猪链球菌3-羟基-3-甲基戊二酰辅酶A还原酶的原核表达、活性检测和同源建模
2016-12-13刘璨颖徐卓菲
刘璨颖,徐卓菲,付 强
(1.佛山科学技术学院生命科学与工程技术学院,佛山 528000;2.华中农业大学动物科学与动物医学院,武汉 430070)
猪链球菌3-羟基-3-甲基戊二酰辅酶A还原酶的原核表达、活性检测和同源建模
刘璨颖1,徐卓菲2,付 强1
(1.佛山科学技术学院生命科学与工程技术学院,佛山 528000;2.华中农业大学动物科学与动物医学院,武汉 430070)
旨在以猪链球菌(Streptococcussuis,S.suis)3-羟基-3-甲基戊二酰辅酶A还原酶(HMGR)为药靶,给S.suis病防治研制新型抗菌药提供理论和试验基础。原核表达了S.suisHMGR,用Ni-NTA树脂对重组蛋白质进行了纯化,并用分光光度法测定了其酶活力。此外,运用CLUSTALW对同源HMGR氨基酸序列进行多序列比对。通过SWISS-MODEL对S.suisHMGR三维结构进行同源建模,并用PROCHECK评价模型的质量。酶活力测定结果显示,催化HMG-CoA还原为甲羟戊酸的反应中,S.suisHMGR最佳反应温度和pH分别为37 ℃和5.0,其Vmax和Km为846 U·mg-1和0.213 mmol·L-1。经分析显示,肺炎链球菌HMGR晶体结构(PDB ID:3QAE_A)是构建S.suisHMGR三维结构的最佳模板。构建的三维结构模型经质量评估证实为可靠的理论模型,将有助于虚拟筛选出对抗S.suis感染的新型抗生素,酶活力的测定为用试验手段验证S.suisHMGR抑制剂的有效性提供了可行性的方法。
链球菌;HMGR;原核表达;蛋白质纯化;酶活测定;同源建模
猪链球菌(Streptococcussuis,S.suis)是一种重要的人畜共患病原菌,能引起宿主广泛的临床疾病。猪感染S.suis后的临床症状包括脑膜炎、肺炎、败血症、心内膜炎、多发性浆膜炎、关节炎、流产和脓肿等[1-6]。S.suis甚至可以引起接触过感染猪或消费生猪产品的成人全身性感染[7]。由于针对S.suis感染没有合适的免疫接种措施,并且S.suis的耐药性日益增加,通过疫苗和抗生素治疗不能有效控制S.suis感染[8]。因此,急需从S.suis代谢通路中寻找新的靶标并研制新型抗生素。
细菌中异戊二烯类分子具有许多重要的生物功能,其中包括涉及电子传递,作为细胞壁合成中的载体和参与产生抗生素形成竞争优势等。异戊二烯类分子由五碳前体异戊二烯焦磷酸(isopentenyl diphosphate, IPP)或其同分异构体二甲基丙烯基磷酸衍生而来。合成IPP的途径有两种:甲羟戊酸途径和非甲羟戊酸途径[9-13]。甲羟戊酸途径中3-羟基-3-甲基戊二酰辅酶A还原酶(3-hydroxy-3-methylglutaryl coenzyme A reductase, HMGR)催化HMG-CoA生成甲羟戊酸,是该途径中关键的限速酶。D. A. Bochar等基于氨基酸序列分析认为有两种不同类HMGR。编码I类HMGR的基因存在于所有的真核生物,许多古生菌和一些链霉菌属中。而编码II类HMGR的基因存在于一些真细菌和闪烁古生球菌中[14-16]。I类和II类HMGR有共同的催化机制,但是其三维结构和对他汀类抑制作用的敏感性有显著的差异[17-20]。在细菌中,W. Li等以肺炎链球菌II类HMGR为药靶,用基于结构的虚拟筛选和试验验证,得到3个肺炎链球菌新型抗生素[21]。此外,E. I. Wilding等通过基因干扰试验证实编码II类HMGR的基因mvaA对金黄色葡萄球菌体外生长是必须的[14]。因此,细菌II类HMGR可以成为针对多耐药性病原菌抗生素靶标。
本研究成功原核表达、纯化了猪链球菌HMGR,并测定其酶促动力学参数,此外,运用同源建模设计了猪链球菌HMGR理论三维结构模型,为今后虚拟筛选出对抗S.suis感染的HMGR抑制剂和运用试验验证抑制剂的有效性奠定了理论和试验基础。
1 材料与方法
1.1 菌株
猪链球菌SS2 SC-19菌株2005年分离于中国四川省流行病暴发时的一头病猪[21]。感受态大肠杆菌DH5α、BL21购自宝生物工程(大连)有限公司;原核表达载体pET-28a(含有His标签)购自Novagen公司。S.suis培养在添加了10%新生牛血清的TSB培养基中。大肠杆菌培养在LB或LA培养基中。抗性筛选时,大肠杆菌培养在含有50 μg·mL-1卡那霉素的培养基中。
1.2 主要试剂
限制性内切酶BamHⅠ和XhoⅠ,TaqDNA聚合酶,T4 DNA连接酶,异丙基硫代半乳糖苷(IPTG),DNA marker DL2000购自宝生物工程(大连)有限公司;蛋白质预染marker购自加拿大Fermentas公司;HMG-CoA购自sigma公司;DTT,新生牛血清购自四季青公司;NaCl,蛋白胨和酵母提取物购自OXXID公司;琼脂粉购自武汉洁洋盛生物科技有限公司;TSB购自BD公司;DNA(胶)回收试剂盒购自上海捷瑞生物工程有限公司;细菌DNA提取试剂盒,Ni-NTA树脂购自Qiagen公司。
1.3 引物设计与基因的扩增
根据GenBank中登录的S.suisBM407的mvaA基因序列(登录号:YP_003029230.1)设计引物,上游引物:(5′-ATCGGGATCCATGTCAAC TTTTTCCGGATT-3′)和下游引物:(5′-GGCG CTCGAGGCCATCATGGATGTCTTTC-3′),下划线部分分别为BamHⅠ和XhoⅠ酶切位点,引物在上海生工生物技术有限公司合成。用细菌DNA提取试剂盒提取猪链球菌SS2 SC-19菌株基因组DNA,以其为模板,PCR扩增mvaA基因,反应条件为94 ℃预变性5 min;94 ℃ 30 s, 54 ℃ 30 s, 72 ℃ 1 min共35个循环;最后72 ℃再延伸7 min。
1.4 重组原核表达质粒的构建
用BamHⅠ和XhoⅠ分别双酶切原核表达载体pET-28a和回收的mvaA片段,酶切产物经胶回收试剂盒纯化后,用T4 DNA Ligase 16 ℃连接过夜,将连接产物转化感受态大肠杆菌DH5α,提取重组质粒,经PCR和酶切鉴定。
1.5 重组HMGR蛋白的表达和纯化
将鉴定正确的重组质粒送至上海生工生物有限公司测序后,利用测序结果在线NCBI Blastn比对GenBank数据库(http://blast.ncbi.nlm.nih.gov/Blast.cgi),检测序列的同源基因。取含有重组质粒的BL21菌液接种到含有Kanamyci终浓度为50 μg·mL-1的LB液体培养基,180 r·min-137 ℃培养过夜。次日以1∶100体积比例接种到含有Kanamyci终浓度为50 μg·mL-1的LB液体培养基,200 r·min-137 ℃培养2.5 h,菌液OD值达到0.5~0.8时,加入IPTG至终浓度为0.8 mmol·L-1诱导HMGR蛋白表达。在180 r·min-137 ℃培养5 h后,菌液于4 ℃ 10 000 r·min-1离心2 min。沉淀用4 ℃裂解液(含有50 mmol·L-1磷酸钠,0.3 mol·L-1NaCl,5 mmol·L-1DTT,10 mmol·L-1咪唑,pH 8.0)重悬并高压破碎,收集上清液并用SDS-PAGE检测目的蛋白质的表达。用1 mLNi-NTA琼脂糖柱纯化HMGR重组蛋白。高压破碎后收集的上清液加入Ni-NTA琼脂糖柱,用4 ℃裂解液平衡。用5倍柱体积的洗涤液(含有50 mmol·L-1磷酸钠,0.3 mol·L-1NaCl,5 mmol·L-1DTT,20 mmol·L-1咪唑,pH 8.0)洗柱。用8 mL洗脱液(含有50 mmol·L-1磷酸钠,0.3 mol·L-1NaCl,5 mmol·L-1DTT,250 mmol·L-1咪唑,pH 8.0)洗脱Ni-NTA琼脂糖柱上结合的重组HMGR蛋白。用生物分光光度计(Eppendorf)测量蛋白质浓度。
1.6 HMGR活性测定
HMGR活性测定参照李鹏等报道的方法并加以改进[22]。用BioTek分光光度计(Synergy, Gene Company limited)测定NAD(P)H在340 nm处吸光值的减少来检测分析HMGR酶活力。标准的酶反应体系(总体积为200 μL)包括0.25 mmol·L-1NADPH, 0.25 mmol·L-1HMG-CoA, 50 mmol·L-1NaCl, 1 mmol·L-1EDTA, 5 mmol·L-1DTT, 25 mmol·L-1KH2PO4(pH 6.5)和纯化后的HMGR。反应体积中加入HMG-CoA后,反应开始。一个酶活力单位定义为每分钟消耗2 μmol NADPH所需要的HMGR酶量。以未加IPTG诱导的细胞破碎上清液经过同样Ni-NTA柱纯化处理后的液体作为阴性对照。
1.7 序列分析和同源建模
利用NCBI blastp (http://blast.ncbi.nlm.nih.gov/Blast.cgi) 对S.suisHMGR氨基酸序列进行序列比对分析和保守域预测。运用CLUSTALW对球菌和博氏螺旋体中HMGR氨基酸序列进行多序列比对,其中包括肺炎链球菌(GenBank登录号EDK78539.1),无乳链球菌(GenBank登录号AKI57711),酿脓链球菌(GenBank登录号AKL62895.1),猪链球菌(GenBank登录号YP_003029230.1),表皮葡萄球菌(GenBank登录号KTF26195.1),金黄色酿脓葡萄球菌(GenBank登录号WP_001180324.1),PseudomonasMevalonii(PDB登录号4I6A_B)和博氏螺旋体(GenBank登录号NP_212819.1)[23]。将S.suisHMGR氨基酸序列上传至SWISS-MODEL在线服务器(http://swissmodel.expasy.org)进行同源建模,获得该蛋白质的三级结构[24]。用PROCHECK评价所建模型的质量[25]。
2 结 果
2.1 序列分析和同源建模
经PCR和酶切鉴定为阳性的重组质粒送至公司测序,结果显示重组质粒中mvaA基因与已报道的S.suismvaA基因(YP_003029230.1)核苷酸序列相似性为100%。鉴定正确的表达载体pET-HMGR转化到E.coliBL21,用0.8 mmol·L-1IPTG于37 ℃诱导5 h。纯化后的表达产物经SDS-PAGE分析显示出一条相对分子质量约为46 ku的蛋白质条带(图1),与其理论值相符。可溶性粗提取物在经过Ni-NTA树脂纯化后的比活力为6.56 μmol·(min·mg)-1。
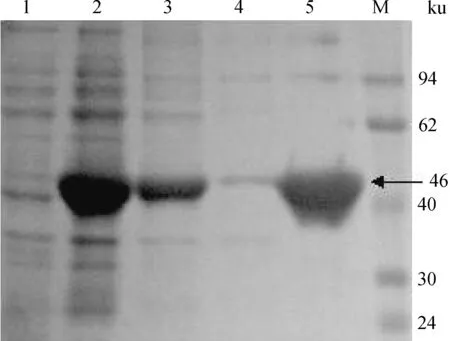
1. 未经IPTG诱导; 2. IPTG诱导; 3. 表达产物上清; 4. 表达产物沉淀; 5. 纯化后的重组HMGR蛋白; M. 蛋白质相对分子质量标准1. Not induced with IPTG; 2. IPTG induction; 3. Supernatant of expression product; 4. Precipitate of expression product; 5. The purified recombinant HMGR protein; M. Protein molecular weight marker图1 重组HMGR蛋白SDS-PAGEFig.1 SDS-PAGE of the recombinant HMGR
2.2 HMGR的动力学特征
重组HMGR的酶活力测定基于分光光度计,检测NADPH在340 nm处吸收值的变化。HMGR的最适反应pH为5.0,最适反应温度为37 ℃(图2)。在催化HMG-CoA还原为甲羟戊酸的反应中,HMGR的Vm和Km值分别为846 U·mg-1和0.213 mmol·L-1。
2.3 序列分析与同源建模
NCBI blastp分析显示,S.suisHMGR氨基酸序列5—417区段属于Class II HMGR (CDD ID:cd00644)。S.suisHMGR与肺炎链球菌、无乳链球菌、酿脓链球菌、表皮葡萄球菌、金黄色酿脓葡萄球菌、PseudomonasMevalonii和博氏螺旋体中HMGR氨基酸序列相似性分别为60%、60%、59%、43%、44%、42%和33%。将S.suisHMGR和球菌、螺旋体HMGR氨基酸序列进行多序列比对,结果如图3。有研究Pseudomonasmevalonii
HMGR晶体结构的报道揭示,其与底物HMG-CoA结合的保守序列为ENVIG(I),与底物NAD(H)结合的保守序列为DAMGXN(II)和GTVGG(III),这三段保守序列在图3红色方框中标示出来[18]。
SWISS MODEL分析显示肺炎链球菌HMGR的晶体结构(PDB-ID: 3QAE-1A)是S.suisHMGR建模的最佳模板。SWISS MODEL服务器对S.suisHMGR建模的结果用二维图像显示见图4。HMGR为对称的二聚体结构,SWISS MODEL显示的为整体结构。
用PROCHECK检测S.suisHMGR三维模型的合理性,结果见图5。图5中拉马钱德兰(Ramachandran)图统计表明在SWISS MODEL构建的模型中,分别有91.8%氨基酸残基的二面角落在最适宜区域(图中红色区域),7.2%的氨基酸残基的二面角落在其他的允许区域(图中黄色区域),0.6%的氨基酸残基的二面角落在勉强的允许区域(图中浅黄色区域)。而构建的模型A链和B链中164位的苯丙氨酸(PHE)和174位的谷氨酰胺(GLN)落在非允许区,但这两个氨基酸并不位于HMGR与底物结合的保守序列中(图3)。整体表明S.suisHMGR模型的立体构象符合化学二面角分布的要求。SWISS MODEL构建的模型G-factor参数为0.16,大于-0.5,说明模型中共价键及键角存在的构象合理。质量评价结果说明SWISS MODEL构建的S.suisHMGR三维结构模型合理可靠。
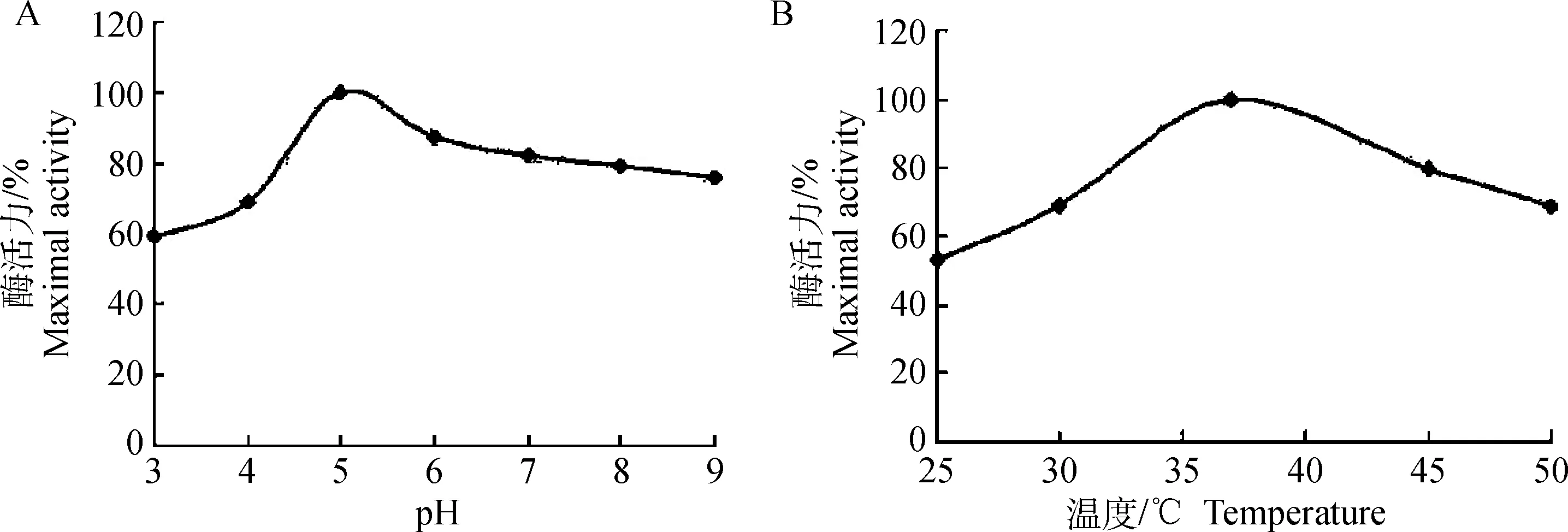
A. pH对酶活力的影响; B. 温度对酶活力的影响A. Effect of pH on activity; B. Effect of temperature on activity图2 pH和温度对S. suis HMGR酶活力的影响Fig.2 Effect of pH, temperature on HMGR activity

破折号表示引入序列的空隙以优化比对。数字表示氨基酸的位置。在所有的比对中,缩写如下:S. pne, 肺炎链球菌;S. aga, 无乳链球菌;S. pyo, 酿脓链球菌;S. suis, 猪链球菌;S. epi, 表皮葡萄球菌;S. aur, 金黄色酿脓葡萄球菌;P. Mev, Pseudomonas Mevalonii;B. bur, 博氏螺旋体Dashes indicate gaps introduced into the sequence to optimize the alignment. Numbers refer to amino acid positions. In all alignments, abbreviations are as follows: S. pne, Streptococcus pneumoniae; S. aga, Streptococcus agalactiae; S. pyo, Streptococcus pyogenes; S. suis, Streptococcus suis; S. epi, Staphylococcus epidermidis; S. aur, Staphylococcus aureus; P. Mev, Pseudomonas Mevalonii; B. bur, Borrelia burgdorferi 图3 球菌和螺旋体中HMGR多序列比对Fig.3 Multiple alignment of sequences of HMGR from coccus and Borrelia burgdorferi
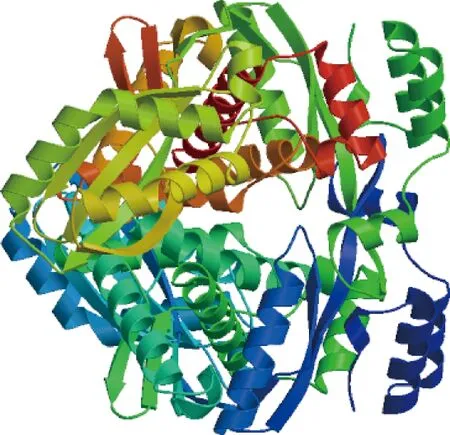
图4 S.suis HMGR同源建模二维显示Fig.4 Two-dimensional display of homology modeling of S. suis HMGR
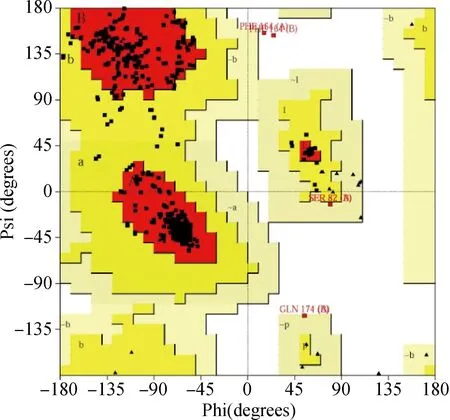
图5 S. suis HMGR模型的拉马钱德兰图Fig.5 Ramachandran plot of the 3D model of S. suis HMGR
3 讨 论
S.suis给养猪业带来了严重的经济损失,阻碍了畜牧业的发展,也影响着人们的健康安全。人感染S.suis后出现的临床症状包括脑膜炎、败血病、关节炎等。有研究统计,目前为止,全球有30多个国家报道过人感染S.suis,有大约1 600例严重的人感染S.suis事例,并且在过去十年里,人感染S.suis事例急剧增加。为了对抗S.suis感染,研究人员用试验证实至少有15个保护性抗原可以作为S.suis的候选疫苗。但是由于重组蛋白质表达方式、菌株、免疫程序、免疫动物的差异,导致同一保护性抗原的免疫保护力结果不同[6, 26]。而且,S.suis日益增高的耐药性使抗生素防控手段受限[8]。因此,需要寻找新的药物靶标,挖掘新型抗生素对抗S.suis感染。
合成IPP的甲羟戊酸途径对部分革兰阳性菌的存活非常重要[14]。HMGR是甲羟戊酸途径中的关键限速酶,两类HMGR都被作为药靶研究报道过。有研究者以肺炎链球菌II类HMGR为药靶,筛选出相应的抑制剂[27]。本试验中研究的S.suisHMGR也属于II类HMGR,与肺炎链球菌HMGR氨基酸序列相似性为60%,可以以S.suisHMGR作为药靶,进一步挖掘防控S.suis感染的新型抗生素。本研究原核表达、纯化了S.suisHMGR,并通过分光光度法测定了其最佳反应条件和酶促动力学参数,为下一步筛选S.suisHMGR抑制剂提供方法和参考数据。由于S.suisHMGR三维结构未知,本研究利用同源建模的方法,以已公布的肺炎链球菌HMGR晶体结构为模板,构建了S.suisHMGR可靠的三维结构理论模型。基于其理论三维结构模型,下一步可以通过模拟分子对接,虚拟筛选针对S.suis感染的新型抗生素。本研究中S.suisHMGR酶活力的测定和理论模型的建立为筛选S.suisHMGR抑制剂,防控S.suis感染奠定了基础。
甲羟戊酸途径中5个酶——HMG-CoA合成酶、HMGR、甲羟戊酸激酶、磷酸甲羟戊酸激酶和甲羟戊酸二磷酸脱羧酶,其编码基因被阻断后,限制了病原菌的增殖[14]。因此,除了HMGR外,其他4个酶也可以作为针对多耐药性病原菌的新型抗生素靶标,有进一步研究的价值。例如,肺炎链球菌中HMG-CoA合成酶也被作为潜在的药靶进行初步研究过[28]。病原菌利用甲羟戊酸途径或非甲羟戊酸途径合成IPP,疾病防控中将甲羟戊酸途径和非甲羟戊酸途径中酶的抑制剂联合使用,可以作用于多靶点,抑制不同的致病菌,扩大抗菌谱,具有广阔的应用前景。
4 结 论
原核表达并纯化S.suisHMGR后,测定该酶的活性,确定该酶在催化HMG-CoA还原为甲羟戊酸的反应中的最佳反应温度和pH,以及Vmax和Km值。酶活力的测定为用试验手段验证S.suis抑制剂的有效性提供了可行性的方法。用同源建模的方法构建S.suisHMGR三维结构模型,构建的S.suisHMGR模型将有助于虚拟筛选出对抗S.suis感染的新型抗生素。
[1] WINDSOR R S, ELLIOTT S D. Streptococcal infection in young pigs. IV. An outbreak of streptococcal meningitis in weaned pigs[J].JHyg(Lond), 1975, 75(1): 69-78.
[2] WINDSOR R S. Meningitis in pigs caused byStreptococcussuistype 2[J].VetRec, 1977, 101(19): 378-379.
[3] ERICKSON E D.Streptococcosis[J].JAmVetMedAssoc, 1987, 191(11): 1391-1393.
[4] HIGGINS R, GOTTSCHALK M, MITTAL K R, et al.Streptococcussuisinfection in swine. A sixteen month study[J].CanJVetRes, 1990, 54(1): 170-173.
[5] CHANTER N, JONES P W, ALEXANDER T J. Meningitis in pigs caused byStreptococcussuis-a speculative review[J].VetMicrobiol, 1993, 36(1-2): 39-55.
[6] GOYETTE-DESJARDINS G, AUGER J P, XU J, et al.Streptococcussuis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing[J].EmergMicrobesInfect, 2014, 3 (6): e45.
[7] WERTHEIM H F, NGUYEN H N, TAYLOR W, et al.Streptococcussuis, an important cause of adult bacterial meningitis in northern Vietnam[J].PLoSOne, 2009, 4(6):e5973.
[8] REAMS R Y, GLICKMAN L T, HARRINGTON D D, et al.Streptococcussuisinfection in swine: a retrospective study of 256 cases. Part I. Epidemiologic factors and antibiotic susceptibility patterns[J].JVetDiagInvest, 1993, 5(3): 363-367.
[9] HEUSTON S, BEGLEY M, GAHAN C G, et al. Isoprenoid biosynthesis in bacterial pathogens[J].Microbiology, 2012, 158(Pt6):1389-1401.
[10] REUSCH V M Jr. Lipopolymers, isoprenoids, and the assembly of the gram-positive cell wall[J].CritRevMicrobiol, 1984, 11(2):129-155.
[11] MATSUMI R, ATOMI H, DRIESSEN A J, et al. Isoprenoid biosynthesis in Archaea-biochemical and evolutionary implications[J].ResMicrobiol, 2011, 162(1): 39-52.
[12] BOUCHER Y, DOOLITTLE W F. The role of lateral gene transfer in the evolution of isoprenoid biosynthesis pathways[J].MolMicrobiol, 2000, 37(4): 703-716.
[13] SETO H, WATANABE H, FURIHATA K. Simultaneous operation of the mevalonate and non-mevalonate pathways in the biosynthesis of isopentenyl diphosphate inStreptomycesaeriouvifer[J].TetrahedronLett, 1996, 37(44): 7979-7982.
[14] WILDING E I, BROWN J R, BRYANT A P, et al. Identification, evolution, and essentiality of the mevalonate pathway for isopentenyl diphosphate biosynthesis in gram-positive cocci[J].JBacteriol, 2000, 182(15):4319-4327.
[15] WILDING E I, KIM D Y, BRYANT A P, et al. Essentiality, expression, and characterization of the Class II 3-hydroxy-3-methylglutaryl coenzymeA reductase ofStaphylococcusaureus[J].JBacteriol, 2000, 182(18): 5147-5152.
[16] BOCHAR D A, STAUFFACHER C V, RODWELL V W. Sequence comparisons reveal two classes of 3-hydroxy-3-methylglutaryl coenzymeA reductase[J].MolGenetMetab, 1999, 66(2):122-127.
[17] MIZIORKO H M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis[J].ArchBiochemBiophys, 2011, 505(2): 131-143.
[18] LAWRENCE C M, RODWELL V W, STAUFFACHER C V. Crystalstructure ofPseudomonasmevaloniiHMG-CoA reductase at 3.0 angstrom resolution[J].Science, 1995, 268(5218):1758-1762.
[19] ISTVAN E S. Bacterial and mammalian HMG-CoA reductases: related enzymes with distinct architectures[J].CurrOpinStructBiol, 2001, 11(6): 746-751.
[20] ALBERTS A W, CHEN J, KURON G, et al. Mevinolin: a highly potent competitive inhibitor of hydroxymethylglutaryl-coenzyme A reductase and a cholesterol-lowering agent[J].ProcNatlAcadSciUSA, 1980, 77(7): 3957-3961.
[21] LI W, HU X Y, LIU L, et al. Induction of protective immune response againstStreptococcussuisserotype 2 infection by the surface antigen HP0245[J].FEMSMicrobiolLett, 2011, 316(2): 115-122.
[22] 李 鹏, 陈兰英. 血脂康抑制猪肝HMG-CoA还原酶的活力[J]. 基础医学与临床, 2003, 23(5): 531-534.
LI P, CHEN L Y. Xuezhikang inhibits the activity of HMG-CoA reductase in pig liver[J].BasicMedicalSciencesandClinics, 2003, 23(5): 531-534.(in Chinese)
[23] LI K B. ClustalW-MPI: ClustalW analysis using distributed and parallel computing[J].Bioinformatics, 2003, 19(12): 1585-1586.
[24] BIASINI M, BIENERT S, WATERHOUSE A, et al. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information[J].NucleicAcidsRes, 2014, 42: 252-258.
[25] LASKOWSKI R A, MACARTHUR M W, MOSS D S, et al. PROCHECK: a program to check the stereochemical quality of protein structures[J].JApplCryst, 1993, 26(2): 283-291.
[26] FENG Y J, ZHANG H M, WU Z W, et al.Streptococcussuisinfection an emerging/reemerging challenge of bacterial infectious diseases?[J].Virulence, 2014, 5(4): 477-497
[27] LI D, GUI J, LI Y J, et al. Structure-based design and screen of novel inhibitors for class II 3-hydroxy-3-methylglutaryl coenzyme A reductase fromStreptococcuspneumoniae[J].JChemInfModel, 2012, 52(7): 1833-1841.
[28] BEN Y L, CUI G Z, LI C, et al. Expression, purification, characteristics and homology modeling of the HMGS fromStreptococcuspenumoniae[J].BiomedEnvironSci, 2009, 22(3): 229-236.
(编辑 白永平)
Prokaryotic Expression, Activity Assay and Homology Modeling of the 3-hydroxy-3-methylglutaryl Coenzyme A Reductase fromStreptococcussuis
LIU Can-ying1, XU Zhuo-fei2, FU Qiang1
(1.DepartmentofLifeSicenceandEngineering,FoshanUniversity,Foshan528000,China; 2.CollegeofAnimalScienceandVeterinaryMedicine,HuazhongAgriculturalUniversity,Wuhan430070,China)
The aim of this study was to take theStreptococcussuis(S.suis) 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGR) as a drug target, and provide theoretical and experimental basis for the development of novel antibiotics for the prevention and treatment ofS.suisinfection. In this experiment,S.suisHMGR was prokaryotically expressed and purified by Ni-NTA agrose. Its enzyme activity was determined by spectrophotometric method. In addition, a multiple sequence alignment of HMGR was performed by CLUSTALW to identify conserved domains. Homology modeling of the three-dimensional (3D) structure ofS.suisHMGR was performed by SWISS-MODEL. The quality of the model was evaluated by PROCHECK. The result of enzyme activity analyses showed that optimal reaction conditions ofS.suisHMGR was pH 5.0 and 37 ℃, theVmaxandKmvalue was 846 U·mg-1and 0.213 mmol·L-1respectively. The analysis of SWISS-MODEL showed that crystal structure ofStreptococcuspneumoniaHMGR was the appropriate template for homology modeling ofS.suisHMGR. The theoretic model of 3D structure ofS.suisHMGR was proven to be reliable through quality assessment. TheS.suisHMGR model will contribute to virtual screening of novel antibacterial agents againstS.suisinfection. Enzyme activity assay provides a feasible method for validation of effective inhibitors againstS.suisHMGR in biochemical experiment.
Streptococcussuis; 3-hydroxy-3-methylglutaryl coenzyme A reductase; prokaryotic expression; protein expression; enzyme activity assay; homology modeling
10.11843/j.issn.0366-6964.2016.11.021
2016-06-22
广东普通高校青年创新人才项目(2015KQNCX173);广东省自然科学基金-博士启动(2014A030310020)
刘璨颖(1986-),女,湖北武汉人,助教,博士生,主要从事基础兽医学研究,E-mail: liucy3032@163.com
S852.61
A
0366-6964(2016)11-2318-07
