甲羟戊酸激酶基因真核及shRNA表达载体构建及其对BxPC-3细胞周期蛋白表达的影响
2016-11-28张胜权
金 锐,王 芳,栾 康,罗 欣,张胜权
甲羟戊酸激酶基因真核及shRNA表达载体构建及其对BxPC-3细胞周期蛋白表达的影响
金 锐,王 芳,栾 康,罗 欣,张胜权
目的 构建甲羟戊酸激酶(MVK)基因真核及shRNA表达载体并转染至BxPC-3细胞中,观察其对细胞周期蛋白的影响。 方法 从BxPC-3细胞中提取总RNA, 逆转录PCR(RT-PCR)法获得目的基因MVK以及通过化学合成MVK(751~779)和MVK(1 089~1 117)位点寡核苷酸片段,构建真核表达及shRNA表达载体并转染至BxPC-3细胞,筛选稳定表达细胞系。 Western blot法分析转染后对BxPC-3细胞周期蛋白的影响。 结果 成功获得MVK基因真核表达(pcDNA3.1-mvk)及两个针对MVKshRNA表达载体(piLenti-RNAi-shRNA1/2)。通过抗生素筛选及Western blot分析获得了稳定的MVK过表达及低表达细胞株。Western blot分析显示:过表达MVK的细胞株及MVKknockdown的细胞株中细胞周期蛋白Cyclin B1、Cyclin E表达量与正常及对照细胞相比均显著降低。 结论MVK过表达及低表达均抑制BxPC-3细胞周期蛋白Cyclin B1、Cyclin E的表达。
甲羟戊酸激酶;基因重组;寡核苷酸;细胞周期蛋白;BxPC-3;shRNA
甲羟戊酸途径是体内胆固醇生物合成的重要代谢途径[1],其主要是由乙酰辅酶A为原料合成甲羟戊酸,然后在甲羟戊酸激酶(mevalonate kinase,MVK)的催化作用下最终生成法尼基焦磷酸(FPP)、牛耳基牛儿焦磷酸(GGPP)、胆固醇(CH)等。中间代谢产物FPP、GGPP是细胞蛋白质酯化的戊二烯供体,参与调节细胞增殖、凋亡、分化等过程[2]。研究[3]证实:几乎存在一半以上肿瘤细胞中的p53能够显著上调细胞中甲羟戊酸途径代谢及蛋白质的酯化过程,提示甲羟戊酸途径可能成为肿瘤治疗的一个新靶点。胰腺癌是常见的消化系统恶性肿瘤,患者死亡率高。Harrison et al[4]研究发现苯乙酸盐能够抑制甲羟戊酸途径中类异戊二烯的生物合成进而对胰腺癌细胞有生长抑制作用。真核细胞进行增殖主要依靠有丝分裂方式。分裂周期分为:间期和分裂期。细胞周期的顺利进行需要细胞周期蛋白(Cyclins)及周期蛋白依赖性激酶(CDK)进行调节。当甲羟戊酸代谢途径紊乱,可能对胰腺癌细胞周期蛋白合成产生影响最终产生生长抑制。该研究通过体外构建不同基因表达载体研究其对胰腺癌BxPC-3细胞周期蛋白的作用及机制,旨在为胰腺癌治疗提供一个新的思路。
1 材料与方法
1.1 材料
1.1.1 基因重组试剂 TRIzol试剂、逆转录试剂盒、DNA上样缓冲液(6×Loading Buffer、DNA Marker(DL2 000等)、PCR反应试剂、Hind Ⅲ和XhoⅠ均购自日本TaKaRa公司;T4 DNA连接酶、限制性核酸内切酶BbsI购自美国NEB公司;质粒提取试剂盒购自美国OMEGA公司;胶回收试剂盒购自美国AxyGen公司。
1.1.2 细胞培养及转染试剂 DMEM、1640、胎牛血清购自美国Gibco公司;G418和嘌呤霉素(Puromycin)购自美国Sigma公司;MegaTran1.0 Transfection Reagent购自美国Origene公司。
1.1.3 细胞、质粒、菌株 胰腺癌细胞BxPC-3购自中国科学院细胞中心; pcDNA.3.1/myc-His A, B, and C、piLenti-RNAi-GFP、E.coliJM109均购自美国Invitrogen公司。
1.2 方法
1.2.1 引物设计 根据NCBI GenBank 提供的MVK序列,设计MVK真核表达引物。 上游引物(带有Hind Ⅲ酶切位点):5′-CCCAAGCTTATGTTGTCAGAAGTCCTACTGG-3′; 下游引物(带有Xho Ⅰ酶切位点):5′-CCGCTCGAGTCAGAGGCCATCCAGGGCTTGC-3′。利用shRNA在线软件设计两条针对MVK基因 mRNA的shRNA序列。针对MVK(751-779)的序列:5′-AGAAACAGGCTGCTCAAGTTCCCAGAGAT-3′和MVK(1 089~1 117)的序列:5′-TGGCTTTGACTGCTTGGAAACCAGCATCG-3′。寡核苷酸序列由上海生物工程有限公司合成。1.2.2 HaCaT细胞培养收集及总RNA提取 复苏HaCaT细胞,用含10%胎牛血清的DMEM于37 ℃、5% CO2的培养箱中培养24 h,待其状态良好,弃去培养液,PBS洗3次,0.25%胰酶消化,收集细胞悬液,TRIzol试剂提取细胞总RNA。
1.2.3 RT-PCR扩增野生型MVK基因
1.2.3.1 逆转录反应 采用逆转录试剂盒进行逆转录反应,依次加入总RNA 5 μl,10×RT Buffer 1 μl,Oligo dT 0.5 μl,10 mmol/L dNTP 1 μl,25 mmol/L MgCl22 μl,AMV Reverse Transcriptase 0.25 μl。轻弹管底使其混匀,点动离心,42 ℃ 60 min, 95 ℃ 5 min条件下逆转录,加入40 μl ddH2O即为cDNA备用溶液。
1.2.3.2 PCR反应 取上述cDNA 5 μl,依次加入TaqDNA Polymerase 1 μl、上下游引物各0.5 μl、10 mmol/L dNTP 0.5 μl、10×Buffer 2.5 μl、25 mmol/L MgCl21.5 μl,补ddH2O 8.5μl。94 ℃ 5 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,72 ℃ 10 min,循环30次。琼脂糖凝胶电泳分析结果。
1.2.4 重组体的构建及筛选
1.2.4.1MVK基因真核表达载体的构建、筛选 将质粒pcDNA.3.1和MVK在37 ℃的恒温体系中分别用核酸内切酶Hind Ⅲ和XhoⅠ双酶切4 h。琼脂糖凝胶电泳分析酶切结果,用胶回收试剂盒回收所需酶切产物。pcDNA.3.1和目的片段按5︰1摩尔比加入T4 DNA连接酶体系中,16 ℃连接过夜。-80 ℃取出感受态JM109细胞, 将上述重组体转化至JM109中,37 ℃培养12 h,菌落扩增后经PCR、酶切进行重组体鉴定,测序。-20 ℃保存,备用。重组体命名为pcDNA 3.1-mvk。
1.2.4.2MVK不同位点shRNA表达载体的构建、筛选 合成的oligoDNA序列在缓冲体系中依次经过变性、退火反应。质粒piLenti-RNAi-GFP及oligoDNA经Bbs I酶切、电泳、回收,以5 ∶1的摩尔比16 ℃连接过夜。将连接产物转化入JM109,经抗生素及PCR筛选出阳性克隆,扩增,测序,提取质粒。-20 ℃保存,备用。重组体命名为piLenti-RNAi-mvk1/2。
1.2.4.3 重组体转染至BxPC-3细胞 复苏冻存的人胰腺癌BxPC-3细胞,于含10%胎牛血清的1640培养基、37 ℃、5% CO2中培养,待其状态良好,按2×105/ml密度接种于12孔板中培养,待其达到90%~95%汇合时,用MegaTran1.0 Transfection Reagent将重组质粒转染至BxPC-3细胞中,设置复孔培养24 h后,分别用G418和Puromycin抗性培养基筛选。同时设置空白未处理及转染空载pcDNA3.1、piLenti-RNAi-GFP细胞组作为对照。
1.2.4.4 Western blot 检测MVK基因表达产物 6组细胞(空白未处理细胞、pcDNA3.1、pcDNA3.1-mvk、piLenti-RNAi-GFP、piLenti-RNAi-mvk1/2稳定转染的细胞)培养后,提取细胞总蛋白进行SDS-PAGE凝胶电泳,按10 μl /孔上样量加入12%凝胶孔中,依次进行:电泳(浓缩胶60 V,分离胶120 V约2 h)、转移(150 mA恒流3 h左右)、封闭(5%脱脂牛奶常温封闭2 h)、孵育一抗(4 ℃孵育过夜)、孵育二抗(常温摇床孵育2 h)、Thermo Scientific SuperSignal West Pico Trial Kit化学发光底物显影,分析MVK基因表达产物。
1.2.4.5 Western blot 检测细胞周期蛋白表达 通过1.2.4.4步骤鉴定出MVK基因高表达及knockdown的细胞,接种24孔培养板,Western blot步骤同1.2.2.4,化学发光分析各组细胞中细胞周期蛋白(Cyclin B1及Cyclin E)的表达。

2 结果
2.1 PCR扩增MVK基因编码区(CDS)及化学合成shRNA 经RT-PCR获得大量MVK基因全长片段,1.5%琼脂糖凝胶电泳可见1 190 bp左右特异性扩增条带,见图1A。
2.2MVK真核表达重组体的构建与鉴定MVK基因经酶切、回收、连接、转化后。PCR及双酶切筛选阳性克隆,结果可见1 190 bp的MVKPCR产物,酶切后可见5 500 bp的pcDNA3.1及1 190 bp的MVK片段,见图1A、1B 。阳性克隆扩增后测序,结果经Blast比对正确(图1C,给出MVK的部分序列),保存备用。
2.3 shRNA表达载体的构建与鉴定 shRNA经酶切、电泳、回收、转化、连接、转化后,经酶切筛选阳性克隆(图1D)。扩增后测序,结果经Blast比对正确(图1D,给出shRNA1的序列),保存备用。
2.4 转染BxPC-3观测GFP荧光和筛选稳定表达细胞系 piLenti-RNAi-GFP-mvk(751-779,shRNA1)及piLenti-RNAi-GFP-mvk(1 089~1 117, shRNA2)转染BxPC-3经Puromycin(0.5 μg/ml) 筛选,荧光镜下结果见图2, Western blot分析显示shRNA1稳定转染的细胞中MVK蛋白表达显著降低,见图3。
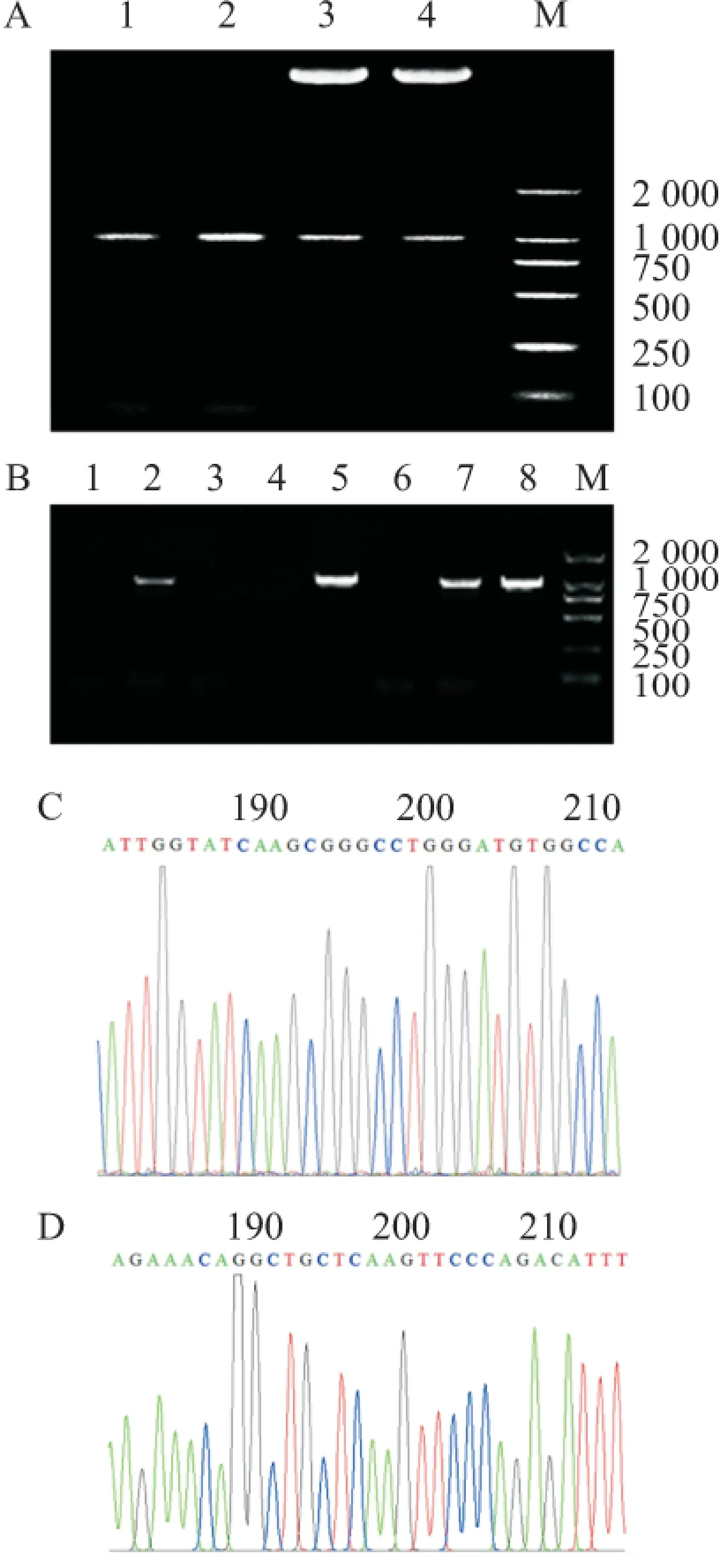
图1 MVK真核表达载体及shRNA表达载体构建
M: Marker;A:MVK全长片段获取及重组体筛选;1、2:MVKPCR扩增产物;3、4:MVK重组体酶切结果;B: PCR筛选MVK重组体克隆; 1、3、4、6:阴性克隆;2、5、7、8:阳性克隆;C:MVK测序结果(部分序列);D: shRNA1 测序结果(部分序列)
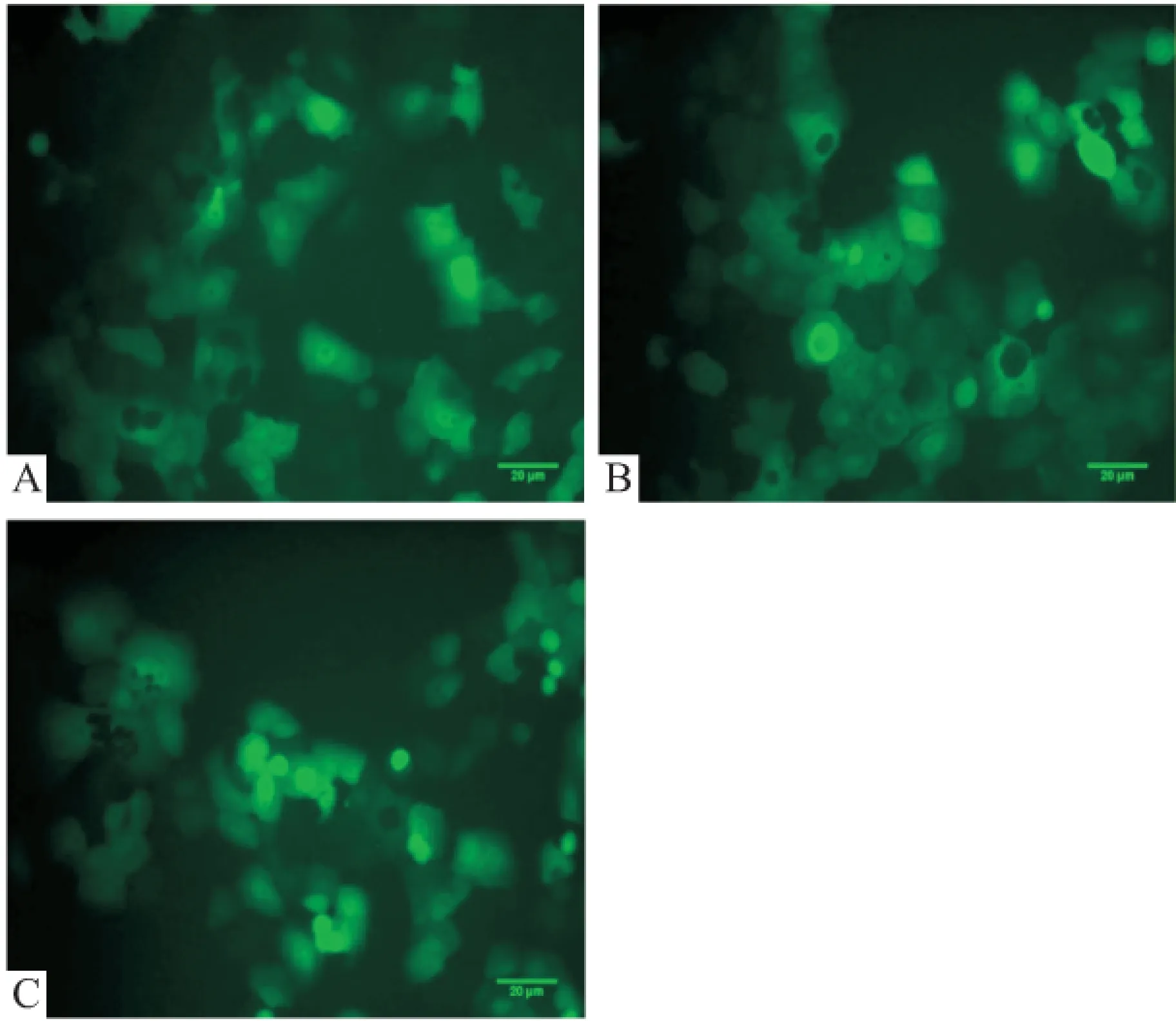
图2 荧光筛选MVKshRNA转染稳定表达细胞系
A: 转染piLenti-RNAi-GFP组; B: 转染mvkshRNA1组; C: 转染mvkshRNA2组

图3 Western blot分析MVK及Cyclins结果
A:空白未处理组;b:转染pcDNA3.1组;c:转染pcDNA3.1-mvk组;d:转染piLenti-RNAi-GFP组; e:转染 piLenti-RNAi-GFP-mvk(751-779, shRNA1) 组;f:piLenti-RNAi-GFP-mvk(1 089~1 117,shRNA2) 组;与空白未处理组比较:*P<0.05;与转染pcDNA3.1组比较:#P<0.05;与转梁piLenti-RNAi-GFP组比较:△P<0.05
pcDNA3.1-mvk转染BxPC-3后,用抗生素G418(500 μg/ml)筛选稳定表达细胞株,Western blot结果显示pcDNA3.1-mvk稳定转染的细胞中MVK蛋白表达显著升高,见图3。
2.5 Western blot分析转染后MVK基因在BxPC-3中表达情况以及对BxPC-3细胞周期蛋白表达的影响 a到f组细胞各指标分别进行方差分析,SPSS 17.0软件分析得出FMVK=716.86,FCyclinB1=55.346,FCyclinE=93.321,各组均为P>0.05,即方差齐性,对各指标分别进行t检验,结果显示:转染pcDNA3.1-mvk组、空白未处理组、转染pcDNA3.1组相比,MVK相对表达量显著升高,细胞周期蛋白(Cyclin B1、Cyclin E)相对表达量显著降低,差异有统计学意义(P<0.05)。转染MVKshRNA1/shRNA2组与空白未处理组、转染piLenti-RNAi-GFP组相比,MVK相对表达量显著降低,细胞周期蛋白(Cyclin B1、Cyclin E)相对表达量也显著降低,差异有统计学意义(P<0.05)。见图3。
3 讨论
甲羟戊酸代谢途径是体内合成脂类物质的一种重要代谢途径,参与细胞的生理进程,几乎存在于所有高等真核生物和病毒中[5]。其中MVK起到关键作用,是合成下游重要营养物质的桥梁[6]。研究[7-8]表明甲羟戊酸基因突变将导致MVK缺失症,是一种稀有的常染色体代谢性疾病。
本研究通过体外构建MVK基因的真核表达载体转染至人胰腺癌细胞BxPC-3,旨在研究MVK基因不同程度表达对其增殖相关蛋白的影响。通过体外研究发现,转染含有野生型MVK基因的重组体使BxPC-3细胞MVK基因高表达,并在很大程度上降低细胞周期蛋白Cyclin B1、Cyclin E的表达水平;转染MVK基因的shRNA1/2均能下调BxPC-3细胞MVK基因的表达水平且不同程度地抑制其周期蛋白Cyclin B1、Cyclin E的表达。该研究采用基因工程及knockdown技术构建不同基因表达载体来干预胰腺癌细胞MVK基因表达程度,从而为更详细地研究甲羟戊酸途径在胰腺癌中的作用机制打下了一个新的基础。
纵观甲羟戊酸整个代谢途径,当MVK缺失后将会导致下游的FPP、GGPP、CH等的缺失及上游甲羟戊酸的累积。其中FPP、GGPP是细胞蛋白质酯化的戊二烯供体[9],细胞内蛋白质的异戊二烯化使定位于细胞膜上的GTP酶介导细胞信号的转导,从而调控细胞的生长及凋亡进程。其中,研究最多的就是小G蛋白参与的细胞调控机制。常见的小G蛋白如Ras、Rho、Rab、Ran家族等,它们是细胞内多个信号转导的中心环节,因此参与对细胞生理功能的影响,并与抗肿瘤治疗有关[10]。代谢中产生的CH是细胞膜系统的重要组成成分,在细胞的生理功能各个方面扮演着非常重要的角色。由此可以推断,MVK基因突变后导致MVK功能缺失,造成下游产物FPP、GGPP等缺乏,导致细胞蛋白酯化过程不能进行,从而可能影响细胞周期蛋白及其蛋白因子表达,进而阻碍细胞的生长。而当MVK活性过度增强时,将很大程度加快代谢反应的进程,促使下游产物FPP、GGPP等合成持续增加,促进细胞蛋白质酯化过程加快过度,从而可能不同程度地影响细胞的信号转导造成生理功能紊乱,如降低周期蛋白的表达水平,表现为细胞生长抑制,其深入的分子机制有待进一步研究。因此,下一步将重点进行MVK正常表达、低表达和高表达情况下,BxPC-3细胞增殖相关情况及其相同条件下小G蛋白酯化情况及其介导的信号通路方面的研究。
[1] Rauthan M, Pilon M. The mevalonate pathway inC.elegans[J]. Lipids Health Dis, 2011, 10: 243.
[2] Joo J H, Jetten A M. Molecular mechanisms involved in farnesol-induced apoptosis[J]. Cancer Lett, 2010, 287(2):123-35.
[3] Thurnher M, Nussbaumer O, Gruenbacher G. Novel aspects of mevalonate pathway inhibitors as antitumor agents [J]. Clin Cancer Res, 2012, 18(13):3524-31.
[4] Harrison L E, Wojciechowicz D C, Brennan M F, et al. Phenylacetate inhibits isoprenoid biosynthesis and suppresses growth of human pancreatic carcinoma[J]. Surgery, 1998, 124(3):541-50.
[5] Gelb M H, Brunsveld L, Hrycyna C A, et al. Therapeutic intervention based on protein prenylation and associated modifications[J]. Nat Chem Biol, 2006,2(10):518-28.
[6] Haas D, Hoffmann G F. Mevalonate kinase deficiencies: from mevalonic aciduria to hyperimmunoglobulinemia D syndrome[J]. Orphanet J Rare Dis, 2006,1:13.
[7] Tricarico P M, Marcuzzi A, Piscianz E, et al. Mevalonate kinase deficiency and neuroinflammation: balance between apoptosis and pyroptosis[J]. Int J Mol Sci, 2013, 26; 14(12):23274-88.
[8] De Leo L, Marcuzzi A, Decorti G, et al. Targeting farnesyl-transferase as a novel therapeutic strategy for mevalonate kinase deficiency:invitroandinvivoapproaches[J]. Pharmacol Res, 2010, 61(6):506-10.
[9] Buhaescu I, Izzedine H. Mevalonate pathway: a review of clinical and therapeutical implications[J]. Clin Biochem, 2007, 40(9-10):575-84.
[10]Galle M, Crespo R, Kladniew B R, et al. Suppression by geraniol of the growth of A549 human lung adenocarcinoma cells and inhibition of the mevalonate pathway in culture andinvivo: potential use in cancer chemotherapy[J]. Nutr Cancer, 2014, 66(5):888-95.
The construction of eukaryotic and shRNA expression vector of mevalonate kinase gene and its effect on cyclins expression in BxPC-3
Jin Rui, Wang Fang, Luan Kang, et al
(Dept of Biochemistry and Molecular Biology, Anhui Medical University, Hefei 230032)
Objective To construct the eukaryotic expression and shRNA expression vectors of mevalonate kinase (MVK) gene and study the effect on cyclins expression in BxPC-3 cells. Methods Total RNA was extracted from BxPC-3 cells, and the desired geneMVKwas obtained by RT-PCR and acquiring chemical synthesisMVK(751-779) andMVK(1089-1117) locus oligo fragments. After construction of eukaryotic and shRNA expression vectors using genetic engineering, the recombinant was transfected into BxPC-3 and cell lines of stable expression were selected by antibiotic. Western blot was used to analyze the expression ofMVKand cyclins in BxPC-3. ResultsMVKeukaryotic (pcDNA3.1-mvk) and two shRNA (piLenti-RNAi-shRNA1/2) expression vectors forMVKwere successfully constructed. The cell lines with stableMVKover-expression and knockdown were obtained by antibiotic selection and Western blot. Western blot results showed that Cyclin B1 and Cyclin E expressed significantly lower both in BxPC-3 cell withMVKover-expression andMVKknockdown compared with the control. Conclusion BothMVKover expression and knockdown can inhibit the expression of Cyclin B1, Cyclin E in BxPC-3.
mevalonate kinase; genetic recombination; oligos; cyclins; BxPC-3 cell; shRNA
时间:2015-12-30 14:38
http://www.cnki.net/kcms/detail/34.1065.R.20151230.1438.012.html
国家自然科学基金(编号:81271748)
安徽医科大学生物化学与分子生物学教研室,合肥 230032
金 锐,男,硕士研究生;
张胜权,男,教授,硕士生导师,责任作者,E-mail:sqz36@yahoo.com
R 349.83
A
1000-1492(2016)01-0022-05
2015-10-12接收
