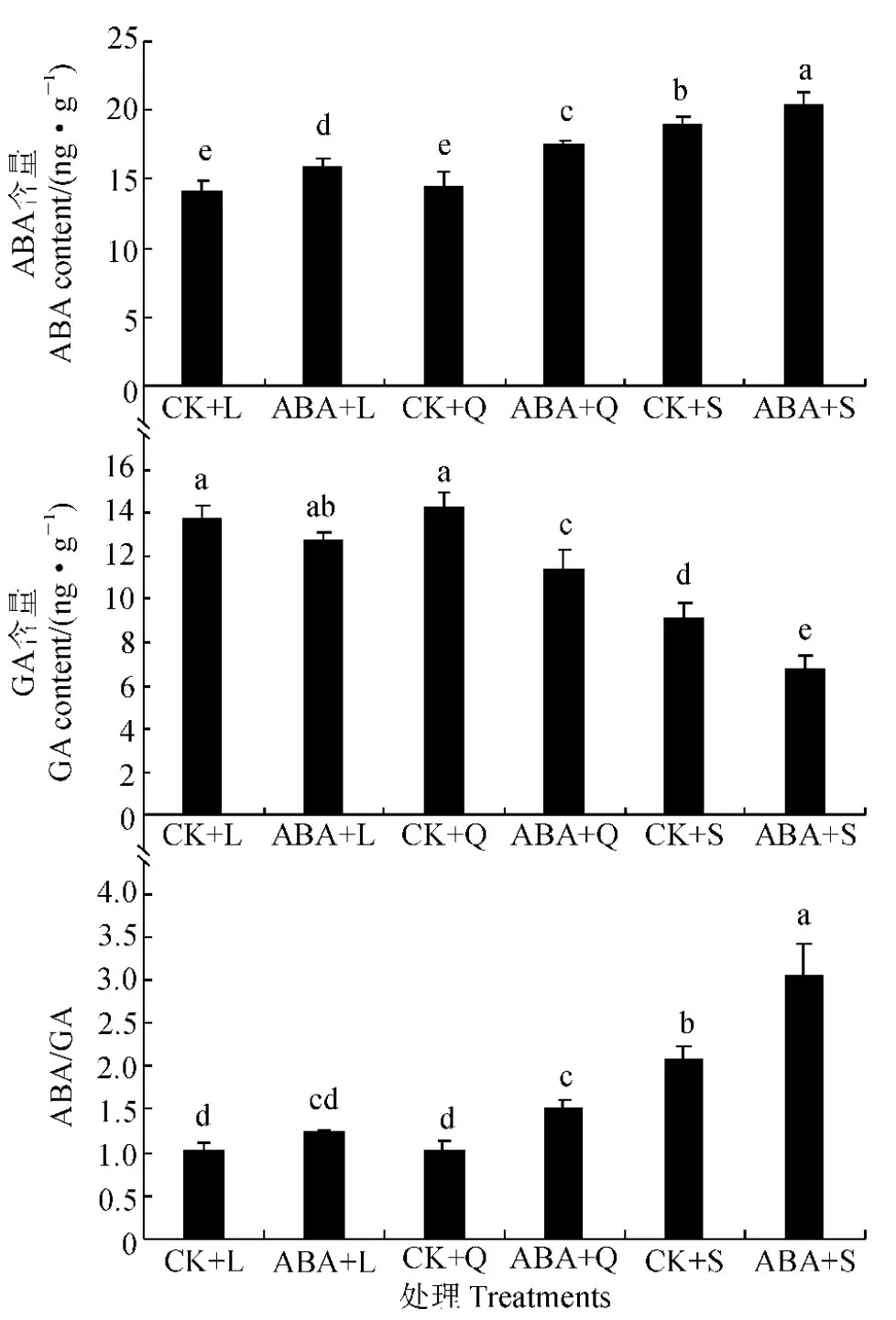
图2 低温不同光周期下外施10 μmol/L ABA后四季秋海棠ABA和GA含量Fig.2 ABA and GA contents under low temperature and different photoperiodic treatments with and without 10 μmol/L ABA on leaves of B. semperflorens
外源喷施ABA增加四季秋海棠叶片的内源ABA含量,而抑制内源GA含量,从而提高ABA/GA的比值(图2)。其中,在单独低温条件下,短日照处理四季秋海棠叶片的内源ABA含量比长、等日照处理显著较高,而内源GA含量泽显著较低,从而致使其ABA/GA的比值也显著较高;与单独低温处理对照组相比较,四季秋海棠叶片内源ABA含量在长、等、短日照处理分别显著增加12.35%、20.00%、10.59%,而内源GA含量却分别降低了7.61%、20.00%、25.00%,内源ABA/GA的比值则明显上升。因此,外源ABA调整了植物体内源激素的比例,使植物体内具有较高的内源ABA含量和较低GA的含量,内源ABA/GA的比值显著升高。

图3 低温不同光周期下外施10 μmol/L ABA对四季秋海棠叶片NADPH/NADP+比值变化Fig.3 NADPH/NADP+ ratio differences under low temperature and different photoperiodic treatments with and without 10 μmol/L ABA on leaves of B. semperflorens
2.4 低温不同光周期下外源ABA对四季秋海棠叶片NADPH和NADP+含量的影响
在花色素苷合成的途径中,DFR对花色素苷的最终形成起决定性作用,其反应需要NADPH参与[19]。由图3可知,外源ABA虽未显著提高低温和不适宜的光周期(长、等日照)下四季秋海棠叶片的NADPH+NADP+的总量,但却显著增加了NADPH的含量(分别比对照提高了6.58%和11.72%),因此NADPH/NADP+比值显著增加。这说明外源ABA处理后促进植物体内的NADP+向NADPH转变,因此为DFR催化二氢类黄酮醇生成无色花青素提供H+,从而启动了DFR的活性,进而促进花色素苷的合成。该结果验证了我们的推测:低温下不同光周期处理对四季秋海棠叶片花色素苷的诱导差异的物质基础是ABA,即低温短日照条件诱导四季秋海棠叶片合成花色素苷的关键物质基础是ABA的积累。
3 讨 论
通过考察外施ABA对四季秋海棠花色素苷合成的影响,发现不同浓度外源ABA处理下四季秋海棠花色素苷合成关键酶的变化趋势与花色素苷含量的动态趋势基本相吻合,较低浓度ABA(5~10 μmol/L)促进花色素苷合成酶(PAL、CHI、DFR和UFGT)及运输酶(GST)活力的提高,进而提高了花色素苷的含量,表明ABA是通过调节花色素苷合成的关键酶来调控四季秋海棠叶片花色素苷的合成。而较高浓度的外源ABA(50、100、500 μmol/L)则对这些酶的活力没有显著影响,因此没有花色素苷的生成。
在本试验中,低浓度的外源ABA通过启动花色素苷合成相关的酶(PAL、CHI、DFR和UFGT)和运输酶(GST),显著诱导了花色素苷在四季秋海棠叶片中的合成和积累。有研究表明,ABA的合成在果皮花色素苷的积累过程中起着关键的作用[20],900 mg/L的ABA处理能显著提高葡萄果实中的花色素苷含量[21];雷鸣等也发现1 000 mg/L的ABA对红地球葡萄果实的花色素苷具有明显的促进作用[22]。本试验研究表明低温、不同光周期条件下外源10 μmol/L ABA明显增加四季秋海棠叶片花色素苷的积累,这说明低温不同光周期条件下10 μmol/L ABA对四季秋海棠叶片花色素苷的积累有明显的促进作用,且花色素苷积累量与光周期密切相关,短日照时花色素苷积累量最大而长日照时积累量最小。
外施ABA处理能够调节植物体内ABA和GA的平衡,进而调控花色素苷的形成[23],本试验中外源ABA 对四季秋海棠叶片花色素苷的合成诱导可能是通过提高内源的ABA含量,并降低GA的含量来实现的。低温条件下,只有短日照启动了花色素苷合成及运输相关的酶活性,并诱导花色素苷显著积累,叶片显著变红,这可能与低温和短日照条件能显著诱导植物体内ABA含量增加有关。而在低温条件下,长日照和等日照处理植株体内的ABA 含量较低,而GA含量较高,因此花色素苷相关的合成酶和运输酶未被启动,因此没有花色素苷的合成。
花色素苷的生物合成过程中,要通过花色素苷合成关键酶(PAL、CHI、DFR 和 UFGT)相互协调,使花色素苷合成过程顺利进行。郭磊等[24]研究发现ABA参与了花色素苷合成的调控,对花色素苷合成具有促进作用。ABA还能通过转录因子调控结构基因的表达,进而提高酶的活性,最终促进花色素苷合成。本试验中,ABA处理后长日照和等日照下的四季秋海棠叶片花色素苷合成酶和运输酶活性显著提高,尤其是提高花色素苷合成途径中下游的DFR和UFGT活性,DFR在花色素苷合成的途径中发挥着关键作用,与花色的产生直接相关[25]。因此,喷施ABA能够诱导低温长日照和等日照处理下的四季秋海棠叶片合成花色素苷,关键因素可能在于对DFR活性的诱导。DFR是将二氢黄酮醇转化为花色素反应的第一个酶,它以不同二氢类黄酮醇为底物,利用辅因子NADPH中的H将4位的羰基还原为羟基[18],以转化生成相对应的不稳定的无色花青苷元,然后这些无色花青苷元在ANS,3GT催化下分别形成花青素、花葵素、花翠素。因此DFR活性的启动与否与NADPH的含量有直接关系,NADPH含量及NADPH/NADP+比值的增加为DFR提供足够的H供体,因此能启动DFR活性。本试验中,低温短日照处理下叶片NADPH的含量较长日照和等日照处理的高,NADPH/NADP+的比例也较高,因此DFR活性被启动,相应地,其下游的UFGT活性也显著增加,花色素苷最终合成;而低温长日照或等日照处理的植株,在外源喷施ABA之后,NADPH含量和NADPH/NADP+的比例显著增加,因此DFR和UFGT被启动,诱导花色素苷合成。
本试验研究发现,低温不同光周期条件下外施10 μmol/L ABA 增加了 四季秋海棠叶片NADPH/NADP+比值,这说明外施 ABA 增加了 DFR 还原反应的 H 供体的NADPH 含量,这可能是提高 DFR活性的一个重要因素。因此,DFR对花色素苷的最终形成起决定性作用。
综上所述,不同浓度ABA对四季秋海棠花色素苷合成关键酶的变化趋势与花色素苷含量的动态趋势相吻合,较低浓度ABA促进花色素苷合成酶PAL、CHI、DFR和UFGT活力的提高,进而提高了花色素苷的含量,而较高的浓度则抑制了PAL、CHI、DFR和UFGT酶活力,抑制了花色素苷的生成,这说明ABA通过调节花色素苷合成的关键酶来影响花色素苷的合成。外源ABA在低温不同光周期条件下能使花色素苷含量、相关酶活性、ABA/GA提高,但对叶绿素含量和GA含量却有抑制作用,尤其在低温短日照条件效果更显著。因此,ABA处理可作为一种有效的方法,用于叶片着色机制的研究。
[1] 丁 锐.国外花色素苷的研究现状与进展[J].汉中师范学院学报(自然科学),2004,22(2):73-78.
DING R. Advances on the research of anthocyanins at abroad [J].JournalofHanzhongTeachersCollege(Natural Science), 2004, 22(2):73-78.
[2] DOONER H K. What is anthocyanin[J].Mol.Gen.Genet, 1983,198:136-141.
[3] 于晓南,张启翔.观赏植物的花色素苷与花色[J].林业科学,2002,38(3):147-153.
YU X N, ZHANG Q X. Anthocyanin in ornamental plant and color ecpress[J].ScientiaSilvaeSinicae, 2002,38(3):147-153.
[4] 张学英,张上隆,骆 军,等.果实花色素苷合成研究进展[J].果树学报,2004,21(5):456-460.
ZHANG X Y, ZHANG S L, LUO J,etal. Advances in research on fruit anthocyanin synthesis [J].JournalofFruitScience, 2004,21(5):456-460.
[5] 李红秋,刘石军.光强度和光照时间对色叶树叶色变化的影响[J].植物研究,1998,18 (2):194-205.
LI H Q, LIU S J. Effects of light intensity and illumination time on leaf colour variations of coloured leaf trees [J].BulletinofBotanicalResearch,1998,18(2):194-205.
[6] 张启翔,吴 静,周肖红.彩叶植物资源及其在园林中的应用[J].北京林业大学学报,1998,18(2):194-205.
ZHANG Q X, WU J, ZHOU X H. Resource of color leaf plants and its application in Gardens [J].JournalofBeijingForestryUniversity,1998,18(2):194-205.
[7] SMITH L B, WASSHAUSEN D C. Begonia solimutata a new Brazilianspeciesw hose leaf color varies with light intensity [J].Begonian, 1990, 57: 151-152.
[8] JUZ G, LIU C L, YUAN Y B. Activity of chalcone synthesis and UGDP: falvonoid-3-glycosy transferaseinrelation to anthocyanin synthesis in apple[J].ScientiaHortieuhurae, 1995, 63: 175-l 85.
[9] 陈文龙,赵昶灵,支伟特,等.高等植物花色苷生物合成调控的研究进展[J].中国农学通报, 2013,29(33): 271-276.
CHEN W L, ZHAO C L, ZHI W T,etal. Progress of regulation of anthocyanin biosynthesis in higher plants[J].ChineseAgriculturalScienceBulletin,2013,29(33):271-276.
[10] 刘延吉,王 嵩,吕德国.ABA对南果梨花色素苷含量、合成酶及信号分子的影响[J].沈阳农业大学学报,2009,40(1):7-10.
LIU Y J, WANG S, LÜ D G. Effect of ABA on anthocyanin, partial synthetic enzymes and signaling molecule Nanguo Pear [J].JournalofShenyangAgriculturalUinversity,2009,40(1):7-10.
[11] MITA S, MURANO N, AKAIKE M,etal. Mutants of Arabidopsis thaliana with pleiotropic effects on the expression of the gene for β-amylase and on the accumulation of anthocyanin that are inducible by sugars [J].ThePlantJournal, 1997, 11(11): 841-851.
[12] PORRA R J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophyll a and b[M]. Discoveries in Photosynthesis. Springer Netherlands, 2005: 633-640.
[13] MURRAY J R, HACKETT W P. Dihydroflavonol reducatase activity in relation to differential anthocyanin accumulation in juvenile and mature phaseHederahelixL.[J].PlantPhysiology, 1991, 97: 343-351.
[14] LISTER C E, LANCASTER J E, WALKER J R L. Developmental Changes in Enzymes Biosynthesis in the Skins of Red and green apple cultivars[J].JournalofScienceFoodandAgriculture, 1996, 71: 313-320.
[15] STAFFORD H A, LESTER H H. Flavan-3-ol biosynthesis the conversion of (+)-dihydroquercetin and flavan-3, 4-cis-diol (leucocyanidin) to (+)-catechin by reductases extracted from cell suspension cultures of douglas fir [J].PlantPhysiology, 1984, 76(1): 184-186.
[16] ZHANG K M, LI Z, LI Y,etal. Carbohydrate accumulation may be the proximate trigger of anthocyanin biosynthesis under autumn conditions inBegoniasemperflorens[J].PlantBiology,volume,2013, 15: 991-1 000.
[17] ZHANG K M, GUO M L, HE D,etal. The inhibition effect and excessive carbon flux resulting from blocking anthocyanin biosynthesis under darkness inBegoniasemperflorens[J].PlantGrowthRegul., 2016, 35: 22-30.
[18] 王 珂.外部因素对四季秋海棠花色素苷合成的影响[D].郑州:河南农业大学,2014.
[19] 张学英,张上隆,骆 军,等.果实花色素苷合成研究进展[J].果树学报,2004,21(5):456-460.
ZHANG X Y, ZHANG S L, LUO J,etal. Advances in research on fruit anthocyanin synthesis [J].JournalofFruitScience,2004,21(5):456-460.
[20] 李 明.ABA、S3007促进苹果果实着色的研究[D].沈阳:沈阳农业大学,2005:20-25.
[21] 胡春霞,王 丽.ABA、BR对葡萄果实花色素苷和可溶性糖的影响[J].鞍山师范学院学报,2009,11(2):42-44.
HU C X, WANG L. Effect of ABA and BR on anthocyanin and soluble sugar of grape seeds [J].JournalofAnshanNormalUniversity, 2009, 11(2): 42-44.
[22] 雷 鸣,吴 江,等. ABA与NAA对红地球葡萄果实形状的影响[J].浙江农业科学,2008,2:153-155.
LEI M, WU J,etal. Effect of ABA and NAA of Hongdiqiu grape fruit shape [J].JournalofZhejiangAgriculturalSciences,2008, 2:153-155.
[23] 刘延吉,向文娟,田晓艳.南果梨愈伤组织及花青素诱导的初步研究[J].食品与生物技术学报,2008,27(3):83-87.
LIU Y J, XIANG W J, TIAN X Y. Study on inducing callus and cyaniding of Nanguo pear [J].JournalofFoodScienceandBiotechnology,2008,27(3):83-87.
[24] 郭 磊,蔡志翔,张斌斌,等.叶片喷施脱落酸对桃果实着色及相关基因表达的影响[J].植物营养与肥料学报,2013,19(6):1 463-1 469.
GUO L, CAI Z X, ZHANG B B,etal. Effect of foliar application of ABA on fruit coloration and expression of some related genes in peach [J].JournalofPlantNutritionandFertilizer, 2013,19(6):1 463-1 469.
[25] 周 琳,王 雁,任 磊,等.牡丹二氢黄酮醇4一还原酶基因PsDFR1的克隆及表达分析[J].植物生理学报,2011,47(9):885-892.
ZHOU L, WANG Y, REN L,etal. Cloning and expression analysis of dihydroflavonol 4-reductase genePsDFR1 from tree peony (PaeoniasuffruticosaAndr)[J].PlantPhysio-logyJournal, 2011,47(9):885-892.
(编辑:裴阿卫)
Mechanism of ABA-induced Anthocyanin Biosynthesis in Begoniasemperflorens under Different Photoperiods and Low Temperatures
GUO Meili, WANG Jiawan, WANG Ke, JIN Huihui, ZHANG Kaiming*
(Forestry College, Henan Agricultural University, Zhengzhou 450000, China)
Taking the ‘Super Olympic’BegoniaSemperflorensas material, at room temperature (25 ℃/15 ℃) under normal sunshine condition or low temperature (15 ℃/6 ℃) under the different photoperiods, we sprayed the plants with abscisic acid (ABA). The endogenous pigment content, hormone contents and enzyme activities were comparatively analyzed to discuss the effects of exogenous ABA on anthocyanin biosynthesis and its mechanism. The results showed: (1) at room temperature and normal sunshine condition, the leaves will get red obviously on the 3th days when applying ABA (5 and 10 μmol/L). The anthocyanin content and endogenous ABA content of pigment increased significantly, while the endogenous gibberellin (GA) content decreased significantly. The anthocyanin biosynthesis and trans portation also increased significantly. (2) Under the condition of low temperature, the anthocyanin accumulation was closely related to photoperiod and the anthocyanin accumulation was the largest in short day; The anthocyanin accumulation in sunshine condition also increased, but there was no significant difference between sunshine and long sunshine. (3) The addition of 10 μmol/L ABA treatment increased the content of H donor NADPH in DFR reduction under long day and sunshine condition. Under low temperature treatment, the activities of DFR and UFGT enzymes increased and then increased the anthocyanin content. In addition, exogenous ABA treatment could increase endogenous ABA content and decrease the content of endogenous GA in low temperature and different photoperiod treatments, which was consistent with the production of anthocyanin. The results showed that exogenous ABA can affect the synthesis of anthocyanins by regulating key enzyme in anthocyanin synthesis of flowers. The application of suitable concentration of ABA can promote theB.semperflorensleaf anthocyanin accumulation and can be used to control leaf coloring in actual production.
Begoniasemperflorens; abscisic acid; anthocyanin; low temperature;photoperiod
1000-4025(2016)10-1999-09
10.7606/j.issn.1000-4025.2016.10.1999
2016-05-19;修改稿收到日期:2016-10-15
国家自然科学基金(311101562)
郭美丽(1989-),女,硕士,主要从事园林植物生长发育与调控。E-mail:1278658212@qq.com
*通信作者:张开明,副教授,硕士生导师,主要园林植物生长发育与调控。E-mail:miss1199@126.com
Q945.18; Q945.78
A
