Alpha-突触核蛋白寡聚体致帕金森病小鼠模型的行为学变化
2016-12-06王海鹏盖晓东
王 鹏 历 春 王海鹏 盖晓东
(北华大学基础医学院人体解剖学教研室,吉林 吉林 132013)
Alpha-突触核蛋白寡聚体致帕金森病小鼠模型的行为学变化
王 鹏 历 春1王海鹏 盖晓东1
(北华大学基础医学院人体解剖学教研室,吉林 吉林 132013)
目的 观察家族型帕金森病(PD)alpha-突触核蛋白(α-Syn)突变体A53T和A30P寡聚体导致小鼠PD模型的行为学变化,探讨α-Syn寡聚体在体内的神经毒性作用。方法 制备野生型α-Syn、家族型突变体A53T和A30P的聚集体,对C57BL/6小鼠进行纹状体注射。培养至第30、60天,进行爬杆实验、悬吊实验、滚轴实验和平衡木实验评估行为学改变。免疫组织化学方法检测黑质多巴胺能神经元改变。结果 实验鼠纹状体α-Syn寡聚体注射后30 d出现PD样运动障碍表现,纹状体注射30 d和60 d后,各组小鼠的爬杆实验结果均与对照组无差异(P>0.05)。注射30 d后,野生型α-Syn寡聚体组小鼠平衡木实验、悬挂实验、滚轴实验与对照组无差异(P>0.05);A53T和A30P组小鼠平衡木时间比对照组长(P<0.05),滚轴上时间比对照组小鼠缩短(P<0.05),悬挂评分比对照组明显降低(P<0.01),与野生型α-Syn寡聚体组相比有统计学差异(P<0.05)。注射60 d后,野生型α-Syn寡聚体组小鼠平衡木实验平均通过时间比对照组小鼠长(P<0.05)、悬挂实验评分较对照组小鼠降低(P<0.05);A53T和A30P组小鼠各项行为学评估与对照组小鼠差异显著(P<0.01)、与野生型α-Syn寡聚体组相比有差异(P<0.05)。A53T和A30P组小鼠黑质多巴胺能神经元数目下降(P<0.05)。结论 纹状体注射α-Syn突变体A53T和A30P寡聚体,模型小鼠出现PD样行为学改变和黑质多巴胺能神经元减少。
帕金森病;alpha-突触核蛋白;寡聚体
帕金森病(PD)是以神经元内路易体(LB)为主要病理特征的神经变性病〔1〕。而alpha-突触核蛋白(α-Syn)作为LB的主要成分,被认为是引起PD发病的关键性蛋白〔2〕。研究证实,存在于LB中的α-Syn大部分发生错误折叠而聚集〔3〕。外源性α-Syn的寡聚体进入神经元后,可引起进一步聚集,形成不可溶的淀粉样变性包涵体〔4〕。一些家族性PD患者α-Syn基因突变引起α-Syn错译发生结构变化而易于聚集,其中以第88位碱基发生G-C突变,导致氨基酸序列第30位丙氨酸变成脯氨酸(A30P),第209位核苷酸发生G-A突变,使氨基酸序列中第53位丙氨酸变成苏氨酸(A53T)最常见〔5,6〕。我们已证实α-Syn突变型A53T和A30P的寡聚体可以进入神经元,并较野生型α-Syn寡聚体对神经元的毒性更大、在神经元内形成LB样嗜酸性包涵体。如能利用其神经毒性制作动物模型,对PD的发病机制研究意义重大。本研究利用α-Syn寡聚体神经元感染的特点,对实验动物进行纹状体注射,观察其行为学变化,探讨在短时间内构建PD动物模型的可能,并进一步研究α-Syn 家族型突变体A53T和A30P寡聚体在体内的神经毒性作用。
1 材料与方法
1.1 主要试剂 组织包埋剂(SAKURA Tissue-Tek®O.C.T.Compound),购自美国SAKURA公司;α-Syn 单克隆抗体(ab138501),购自Abcam公司;辣根过氧化物酶标记山羊抗小鼠IgG,购自北京中杉金桥生物有限公司;TH抗体,购自美国Abcam公司。
1.2 实验动物 C57BL/6小鼠,雄性,8周龄,重18~22 g,购自北京维通利华实验动物技术有限公司,实验动物生产许可证:SCXK(京)2013-0001。动物订购后,在26 cm×18 cm×18 cm笼箱内、每笼5只适应性培养7 d后,进行体重测量和纹状体注射的预实验操作。之后进行相应实验操作和按照实验动物饲养标准进行正常饲养。动物实验操作流程经实验动物伦理学委员会批准,动物试验操作者持有实验动物人员岗位证书,证书编号39749。共4组,假手术组、野生型α-Syn寡聚体组、A53T寡聚体组、A30P寡聚体组。每组8只。均采用左侧纹状体侧注射。实验组每只注射5 μg寡聚体〔7〕,体积为2 μl,假手术组注射2 μl PBS。
1.3 方法
1.3.1 纹状体注射 小鼠称重,10%水合氯醛0.004 ml/g体重进行腹腔注射麻醉。将小鼠头颅水平位固定在脑立体定位仪上,距离前囟冠状位中线向前0.8 mm、前囟外侧缘2.0 mm、距离硬脑膜深约3.0 mm、开骨窗,注射样品,用牙托粉封闭颅骨孔。然后缝合筋膜和皮肤,并于腹腔注射氨苄青霉素8万U,术后小鼠包被保暖,清醒后置于笼内正常饲养。
1.3.2 动物行为学观察 通过爬杆实验、悬吊实验、滚轴实验和平衡木实验反映肌力、平衡和协调能力。分别在实验前先训练大鼠3 d,然后开始正式实验。在纹状体注射前、注射第30、60天由经过正式培训的实验人员通过双盲原则完成行为学测试。
1.3.2.1 爬杆实验 参照Ogawa等〔8〕的测试方法,自制直径0.8 cm、高60 cm 的直木杆,杆顶部有一小木球,外面覆盖纱布以防止小鼠打滑。小鼠被头向上放于杆顶端,记录动物从开始运动到完全转为头向下的时间和它们下到杆底的时间。
1.3.2.2 悬挂实验 参照Kuribara等〔9〕的测试方法,自制悬挂杆,水平放置,距地面30 cm。实验中小鼠悬挂于金属杆上,如小鼠用两后爪计3分,如用一后爪抓住电线计2分,如小鼠两后爪均抓不住电线计1 分,最后计算得分情况,测5次取平均值。
1.3.2.3 滚轴实验 采用小鼠电动转轴仪(Rotamex-5,Columbus Instruments),滚轴直径6 cm,转速20 r/min,连续测20次取平均值〔10〕。
1.3.2.4 平衡木实验 105 cm × 4 cm × 3 cm长木杆,木杆离地80 cm,两端由木架支撑。实验时,将小鼠放在起始区面对鼠笼方向,同时开始计时,记录小鼠走过起始区的时间(潜伏期)和小鼠走过整个平衡杆的时间〔11〕。先训练3次,然后开始正式实验。走3次取平均值。
1.3.3 免疫组织化学方法 将实验小鼠灌注取脑,制备脑组织冰冻切片。将组织切片放入含0.3% Triton X-100的PBS中,4℃浸泡2~4 d;将组织切片在含有0.5% H2O2的PBS中处理30 min,去除内源性过氧化物酶。在含1% 山羊血清中封闭30 ~ 60 min。加入TH抗体(1∶1 000),4℃过夜。PBST冲洗后,加入生物素化的山羊抗小鼠IgG,1∶1 000,室温孵育2 h。将切片用Tris-HCl缓冲液漂洗后,放入含有DAB和H2O2的显色剂中显色。显色完成后,将切片移入Tris-HCl缓冲液中终止显色。贴片上,低温烤干或自然干燥,然后在不同浓度的乙醇中逐次脱水,每次5 min,然后移入二甲苯中透明,每次3 min。准备好盖玻片,封片剂封片。晾干后,观察。
1.4 统计方法 采用SPSS18.0软件行单因素方差分析,并用Sigma-plot绘图软件作图。
2 结 果
2.1 小鼠给予α-Syn寡聚体后的一般表现 纹状体注射30 d后小鼠开始出现轻微震颤、运动减少,个别出现后肢张开、竖毛等改变;注射A53T和A30P组小鼠运动能力下降明显,震颤增多,肢体僵硬,个别小鼠有步态不稳、反应迟缓的PD样运动障碍表现。

图1 C57BL/6小鼠爬杆实验行为学观察结果
2.2 行为学检测 见图1~图4。纹状体注射30 d和60 d后,各组小鼠的爬杆实验结果均与对照组无差异(P>0.05)。注射30 d后,野生型α-Syn寡聚体组小鼠平衡木实验、悬挂实验、滚轴实验与对照组无差异(P>0.05);A53T和A30P组小鼠平衡木时间比对照组长(P<0.05),滚轴时间比对照组小鼠缩短(P<0.05),悬挂评分比对照组明显降低(P<0.01),与野生型α-Syn寡聚体组相比有统计学差异(P<0.05)。注射60 d后,野生型α-Syn寡聚体组小鼠平衡木实验平均通过时间比对照组小鼠长(P<0.05)、悬挂实验评分较对照组小鼠降低(P<0.05);A53T和A30P组小鼠平衡木实验平均通过时间、悬挂实验评分和滚轴平衡时间均与对照组小鼠有显著差异(P<0.01)、与野生型α-Syn寡聚体组相比有差异(P<0.05)。
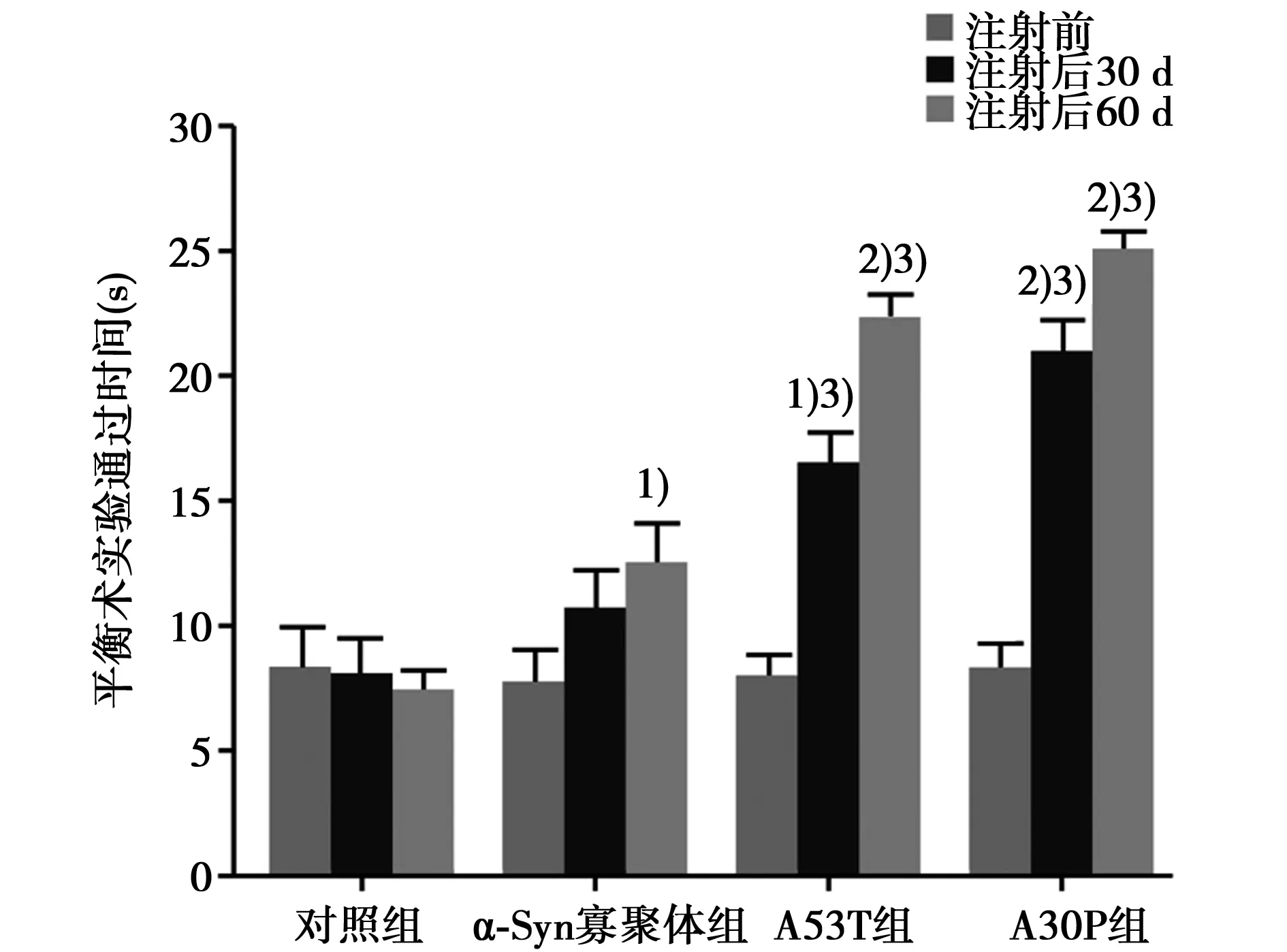
与对照组比较:1)P<0.05,2)P<0.01;与α-Syn寡聚体组比较:3)P<0.05;下图同图2 C57BL/6小鼠平衡木实验行为学观察结果(n=8)

图3 C57BL/6小鼠悬挂实验行为学观察结果(n=8)

与对照组比较:1)P<0.05,2)P<0.01;与A53T组比较:3)P<0.05图4 C57BL/6小鼠滚筒实验行为学观察结果(n=8)
2.3 黑质多皮胺能神经元免疫组织化学检测 纹状体注射第30天各组黑质组织染色正常,未发现多巴胺能神经元减少。第60天,野生型α-Syn寡聚体组小鼠脑黑质多皮胺能神经元较对照组减少约26.53%(P<0.05);A53T和A30P组与对照组相比,黑质多皮胺能神经元数目明显减少约56.34 %和53.21%(P<0.01),存留神经元形态皱缩,突起减少,多巴胺能神经纤维密度减低,见图5。

标尺:20 μm图5 寡聚体组小鼠脑纹状体注射同侧黑质多巴胺神经元较对照组减少
3 讨 论
在神经系统,细胞外源性可溶α-Syn寡聚体进入神经元内后可引起进一步聚集,形成不可溶的淀粉样变性包涵体〔12〕。这些变性蛋白形成的包涵体可以经细胞间的转运感染正常细胞,使病变蔓延〔13〕。向PD患者脑黑质内移植胎儿中脑干细胞,结果显示新移植的神经元细胞内也出现LB样病变〔14〕,进一步证实了LB病变的可转移性。本实验结果证实,在纹状体注射的α-Syn寡聚体可以在神经元间传递的方式进入黑质,并导致黑质多巴胺能神经元的减少。
本文行为学实验结果显示,模型小鼠垂直运动能力未改变而出现悬挂能力减弱、滚筒保持平衡的时间缩短以及通过平衡杆时间延长等表现。在体内证实家族型α-Syn突变体A53T、A30P聚集体对神经元的毒性并具备制作PD模型的可能。良好的动物模型是探讨PD发病机制和治疗方案的基础。已知PD主要发病机制是黑质多巴胺能神经元丧失、纹状体多巴胺含量降低,而使胆碱能系统的功能相对亢进,造成纹状体DA和乙酰胆碱递质失衡。PD模型主要通过损毁的办法破坏黑质-纹状体DA系统,继而产生PD样递质生化改变;药物干扰Ach和DA的平衡,直接模拟PD的递质变化;或通过转基因动物,从发病机制、生化改变以接近人PD改变。但模型均未有PD特征性的病理变化LB形成。我们利用α-Syn寡聚体和家族型α-Syn突变体A53T和A30P进行纹状体注射,发现模型小鼠出现PD样行为学改变和黑质多巴胺能神经元减少,提供了PD模型制备的新方法。
1 Irizarry MC,Growdon W,Gomez-Isla T,etal.Nigral and cortical Lewy bodies and dystrophic nigral neurites in Parkinson′s disease and cortical Lewy body disease contain alpha-synuclein immunoreactivity〔J〕.J Neuropathol Exp Neurol,1998;57(4):334-7.
2 Baba M,Nakajo S,Tu PH,etal.Aggregation of alpha-synuclein in Lewy bodies of sporadic Parkinson′s disease and dementia with Lewy bodies〔J〕.Am J Pathol,1998;152(4):879-84.
3 Conway KA,Lee SJ,Rochet JC,etal.Acceleration of oligomerization,not fibrillization,is a shared property of both alpha-synuclein mutations linked to early-onset Parkinson′s disease:implications for pathogenesis and therapy〔J〕.Proc Natl Acad Sci U S A,2000;97(2):571-6.
4 Uversky VN,Li J,Fink AL.Evidence for a partially folded intermediate in alpha-synuclein fibril formation〔J〕.J Biol Chem,2001;276(14):10737-44.
5 Lou H,Montoya SE,Alerte TN,etal.Serine 129 phosphorylation reduces the ability of alpha-synuclein to regulate tyrosine hydroxylase and protein phosphatase 2A in vitro and in vivo〔J〕.J Biol Chem,2010;285:17648-61.
6 Liu D,Jin L,Wang H,etal.Silencing alpha-synuclein gene expression enhances tyrosine hydroxylase activity in MN9D cells〔J〕.Neurochem Res,2008,33:1401-9.
7 Luk KC,Kehm VM,Zhang B,etal.Intracerebral inoculation of pathological α-synuclein initiates a rapidly progressive neurodegenerative α-synucleinopathy in mice〔J〕.J Exp Med,2012;209(5):975-86.
8 Ogawa N,Hirose Y,Ohara S,etal.A simple quantitative bradykinesia test in MPTP-t reated mice〔J〕.Res Commun Chem Pathol Pharmacol,1985;50(3):435-41.
9 Kuribara H,Higuchi Y,Tadokoro S.Effects of central depressants on rota-rod and traction performances in mice〔J〕.Jpn J Pharmacol,1977;27(1):117-26.
10 Wozniak DF,Xiao M,Yamada KA,etal.Impaired spatial learning and defective theta burst induced LTP in mice lacking fibroblast growth factor 14〔J〕.Neurobiol Dis,2007;26:4-26.
11 Fu XH,Zhu M,Sun XY,etal.Original article hyperbaric oxygen treatment and enteral nutrition support with glutamine relieves traumatic brain injury in the rats〔J〕.Int J Clin Exp Med,2014;7(12):5686-90.
12 Uversky VN,Li J,Fink AL.Evidence for a partially folded intermediate in alpha-synuclein fibril formation〔J〕.J Biol Chem,2001;276(14):10737-44.
13 Clavaguera F,Bolmont T,Crowther RA,etal.Transmission and spreading of tauopathy in transgenic mouse brain〔J〕.Nat Cell Biol,2009;11(7):909-13.
14 Kordower JH,Chu Y,Hauser RA,etal.Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson′s disease〔J〕.Nat Med,2008;14:504-6.
〔2015-12-19修回〕
(编辑 徐 杰)
吉林省科技厅项目(20150101128JC)
盖晓东(1962-),女,教授,硕士生导师,主要从事肿瘤分子病理学研究。
王 鹏(1977-),男,博士,讲师,主要从事人体解剖学和神经生物学基础理论研究。
R74
A
1005-9202(2016)21-5227-04;
10.3969/j.issn.1005-9202.2016.21.005
1 北华大学基础医学院病理学教研室
