华南家鸡MHC-B区域复合微卫星位点LEI0258的遗传多样性与进化研究
2016-12-13黄勋和李丽芝张金枫何丹林张细权陈洁波李威娜杜炳旺钟福生
黄勋和,李丽芝,张金枫,何丹林,张细权,陈洁波,李威娜,杜炳旺,钟福生*
(1. 嘉应学院生命科学学院,梅州 514015; 2. 华南农业大学动物科学学院,广州 510642;3. 广东海洋大学农学院,湛江 524088)
华南家鸡MHC-B区域复合微卫星位点LEI0258的遗传多样性与进化研究
黄勋和1,李丽芝1,张金枫1,何丹林2,张细权2,陈洁波1,李威娜1,杜炳旺3,钟福生1*
(1. 嘉应学院生命科学学院,梅州 514015; 2. 华南农业大学动物科学学院,广州 510642;3. 广东海洋大学农学院,湛江 524088)
旨在应用MHC-B区域复合微卫星位点LEI0258研究华南家鸡的遗传多样性与进化历史。以华南地区和邻省家鸡及商品鸡为试验材料,对微卫星位点LEI0258进行基因分型和测序,分析其遗传多样性和构建中介网络图。结果表明,580份样品共检测到LEI0258的62个等位基因,长度范围为182~501 bp,其中37个为首次发现。华南家鸡LEI0258保持着较高的遗传多样性水平,观察杂合度、期望杂合度和多态信息含量分别为0.813、0.960和0.957。中山沙栏鸡、文昌鸡和五华三黄鸡的遗传多样性水平明显高于广西三黄鸡。LEI0258的28个等位基因与27种已知MHC血清型相对应。基于LEI0258侧翼区变异信息的中介网络图将62个等位基因分为8条进化枝,其中3条进化枝同样存在于红原鸡中,提示华南家鸡主要起源于红原鸡。部分等位基因存在于特定鸡品种中,可作为品种鉴定的参考依据。结果提示,微卫星位点LEI0258适用于地方鸡群体遗传多样性、血清型鉴定、品种鉴定和进化研究。
主要组织相容性复合体;LEI0258;华南家鸡;多态性;进化;血清型
地方家禽遗传资源是生物多样性的重要组成部分,是未来家禽品种改良和适应生产条件变化、保持生产长期持续发展的遗传基础[1]。研究家禽遗传多样性,综合评估品种遗传资源现状与保护潜力,有助于揭示各品种间的历史渊源及其亲缘关系,对合理利用和保护地方品种、优良基因库具有重要的意义,是遗传资源管理不可或缺的一个重要环节[2]。微卫星(Microsatellite)又称短串连重复(Short tandem repeats,STR)或简单序列重复(Simple sequence repeats,SSR),是评价遗传多样性与进化最常用的遗传标记之一。曲鲁江等利用27个微卫星位点评估了中国78个地方鸡品种的遗传多样性,为中国地方鸡品种的利用和保护提供了重要的理论依据[3]。杜胭脂等利用27个微卫星位点对山东汶上芦花鸡群体进行遗传分析,结果显示该群体遗传多样性偏低,建议采取适当的保护措施加以控制[4]。
微卫星位点LEI0258是近期逐渐应用于鸡遗传多样性与进化研究的分子标记[5-8]。与常规的选择中性微卫星标记不同,LEI0258定位于重要功能基因——鸡16号染色体上的主要组织相容性复合体B区域(Major histocompatibility complex B region,MHC-B)[9]。MHC-B与宿主许多传染性疾病(如马可氏病)的抗性或易感性等有关[10-11]。传统采用通过B区血清反应的方法对MHC进行血清型鉴定[12-13]。然而,由于该方法主要基于白来航鸡等商业品种[14],限制了在其他鸡品种尤其是家鸡上的应用,也不利于不同研究之间的比较[8]。微卫星位点LEI0258与MHC血清型有很好的对应关系[15],通过研究LEI0258可解决传统血清型鉴定方法的局限性,因而获得了广泛的关注[5-8]。
华南地区(广东省、香港特别行政区、澳门特别行政区、广西壮族自治区、海南省和福建省中南部)有着丰富的地方鸡遗传资源,有14种地方鸡品种入选中国畜禽遗传资源志[1]。前期微卫星标记研究表明华南地区地方鸡保留着较高的遗传多样性水平[3,16-17],但基于LEI0258的系统研究较少[7,18]。本研究以华南地区和邻省家鸡及商品鸡为试验材料,利用微卫星位点LEI0258分析家鸡的遗传多样性,界定LEI0258等位基因与MHC血清型的关系,探讨利用LEI0258侧翼变异信息作为品种鉴定依据的可行性,同时进一步了解LEI0258的进化动态,从而为华南家鸡的基础研究和保种选育提供理论依据。
1 材料与方法
1.1 试验材料
以10个华南地区鸡种和2个邻省家鸡品种,以及2个商品鸡种共580份样品为试验材料(表1)。五华三黄鸡入选《中国禽类遗传资源》[19],其余11种家鸡均入选《中国畜禽遗传资源志·家禽志》[1]。岭南黄鸡为华南地区常见商品鸡,隐性白洛克常用于新品系培育。五华三黄鸡、阳山鸡样品采自其原产地,其余10个家鸡品种采自保种场,商品鸡采自养鸡场。样品类型包括血液和羽毛。DNA基因组采用试剂盒HiPure Tissue DNA Mini Kit(美基生物,广州)提取,-20 ℃保存备用。
1.2 PCR扩增与基因分型
微卫星位点LEI0258使用引物LEI0258F(5′-CACGCAGCAGAACTTGGTAAGG-3′)和LEI0258R(5′-AGCTGTGCTCAGTCCTCAGTGC-3′)扩增[9]。正向引物5′端加上FAM荧光标记。PCR反应体系为20 μL:1×PCR buffer,200 μmol·L-1dNTP mixture,正、反向引物各200 nmol·L-1,1 UTaqDNA聚合酶(TaKaRa, 大连),50 ng DNA模板。扩增条件:94 ℃预变性4 min;然后35个循环(94 ℃变性30 s,65 ℃退火1 min,72 ℃延伸1 min);最后72 ℃延伸10 min。PCR产物在ABI 3730分析仪进行基因型判定(上海翼和应用生物技术有限公司)。
1.3 微卫星测序
基于基因分型结果挑选特定等位基因进行测序。每个等位基因挑选1~3个样品进行测序;如果一个等位基因分布于多个品种中,则挑选不同品种的样品进行测序。需测序等位基因割胶纯化后连接T载体(TaKaRa, 大连),然后转入大肠杆菌DH5α,LB培养基培养过夜,经LEI0258引物PCR鉴定后送广州艾基生物技术有限公司双向测序。
1.4 数据分析
等位基因数(NA)、观察杂合度(Observed heterozygosity,HO)、期望杂合度(Expected heterozygosity,HE)和多态信息含量(Polymorphic information content,PIC)由软件Cervus3.0.3[20]执行。为减少因样本量差异干扰等位基因多态性的判读,采用稀疏的方法(Rarefaction)计算等位基因丰度[Ar(g)]和私有等位基因丰度[Ap(g)],g代表在一个群体内最少的样本数,在软件ADZE1.0[21]执行。序列由软件BioEdit7.2.5[22]编辑处理,并辅以人工校验。在NCBI核酸数据库(http://www.ncbi.nlm.nih.gov/nuccore/)搜索本研究鉴定的LEI0258的等位基因是否与已发表的相同。序列比对在MEGA6.0[23]进行,使用软件SplitsTree4.10构建基于LEI0258侧翼序列的中介网络图(Median-joining network)[24]。
2 结 果
2.1 LEI0258等位基因多态性
580份样品经基因分型共检测到72个等位基因,长度为181~507 bp,其中等位基因275频率最高(103,0.089),其次是205(94,0.081)、310(75,0.065)和495(67,0.058)。大多数等位基因都分布于南方家鸡中,而等位基因448只在商品鸡中分布。如表1所示,微卫星位点LEI0258在家鸡显示出较高的遗传变异水平,大部分鸡种的HO高于0.8,但
表1 鸡群体微卫星位点LEI0258的遗传变异分析
Table 1 Genetic diversity of LEI0258 for 12 indigenous chicken breeds and 2 commercial chicken breeds
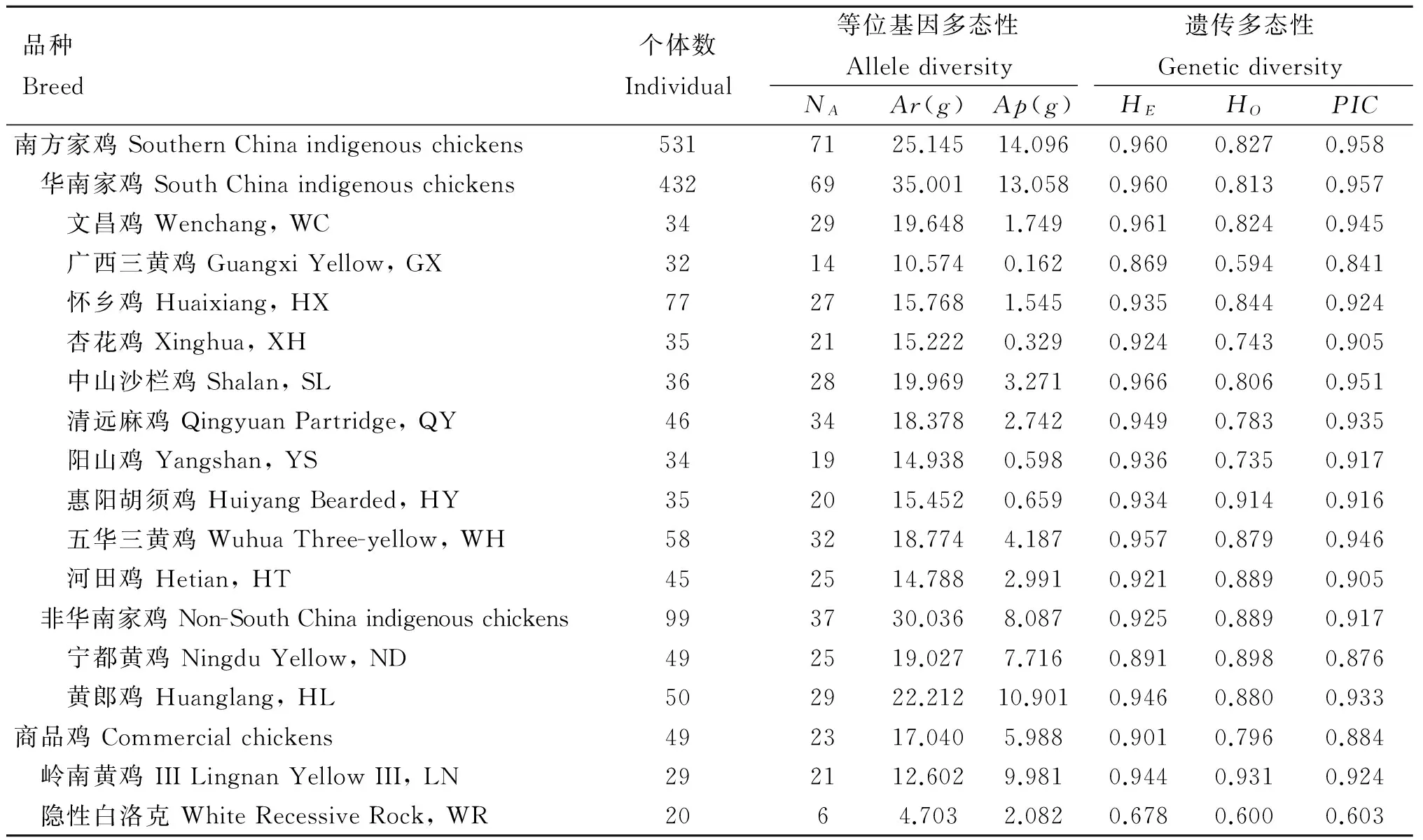
品种Breed个体数Individual等位基因多态性Allelediversity遗传多态性GeneticdiversityNAAr(g)Ap(g)HEHOPIC南方家鸡SouthernChinaindigenouschickens5317125.14514.0960.9600.8270.958 华南家鸡SouthChinaindigenouschickens4326935.00113.0580.9600.8130.957 文昌鸡Wenchang,WC342919.6481.7490.9610.8240.945 广西三黄鸡GuangxiYellow,GX321410.5740.1620.8690.5940.841 怀乡鸡Huaixiang,HX772715.7681.5450.9350.8440.924 杏花鸡Xinghua,XH352115.2220.3290.9240.7430.905 中山沙栏鸡Shalan,SL362819.9693.2710.9660.8060.951 清远麻鸡QingyuanPartridge,QY463418.3782.7420.9490.7830.935 阳山鸡Yangshan,YS341914.9380.5980.9360.7350.917 惠阳胡须鸡HuiyangBearded,HY352015.4520.6590.9340.9140.916 五华三黄鸡WuhuaThree-yellow,WH583218.7744.1870.9570.8790.946 河田鸡Hetian,HT452514.7882.9910.9210.8890.905 非华南家鸡Non-SouthChinaindigenouschickens993730.0368.0870.9250.8890.917 宁都黄鸡NingduYellow,ND492519.0277.7160.8910.8980.876 黄郎鸡Huanglang,HL502922.21210.9010.9460.8800.933商品鸡Commercialchickens492317.0405.9880.9010.7960.884 岭南黄鸡IIILingnanYellowIII,LN292112.6029.9810.9440.9310.924 隐性白洛克WhiteRecessiveRock,WR2064.7032.0820.6780.6000.603
NA.等位基因数:Ar(g) .等位基因丰度;Ap(g). 私有等位基因丰度;南方家鸡对商品鸡g为49,华南家鸡对非华南家鸡g为99,华南家鸡内g为32,非华南家鸡内g为49,商品鸡内g为20.HE.期望杂合度;HO.观察杂合度;PIC. 多态信息含量
NA. Total number of alleles;Ar(g),Ap(g). Allelic and private allelic richness, respectively; With rarefaction for a corresponding sample size of 49 for Southern China indigenous chickensvs. commercial chickens, 99 forSouthChina indigenous chickensvs. non-SouthChina indigenous chickens, 32 within South China indigenous chickens, 49 within non-SouthChina indigenous chickens, 20 for within commercial chickens.HE. Expected heterozygosity;HO. Observed heterozygosity;PIC. Polymorphic information content
广西三黄鸡明显低于其他地方鸡种;所有家鸡的PIC高于0.8。经稀疏处理后,华南家鸡的等位基因丰度高于非华南家鸡,私有等位基因丰度高于非华南家鸡。在华南家鸡群体中,广西三黄鸡的等位基因丰度最低,中山沙栏鸡和文昌鸡的最高;但五华三黄鸡的私有等位基因丰度最高,中山沙栏鸡次之,广西三黄鸡最低。在非华南家鸡中,黄郎鸡的遗传多样性水平要高于宁都黄鸡。总体而言,商品鸡的遗传多样性要明显低于家鸡,特别是高度商品化的隐性白洛克。
2.2 LEI0258核苷酸多态性
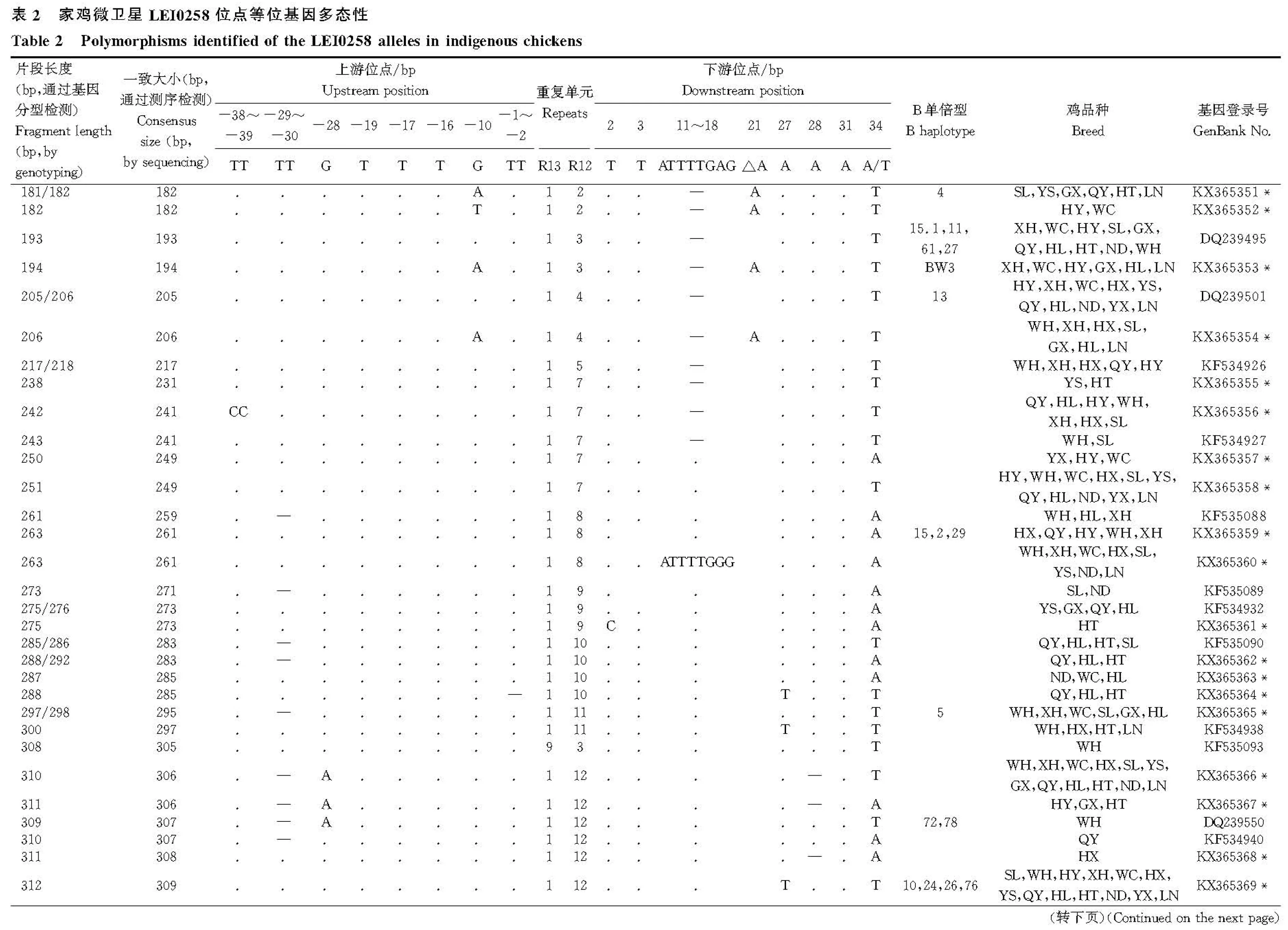
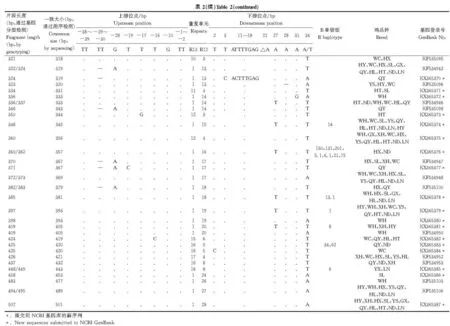
挑选243个片段进行DNA测序,获得了62个等位基因,片段长度为182~501 bp,其中37个等位基因为首次发现(表2)。这些新等位基因序列已提交到NCBI基因库,基因登录号为KX365351~KX365387。基于毛细管电泳分型的等位基因182、193、194、205、206和217与测序结果大小相一致,其他的均小于测序结果。LEI0258有两个典型的重复单元R13(CTATGTCTTCTTT)和R12(CTTTCCTTCTTT),重复次数分别为1~17和2~28,呈现此消彼长的关系,两者的不同组合构成了不同的等位基因。重复区上游有77 bp(包括引物序列),下游76 bp。上游发现8个变异位点,以转换为主,-29~-30 bp出现“TT”缺失的现象。下游发现8个变异位点,以颠换为主。部分等位基因出现下游11~18“ATTTTGAG”的缺失现象,但仅限于小于241 bp的等位基因。11~18 bp位置的多态现象与O.Chazara等[8]和B.Han等[7]的报道相同,但与J.E.Fulton等[15]报道的“ATTTGAGG”有所差异。
2.3 LEI0258侧翼区序列分析
去掉重复区序列后,基于LEI0258侧翼区的变异信息构建中介网络图(图1)。62个等位基因分为8条进化枝(A~H),每条进化枝含1~21个等位基因,E、G和H分别只有1个等位基因,而F则有21个。R13重复次数大于1的等位基因仅限于进化枝C中。在红原鸡中同样发现进化枝A、B和G的等位基因。LEI0258的部分等位基因只出现在单个鸡品种中,如308(怀乡鸡)、367(清远麻鸡)、453(中山沙栏鸡)、477(五华三黄鸡)等;有些只出现在少数鸡品种中,如231(阳山鸡、河田鸡)、357(怀乡鸡、宁都黄鸡)、379(怀乡鸡、清远麻鸡)等;有些是广泛分布,如193、205、249、306、309、369、501等;而443则只分布在商品鸡中。

图1 基于微卫星位点LEI0258侧翼区变异信息的中介网络图Fig.1 Median-joining network of 62 alleles defined within the flanking sequences of LEI0258
27种MHC血清型与28个LEI0258的等位基因相对应,分布于A、B、C、D和F 5条进化枝中,血清型B27在进化枝A出现一次。
3 讨 论
3.1 LEI0258多态性
本研究获得了鸡LEI0258微卫星位点LEI0258的62个等位基因,其中37个未在其他群体中发现[6-8,15]。总体而言,华南家鸡保持着较高的遗传多样性水平,高于许多地方鸡群体如会宁鸡、静宁鸡、丝羽乌骨鸡和商品鸡[8,25]。不同地方品种的遗传多样性有所差异,如广西三黄鸡显著低于其他地方品种,而中山沙栏鸡、文昌鸡和五华三黄鸡最高。这可能与不同品种的选育历史和保育现状有关,如广西三黄鸡经历了长期的选育[26],遗传多样性较低[27];而五华三黄鸡未经系统选育[28-29],因而遗传多样性也较高[30]。此外,商品鸡的遗传多样性明显低于地方家鸡,这是由于商品鸡往往经历了短期高强度的人工选育和较低的有效群体数量的缘故[2],因而在地方鸡品种保种选育过程中,应注意随机交配和保存较大的群体数量。因此,将微卫星位点LEI0258作为研究鸡群体遗传多样性的候选分子标记是可行的。
3.2 LEI0258与血清型
鸡的抗病能力与MHC血清型类型的丰富程度正相关[8,18,31]。本研究确定了LEI0258的28个等位基因的血清型,分别对应于MHC的27种血清型,未发现品种特异的血清型。这些血清型大部分来源于白来航鸡等商品鸡,并已在其他鸡群体中得到了验证[7,15,31]。其他34个等位基因的血清型还有待于确认,将来需开展家鸡的MHC血清型鉴定工作和与抗病能力等的对应关系研究,从而为家鸡的抗病研究提供理论依据。
3.3 LEI0258与品种鉴定
B.Han等研究发现LEI0258稀有等位基因可作为鉴定鸡品种和监测鸡群体保护动态的依据[7]。本研究新发现的37个等位基因是特定鸡品种特有的,如308(怀乡鸡)、367(清远麻鸡)、231(阳山鸡、河田鸡)、357(怀乡鸡、宁都黄鸡)等,说明LEI0258具有作为鸡品种鉴定的候选分子标记的可能性。但由于受限于研究样本,LEI0258私有等位基因作为品种鉴定的依据是否可行还需进一步探讨。同时,随着经贸的发展,不同鸡品种的交流日益频繁,并且不断有新培育品系上市,如岭南黄鸡、新浦东鸡、京海黄鸡等,品种的遗传特性正在逐渐减小,品种鉴定工作难度加大。因此,品种鉴定除了依靠微卫星位点LEI0258外,还需要联合外形特征和其他分子标记,如COI基因[32]、细胞色素b基因[33]等。
3.4 LEI0258与进化
与经典的微卫星位点不同,LEI0258由两个分别为13个碱基和12个碱基的重复单元组成,重复次数多达31次[8,15]。同时侧翼区存在大量的插入缺失和碱基突变,显示出极高的多态性[5-8,15,25]。目前NCBI数据库上LEI0258等位基因有80个,加上本研究新发现的37个,远高于一般的微卫星位点[5-8,15]。相比于微卫星重复区,侧翼区要保守的多,因而侧翼区的变异信息可作为划分等位基因的依据。但需要注意的是,基于侧翼区变异信息构建的中介网络图可能更多反映的是等位基因之间的关系,而非系统发生关系[8]。同时,LEI0258位于MHC-B编码区,受到选择的压力,因而LEI0258也可能处于选择和动态进化中[34]。
中介网络图将本研究鉴定的62个等位基因分为8条进化枝,其中红原鸡同样存在进化枝A、B和G的等位基因,同时与灰原鸡和黑尾原鸡具有一些相同的等位基因,提示南方家鸡主要起源于红原鸡[35-36],但也有其他原鸡的遗传贡献[37]。
4 结 论
华南家鸡在微卫星位点LEI0258上显示出较高的遗传多样性水平,与群体的遗传资源现状相符,说明LEI0258可作为评估鸡群体遗传多样性的分子标记。鉴于LEI0258与MHC血清型关联的特性,通过鉴定LEI0258等位基因,可显著降低血清型分型成本和提高研究效率。LEI0258私有等位基因可作为鸡品种鉴定的参考分子标记,但需联合其他技术手段。另外,从LEI0258侧翼区的变异信息可从侧面验证家鸡的起源驯化。总之,微卫星位点LEI0258是研究地方鸡群体遗传多样性与进化的有力工具,将来需进一步探讨该位点的形成机制和进化动态。
[1] 国家畜禽遗传资源委员会.中国畜禽遗传资源志·家禽志[M].北京:中国农业出版社,2011.
China National Commission of Animal genetic resources. Animal genetic resources in China: Poultry[M]. Beijing: China Agriculture Press, 2011. (in Chinese)
[2] GROENEVELD L F, LENSTRA J A, EDING H, et al. Genetic diversity in farm animals-a review[J].AnimGenet, 2010, 41(S1):6-31.
[3] 曲鲁江,李显耀,徐桂芳,等.利用微卫星标记分析中国地方鸡种的遗传多样性[J].中国科学(C辑:生命科学), 2006,36(1):17-26.
QU L J, LI X Y, XU G F, et al. Evaluation of genetic diversity in Chinese indigenous chicken breeds using microsatellite markers[J].ScienceinChinaSeriesC:LifeScience, 2006, 36(1):17-26. (in Chinese)
[4] 杜胭脂,陈玉霞,康 丽,等.汶上芦花鸡种质资源的遗传多样性分析[J].畜牧兽医学报,2014,45(10):1616-1621.
DU Y Z, CHEN Y X, KANG L, et al. Analysis on the genetic diversity of Wenshang barred chicken breed[J].ActaVeterinariaetZootechnicaSinica, 2014, 45(10):1616-1621. (in Chinese)
[5] IZADI F, RITLAND C, CHENG K M. Genetic diversity of the major histocompatibility complex region in commercial and noncommercial chicken flocks using the LEI0258 microsatellite marker[J].PoultSci, 2011, 90(12):2711-2717.
[6] CHANG C S, CHEN C F, BERTHOULY-SALAZAR C, et al. A global analysis of molecular markers and phenotypic traits in local chicken breeds in Taiwan[J].AnimGenet, 2012, 43(2):172-182.
[7] HAN B, LIAN L, QU L, et al. Abundant polymorphisms at the microsatellite locus LEI0258 in indigenous chickens[J].PoultSci, 2013, 92(12):3113-3119.
[8] CHAZARA O, CHANG C S, BRUNEAU N, et al. Diversity and evolution of the highly polymorphic tandem repeat LEI0258 in the chicken MHC-B region[J].Immunogenetics, 2013, 65(6):447-459.
[9] MCCONNELL S K, DAWSON D A, WARDLE A, et al. The isolation and mapping of 19 tetranucleotide microsatellite markers in the chicken[J].AnimGenet, 1999, 30(3):183-189.
[10] BACON L D, HUNT H D, CHENG H H. Genetic resistance to Marek’s disease[J].CurrTopMicrobiol, 2001, 255:121-141.
[11] LEE L F, BACON L D, YOSHIDA S, et al. The efficacy of recombinant fowlpox vaccine protection againstMarek’s disease: its dependence on chicken line and B haplotype[J].AvianDis, 2004, 48(1):129-137.
[12] BRILES W E, BUMSTEAD N, EWERT D L, et al. Nomenclature for chicken major histocompatibility (B) complex[J].Immunogenetics, 1982, 15(5): 441-447.
[13] FULTON J E, YOUNG E E, BACON L D. Chicken MHC alloantiserum cross - reactivity analysis by hemagglutination and flow cytometry[J].Immunogenetics, 1996, 43(5):277-288.
[14] O’NEILL A M, LIVANT E J, EWALD S J. The chicken BF1 (classical MHC class I) gene shows evidence of selection for diversity in expression and in promoter and signal peptide regions[J].Immunogenetics, 2009, 61(4):289-302.
[15] FULTON J E, JUUL-MADSEN H R, ASHWELL C M, et al. Molecular genotype identification of theGallusgallusmajor histocompatibility complex[J].Immunogenetics, 2006, 58(5-6): 407-421.
[16] ZHANG X Q, LEUNG F C, CHAN D K, et al. Genetic diversity of Chinese native chicken breeds based on protein polymorphism, randomly amplified polymorphic DNA, and microsatellite polymorphism[J].PoultSci, 2002, 81(10): 1463-1472.
[17] YU Y B, WANG J Y, MEKKI D M, et al. Evaluation of genetic diversity and genetic distance between twelve Chinese indigenous chicken breeds based on microsatellite markers[J].IntJPoultSci,2006, 5(6): 550-556.
[18] WANG H Z, MA T, CHANG G B, et al. Molecular genotype identification of different chickens: major histocompatibility complex[J].JSciTechnol, 2014, 2:1-7.
[19] 陈国宏,王克华,王金玉,等.中国禽类遗传资源[M].上海:上海科学技术出版社,2004.
CHEN G H, WANG K H, WANG J Y, et al. Poultry genetic resources in China[M]. Shanghai: Shanghai Scientific and Technical Publishers, 2004. (in Chinese)
[20] KALINOWSKI S T, TAPER M L, MARSHALL T C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment[J].MolEcol, 2007, 16(5):1099-1106.
[21] SZPIECH Z A, JAKOBSSON M, ROSENBERG N A. ADZE: a rarefaction approach for counting alleles private to combinations of populations[J].Bioinformatics, 2008, 24(21): 2498-2504.
[22] HALL T A. BioEdit: a user-friendly biological sequence alignment program for Windows 95/98/NT[J].NucleicAcidsSympSer, 1999, 41:95-98.
[23] TAMURA K, STECHER G, PETERSON D, et al. MEGA6: Molecular evolutionary genetics analysis version 6.0[J].MolBiolEvol, 2013, 30(12):2725-2729.
[24] HUSON D H, BRYANT D. Application of phylogenetic networks in evolutionary studies[J].MolBiolEvol, 2006, 23(2):254-267.
[25] 金丽娜,韩建林.鸡MHC区域内微卫星LEI0258和MCW0312的遗传多态性[J].畜牧与兽医,2015,47(7): 50-55.
JIN L N, HAN J L. Genetic diversity of the microsatellite LEI0258 and MCW0312 in chickenMHC-BRegion[J].AnimalHusbandry&VeterinaryMedicine, 2015,47(7): 50-55. (in Chinese)
[26] 韦凤英,陈宽维,束婧婷,等.广西三黄鸡保护选育和利用研究[J].中国家禽,2011,33(13):26-29.
WEI F Y, CHEN K W, SHU J T, et al. Study on the selection and utilization of Guangxi three-yellow chickens[J].ChinaPoultry, 2011, 33(13):26-29. (in Chinese)
[27] 窦新红,韦凤英,黄 雄,等.广西三黄鸡群体遗传多样性研究[J].中国家禽,2011,33(15):20-23.
DOU X H, WEI F Y, HUANG X, et al. Assessment of genetic variability in Guangxi three-yellow chickens[J].ChinaPoultry, 2011, 33(15):20-23. (in Chinese)
[28] 李威娜,陈云燕,钟福生.广东省五华三黄鸡品种资源保护与利用现状及发展对策[J].湛江师范学院学报, 2011,32(6):132-135.
LI W N, CHEN Y Y, ZHONG F S. Status and development countermeasures of variety resources protection and utilization of Wuhua three-yellow chickens[J].JournalofZhanjiangNormalCollege, 2011, 32(6):132-135. (in Chinese)
[29] 钟福生,李威娜,翁茁先,等.五华三黄鸡品种特性研究[J].中国家禽,2012,34(9):61-63.
ZHONG F S, LI W N, WENG Z X, et al. Breed characterization of Wuhua three-yellow chicken[J].ChinaPoultry, 2012, 34(9):61-63. (in Chinese)
[30] 黄勋和,李威娜,陈 珊,等.五华三黄鸡群体遗传多样性与遗传结构分析[J].中国家禽,2016,38(1):56-58.
HUANG X H, LI W N, CHEN S, et al. Population genetic diversity and genetic structure of Wuhua three-yellow chicken[J].ChinaPoultry, 2016, 38(1):56-58. (in Chinese)
[31] ROGERS S L, KAUFMAN J. High allelic polymorphism, moderate sequence diversity and diversifying selection for B-NK but not B-lec, the pair of lectin-like receptor genes in the chicken MHC[J].Immunogenetics, 2008, 60(8):461-475.
[32] JIN S D, SEO D W, SIM J M, et al. Single nucleotide polymorphism analysis of the COI gene in Korean native chicken[J].KoreanJPoultSci, 2009, 36 (1): 85-88.
[33] YACOUB H A, FATHI M M, SADEK M A. Using cytochrome b gene of mtDNA as a DNA barcoding marker in chicken strains[J].MitochondrialDNA, 2015, 26(2):217-223.
[34] E G X, SHA R N, ZENG S C, et al. Genetic variability, evidence of potential recombinational event and selection of LEI0258 in chicken[J].Gene, 2014, 537(1):126-131.
[35] LIU Y P, WU G S, YAO Y G, et al. Multiple maternal origins of chickens: out of the Asian jungles[J].MolPhylogenetEvol, 2006, 38(1):12-19.
[36] MIAO Y W, PENG M S, WU G S, et al. Chicken domestication: an updated perspective based on mitochondrial genomes[J].Heredity(Edinb), 2013, 110(3): 277-282.
[37] ERIKSSON J, LARSON G, GUNNARSSON U, et al. Identification of the yellow skin gene reveals a hybrid origin of the domestic chicken[J].PLoSGenet, 2008, 4(2):e1000010.
(编辑 郭云雁)
Evaluation of Diversity and Evolution of the Microsatellite LEI0258 in ChickenMHC-BRegion from South China
HUANG Xun-he1,LI Li-zhi1,ZHANG Jin-feng1,HE Dan-lin2,ZHANG Xi-quan2,CHEN Jie-bo1,LI Wei-na1,DU Bing-wang3,ZHONG Fu-sheng1*
(1.SchoolofLifeSciences,JiayingUniversity,Meizhou514015,China; 2.CollegeofAnimalSciences,SouthChinaAgriculturalUniversity,Guangzhou510642,China; 3.CollegeofAgricultural,GuangdongOceanUniversity,Zhanjiang524088,China)
The aim of the study was to investigate polymorphism and evolution in South China chickens by the tandem repeat LEI0258 located within the B region of the chicken major histocompatibility complex (MHC-B region). Genotypes and sequences of LEI0258 were obtained from 12 Southern China chicken breeds and 2 commercial chicken breeds, and then these data were used to analyze genetic diversity and construct median-joining network. A total of 62 alleles were defined by DNA sequencing ranged from 182 to 501 bp in 580 samples, 37 of which was novel for these chicken breeds. The LEI0258 showed high levels of polymorphism in South China chickens, with 0.813, 0.960 and 0.957 for observed heterozygosity, expected heterozygosity and polymorphic information content, respectively. The Shalan, Wenchang and Wuhua Three-yellow chicken breeds had higher levels of genetic diversity than Guangxi Yellow chicken. Twenty-eight alleles of LEI0258 were corresponding to 27 available serotypes of MHC. The 62 alleles of LEI0258 were classified into 8 clusters in median-joining network, based on the SNPs and indels found within the flanking sequences, and 3 of which was found in red junglefowl, indicating that South China chicken mainly originated from red junglefowl. Some alleles existed in a certain chicken breed, which could be as a candidate marker for breed identification. The results suggest that LEI0258 can be used as a molecular marker for elucidate genetic diversity, serology and breed identification, evolution in indigenous chicken breeds.
major histocompatibility complex; LEI0258; South China indigenous chickens; polymorphism; evolution; serotype
10.11843/j.issn.0366-6964.2016.11.004
2016-04-05
广东省自然科学基金项目(2014A030307018);嘉应学院“创新强校工程”项目(CQX019);广东省公益研究与能力建设项目(2015A020208020;2016A030303068)
黄勋和(1982-),男,广东河源人,讲师,博士,主要从事中国家鸡遗传多样性与进化研究,E-mail:hxh826@jyu.edu.cn
*通信作者:钟福生,教授,E-mail:zfs@jyu.edu.cn
S831.2
A
0366-6964(2016)11-2175-09
