武夷山不同龄级香果树开花物候及其生殖特性
2016-10-28郭连金
郭连金, 李 梅, 林 盛, 谢 鹏
( 上饶师范学院, 江西 上饶 334000; 福建武夷山国家级自然保护区管理局, 福建 武夷山 354300 )
武夷山不同龄级香果树开花物候及其生殖特性
郭连金1*, 李梅1, 林盛2, 谢鹏2
( 上饶师范学院, 江西 上饶 334000; 福建武夷山国家级自然保护区管理局, 福建 武夷山 354300 )
该研究采用固定样地观测研究了不同龄级香果树的开花物候和生殖特性,分析了物候指数与生殖构件之间的相关性。结果表明:香果树单花花期一般为5~9 d,平均为6.79 d,随着树龄增大,其单花花期呈幂函数增长趋势;不同树龄香果树的始花期不同,但终花期基本一致;随着树龄的增大,始花期逐渐提前,花期持续时间延长。20~50 a树龄的香果树始花期约为8月9日,花期持续时间最短,仅为36 d。110~140 a树龄的香果树始花期约为7月15日,花期持续时间为61 d;对于单株母树来讲,香果树的花枝数,花朵总数以及果实总数随着树龄的增大均显著增加;单花枝产花量随着树龄的增加呈上升趋势,单花枝产果实数量呈先上升后下降的趋势;香果树花枝和单花枝产花量在树冠方位上的分布由大到小依次为南>东/西>北、上>中>下,但单花枝果实数量则表现出不同规律,即东>南>西>北、中>下>上。香果树具有花多果少现象,开花物候指数与生殖构件数量间的相关性分析表明,其花期持续时间与花枝数、花数和果实数均存在极显著正相关关系,始花期与生殖构件数量呈极显著负相关关系,而物候指数与香果树树龄亦存在显著相关关系。
保护植物, 生殖构件, 单花花期, 生殖成功
开花是植物从营养分生组织转变为生殖分生组织的生理过程,在植物的生产和物种进化中起到重要作用(罗睿等, 2010)。其物候是植物重要的生活史特征之一,研究植物的单花花期、开花期、花期持续时间等开花物候可以了解植物的自身特性及其对环境的适应性(Ollerton et al, 1992)。而植物生殖构件分布和数量特征则为有性生殖是否成功的标志,关系到种群的更新与维持(张文辉等, 2002; Manuel et al, 2002),是植物个体和种群水平上生殖生态学研究的新方向(祖元刚等, 2000)。近年来,国内外学者对植物的开花物候和生殖构件方面进行了研究,张文标等(2008)研究发现濒危植物夏蜡梅(Calycanthuschinensis)为大量开花模式,不同群体和海拔对其开花物候具有显著影响;焦培培等(2007)从花、花序、个体和居群水平上对濒危植物矮沙冬青(Ammopiptanthusnanus)的开花物候进行了研究,认为矮沙冬青开花同步性较低,单花开放时间为4~13 d;肖宜安等(2005)对野生长柄双花木(Disanthuscercidifoliusvar.longipes)的开花物候和生殖构件特征进行了为期4 a 的跟踪观测,发现其个体开花持续时间为49~55 d,种群花期历时63~71 d,其纯林中生殖构件的数量高于毛竹林中生殖构件的数量;何淼等(2014)对侧金盏花(Adonisamrensis)种群的开花物候进行了观测,认为其花期持续时间为20~25 d,开花数目和座果率呈显著正相关,而与始花期呈显著负相关;柴胜丰等(2005)对金花茶的开花物候和生殖构件特征进行研究,发现其生殖构件数量与其个体的径级及树冠的层次有一定关系。尽管不少学者从个体、居群等水平对植物的开花物候和生殖构件进行了研究,但多数研究把开花物候和生殖构件数量的变化归于积温的影响(Blionis et al, 2001),其研究对象多为随机选取(张文标等, 2008; 何淼等, 2014)或选择大小一致的个体(基茎、高度)进行研究(肖宜安等, 2005)。植物的开花物候和生殖构件除了受外界环境影响外,同时也受其内部遗传因素的影响(罗睿等, 2010)。未达到生殖年龄的植物即使在适宜环境中,也无法实现开花结实,且不同生殖年龄会影响其开花时间及生殖构件数量(祖元刚等, 2010),而有关此方面的研究鲜见报道(张大勇, 2004)。研究不同年龄植物的开花物候及生殖构件特征有助于进一步了解植物有性繁殖这一复杂过程,对于珍稀濒危植物而言则可以为研究其生殖生态学特征及濒危机制提供参考,为种群恢复及有效经营管理提供理论依据。
香果树(Emmenopteryshenryi)属茜草科,是第四纪冰川幸存孑遗植物之一,为中国特有单种属植物,是研究茜草科系统发育、形态演化及中国植物地理区系的重要材料。近年来,由于其自然更新能力差,加上环境恶化和人为破坏等因素的影响,该物种已濒临灭绝,被列为国家Ⅱ级重点保护稀有植物和林业部公布的国家珍贵树种(傅立国等, 1992)。日前,由于香果树种群数量的急剧减少以及我国对生物多样性保护的重视,引起了我国一些学者对香果树的关注。李铁华等(2004)和康华靖等(2011)对香果树种子休眠和萌发进行了研究,结果表明香果树种子属于光敏感性种子,20 ℃为最佳萌发温度;Li et al(2008)研究表明香果树的遗传多样性较低,其基因流较低可能是导致香果树濒危的原因之一;Du et al(2010)对香果树愈伤组织经长期继代后的同工酶酶谱进行了研究,结果显示其胚性愈伤组织和非胚性愈伤组织在6种同工酶水平上均有差异。本课题组于2008年对武夷山不同生境的香果树种群的开花物候、生殖构件特征进行初步研究,发现同一生境中不同个体的开花期、持续时间以及生殖构件数量等差异较大。但由于研究内容不同、观测时间短以及不同树龄的香果树数量较少等原因,没有阐述不同树龄香果树的开花物候和生殖构件特征,而这是植物有性生殖过程中的两个关键的环节,对了解种群自然更新机制,阐明植物濒危原因有重要作用。本研究于2010-2014年通过野外踏查,扩大研究范围,增加研究对象,对武夷山不同树龄香果树的开花物候、花枝、花、果等生殖构件分布格局及数量特征进行了持续观测研究。旨在探讨:(1) 不同树龄香果树种群开花物候的变化;(2) 不同树龄香果树种群繁殖格局及其空间差异性;(3) 香果树开花物候与生殖成功的关系等问题。目的是阐明香果树种群有性生殖基本特征及其与树龄的关系,为其种群恢复提供依据。
1 研究地自然概况
研究地位于武夷山国家级自然保护区(27°33′~27°54′ N,117°27′~117°51′ E),总面积为56 527.3 hm2,是世界同纬度现存面积最大,保存最完整的中亚热带森林生态系统.区内自然资源丰富,保存有完好的地带性常绿阔叶林群落,是东南亚大陆生物多样性最丰富的地区,也是具有全球生物多样性保护意义的地区,本区气候属于典型的亚热带季风气候,平均气温12~13 ℃,年降水量1 480~2 150 mm,年蒸发量1 492 mm左右,相对湿度78%~84%,无霜期253~272 d。土壤类型主要有红壤、黄红壤、黄壤和山地草甸土(郭连金, 2009)。香果树在研究地主要星散分布于沟谷水溪旁的阔叶林中,其伴生种主要有银鹊树(TapisciaSinensis)、杉木(Cunninghamialanceolata)、红脉钓樟(Linderarubronervia)、盐肤木(Rhuschinensis)、野鸭椿(Euscaphisjaponica)、毛竹(Phyllostachyspubescens)、亮叶红淡(Adinandranitida)、伞形绣球(Hydrangeaumbellata)、一年蓬(Erigeronannuus),兔儿伞(Syneilesisaconitifolia)、黄堇(Corydalisamabilis)、菝葜(Smilaxchina)、糯米团(Gonostegiahirta)等。
2 研究方法
2.1 样地设置
香果树大多星散分布,其开花植株相距较远,为减小生境对其有性生殖的影响,本研究选择在其分布范围最大、生境相对一致的地段进行研究。由于其大多沿沟谷和溪流分布,故设置4个20 m × 100 m的长形样地进行调查(表1)。调查内容包括:① 生境:海拔,坡向,坡位、土壤含水量、光照、大气温度、大气湿度等,其中土壤含水量采用便携式土壤水分速测仪(TRIME-TD RZ)进行测定,林冠层光照采用照度计(ZDH-10)测定,林内的温、湿度采用通风干湿度温度计(DMH2)测定(选择前两天无雨天气,林内光照、温湿度等的测定于每日10:00、13:00、15:00时进行,连续测定3 d)。② 群落学特征:树种组成、高度、盖度等。③ 香果树特征,对样地内香果树母株进行调查,记录其数量、胸径、树高等。④香果树树龄的确定:利用瑞典生长锥对样地中一部分香果树个体进行年轮分析,确定其树龄,后经胸径和树龄拟合,获得方程(y= 12.172e0.0353x),该方程的拟合度较好(R2= 0.9798,P=0.000),能较准确地通过香果树的胸径推算出其树龄(图1)。

图 1 武夷山香果树胸径与树龄的拟合方程Fig. 1 Fitting equation of DBH and age of E. henryi tree in Wuyi Mountain
2.2 开花物候及生殖构件的调查与统计
2010年6月份,随机标记40株具有花蕾的香果树作为研究对象,按照树龄将20~50 a为Ⅰ龄级,50~80 a为Ⅱ龄级、80~110 a为Ⅲ龄级、110~140 a为Ⅳ龄级,每个龄级选择10棵植株(表2)。由于香果树存在2-4 a开花一次的现象,故于2011-2014年每年6月份选择同年龄的40株具有花蕾的香果树作为研究对象,为了使研究具有延续性,以先前标记过的香果树作为优先选择对象。在每株树冠上随机选取30个花蕾并做好标记,待标记的花蕾开放后,每天观察1次,直至花朵脱落,记录其每朵花持续时间作为单花花期。将所标记香果树树冠均分为上、中、下3层,东、南、西、北4个方向,统计其生殖构件分布及数量特征。由于香果树花序为圆锥状聚伞花序,本研究采用标准枝法进行花枝、花、果调查,每个标准枝长1 m,包括当年生、2 a和3 a生的枝条,在香果树树冠不同方位随机选择5个标准枝(不够的按实际枝数),通过人工攀爬标记每个标准枝。2010-2014年,自所标记植株第一朵花开放开始,每隔3 d观察1次,直至其落叶进入自然休眠期,记录同一标准枝上的花枝、花和果实数量的变化情况,根据单株花数量变化,统计不同龄级香果树种群的始花期、花期持续时间和终花期(马文宝等, 2008)。由不同树冠方向和层位所含标准枝的总数量,估算标准枝在各层及方向所占百分比,来推断整株树冠不同方向、不同层位的花枝数量、开花数量和果实数量,计算花枝、花和果实的比例。

表 1 武夷山香果树种群样地概况

表 2 不同龄级香果树形态指标
某方向(层位)生殖构件数量比例=所调查单株的某方向(层位)生殖构件总数/所有植株的某方向(层位)生殖构件总数
2.3 数据分析
所有数据均以算数平均值为统计数,运用SPSS 19.0软件进行方差分析、LSD多重比较以及双变量相关性分析,并用Origin 9.0软件作图。
3 结果与分析
3.1 香果树单花花期
野外调查发现,武夷山香果树的最低开花树龄为23 a,开花植株的树龄主要集中于30-90 a。由图2知,香果树单花花期随着龄级的增加而呈增长趋势,四个龄级间的单花花期均值存在显著差异。香果树单花花期平均为6.79±0.89 d,最短为5 d,最长为9 d,变异系数为0.13。
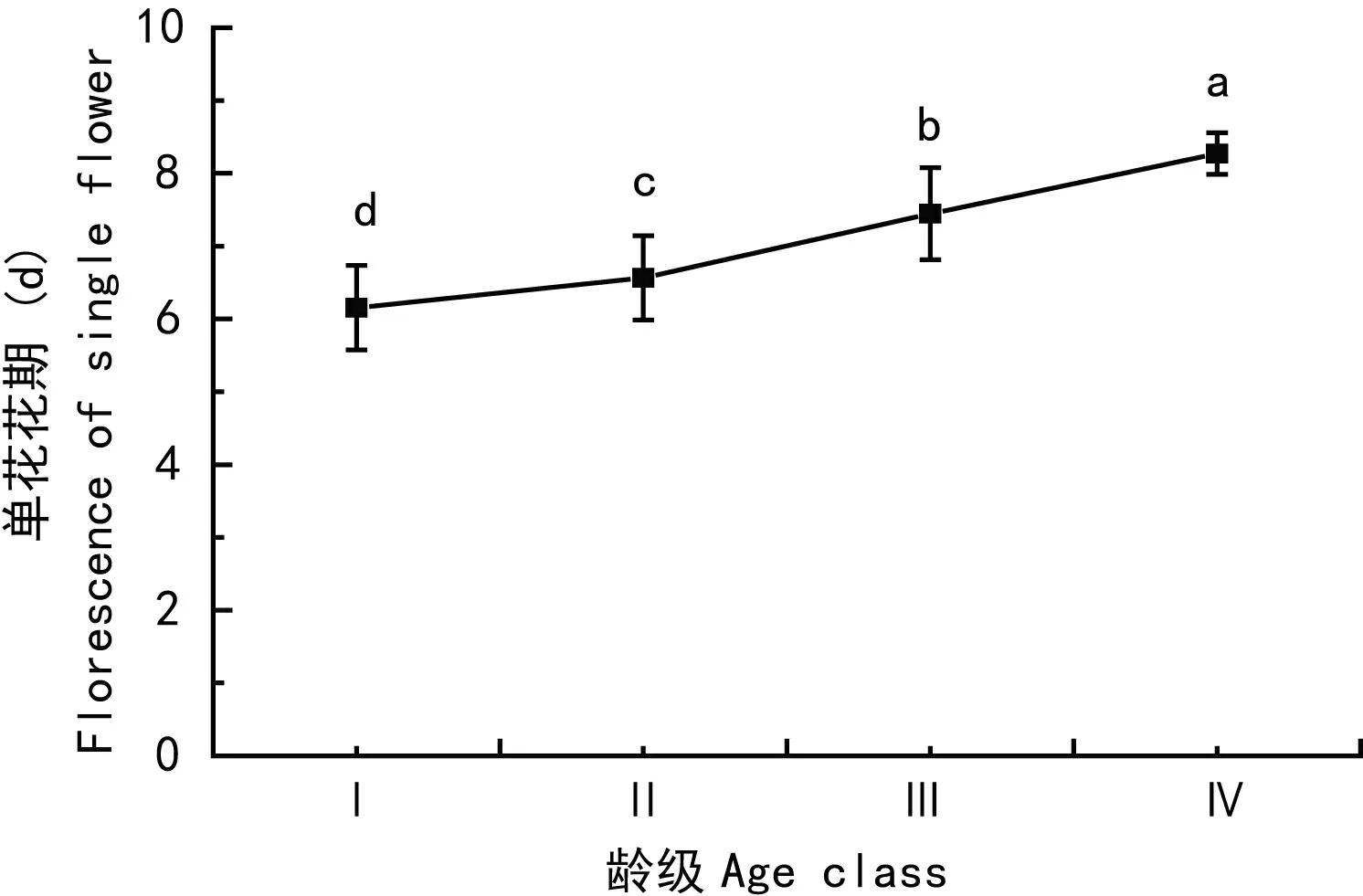
图 2 不同龄级香果树单花花期的变化 a, b, c and d 表示数据在0.05水平上存在显著差异。Fig. 2 Change trend of single flower florescence of E. henryi of different age classes a, b, c and d represent significant differences (P<0.05).
3.2 香果树开花期变化
不同龄级香果树开花进程存在一定差异(图3)。

图 3 不同龄级香果树开花进程 i. Ⅰ龄级植株; ii. Ⅱ龄级植株; iii. Ⅲ龄级植株; iiii. Ⅳ龄级植株。Fig. 3 Flowering course of E. henryi tree of different age classes Age of i, ii, iii and iiii trees represent Ⅰ, Ⅱ, Ⅲ and Ⅳ age class respectively.
由图3:i可知,与其它龄级相比,Ⅰ龄级香果树开花时间相对较晚,为每年8月9号左右,且花期持续时间最短,约为36 d;随着龄级的增加(图3:ii、iii、iiii),香果树始花期逐渐前移,花期持续时间逐渐延长;Ⅳ龄级香果树始花期为每年7月15号左右,花期持续时间可达61 d(图3: iiii);不同龄级香果树终花期基本一致,在每年9月14号左右结束。单花枝日均开花高峰基本位于花期中部。单花枝最高日开花数为10朵/d,最低可连续2 d不开花。
3.3 不同龄级香果树生殖构件分布
不同龄级香果树生殖构件数量差异显著,总体呈现花多果少格局(每73~202朵花形成1个果实)。Ⅰ龄级香果树花枝数、花数/花枝和果数/花枝最少。随着龄级增大,三者数量均逐渐增加。不同龄级香果树的花枝数、单花枝产花数及单花枝座果数均存在显著差异(表3)。单株花枝数量、花总量以及果实总量均随着树龄增加而呈增长趋势。但其变异系数则有所不同,随着树龄增加,三者的变异系数均呈现出先减小后增加的趋势。随着树龄增加,花果转化率逐步增加,而后呈下降趋势(表3)。
3.4 树冠方位对香果树生殖构件数量的影响
由表4可知,香果树生殖构件分布具有向光性,不同方向和层位生殖构件数量差异均达显著水平。花枝和单花枝产花数量由高到低依次为南>西>东>北、上>中>下,即植株树冠阳面和半阳面生殖构件分布较多,阴面分布较少,而单花枝产果量则与之不同,树冠东侧和树冠中层显著高于其他位置。
3.5 香果树开花物候与生殖成功的相关性
香果树的树龄、单花花期、始花期、花期持续时间、花枝数、开花数、果实数之间的相关关系分析结果(表5)显示:香果树树龄与其开花物候及生殖构件数量存在极显著的相关关系,除了与始花期存在极显著负相关以外,其他均为极显著正相关;始花期与其他所有指标均呈极显著负相关,花期持续时间与花枝数、花数和果实数相互间均存在极显著正相关关系。这些关系表明,香果树树龄越大,单花花期越长,始花期越早,花期持续时间越长;花枝数多、开花数多的香果树比花枝数少的个体结果量多;花期持续时间越长的个体产果量越多;始花期较早的个体开花、结果量较大,但单花开放时间较短。
4 讨论
4.1 开花物候特征及其变异
分布于不同地区或同一地区不同海拔 (柴胜丰等, 2009; Butt et al, 2015; 任崇勇等, 2015; Ehrlén et al, 2015),甚至同一地区不同微生境(Hammad et al, 1997)的同种植物开花物候存在一定差异,这主要是由于生境中光照和温度等作用的结果(罗睿等, 2010),但同一生境中不同林龄的同种植物始花期有所不同(祖元刚等, 2000)。本研究中,武夷山香果树单花花期平均为6.79 d,随着树龄的增加,单花花期呈幂函数上升趋势;始花期则随着香果树树龄的增加而逐渐前移,即较低树龄的香果树始花期较晚,而搞树龄的香果树始花期较早,这一结果与白桦类似,其壮年个体的始花期早于刚结实的个体(祖元刚等, 2000)。本研究中,香果树开花时间最早始于每年的7月中旬,最晚始于8月初,但由于终花期基本一致(9月上旬),所以香果树树龄越大,花期持续时间越长。由于植物开花是一个复杂的形态建成过程,是其体内各种因素与环境因子相互作用的结果,其中自身遗传物质起决定作用(吴莉英等, 2007)。较低树龄的香果树为由营养生长转为生殖生长这一过渡阶段的个体,体内各种成化物质可能不完备,需要较长时间的积累,导致其始花期较晚,这种开花物候的变异具有较强的遗传力(Geber et al, 2003; Hendry et al, 2005)。

表 3 不同龄级香果树生殖构件数量特征
注: 数据进行LSD检验,a、b、c、d字母不同代表差异显著(P<0.05)。下同。
Note: LSD multiple comparison, a, b, c and d represent significant differences (P<0.05). The same below.

表 4 香果树树冠生殖构件数量特征
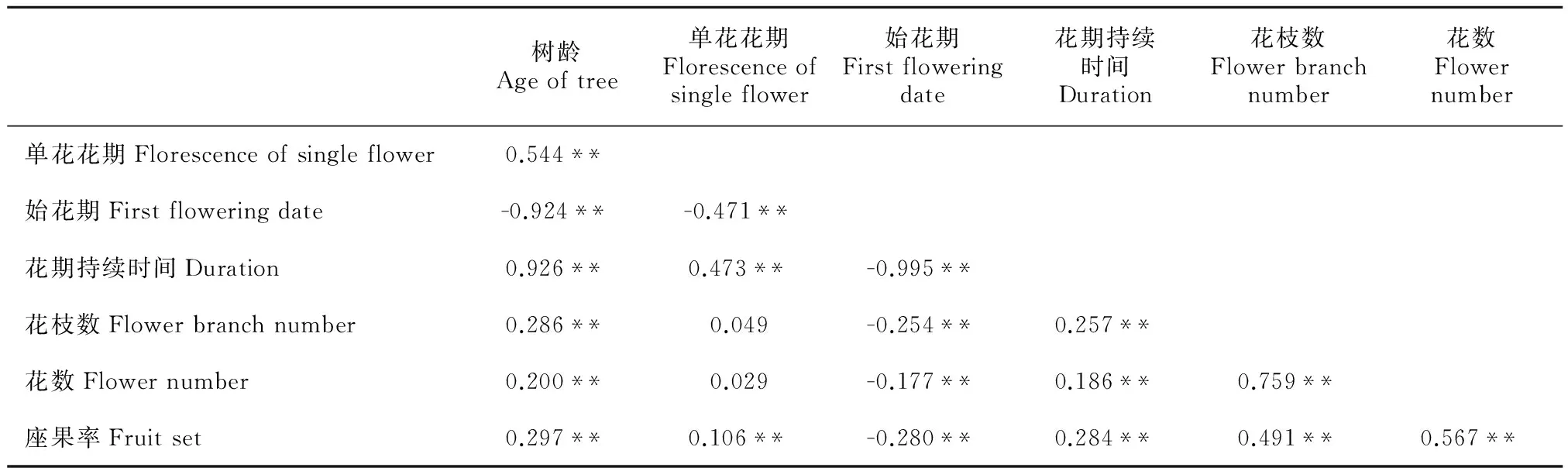
表 5 不同树龄香果树开花物候、繁殖特征各指标间的相关分析
注: **表示相关性极显著; *表示相关性显著。
Note: **Correlation is significant at 0.01 level (2-tailed); *Correlation is significant at 0.05 level (2-tailed).
4.2 生殖构件数量及分布特征
每种植物的生殖构件均有一定的分布格局和数量变化(Lovett, 1980),这种变化与其本身的同化能力以及对体内营养物质的获取和要求有关(边才苗等, 2005)。本研究香果树不同树龄与其生殖构件数量密切相关,香果树个体单花枝产花数随着树龄的增加而呈显著上升趋势,单花枝产果实数呈先升后降趋势,而单株香果树的花枝、花和果实总数均随着其树龄的增加而显著增加。香果树存在花多果少的格局(每73~202朵花形成1个果实),花果转化率在9.13‰~14.07‰之间,转化率极低。树龄较小的香果树与树龄较大的个体在资源竞争中处于劣势,其树高、胸径、枝下高以及冠幅等均小于树龄较大的个体。导致其资源不足,致使其将更多的资源分配给营养器官以提高资源的获取能力(Willson, 1983)。本研究较小树龄的香果树生殖构件之间的转化率较树龄高的植株低,可能是由于其仍然以营养生长为主,其目的是抢占群落的上层空间,克服种内和种间的资源竞争,为转入生殖生长积累必要的有机物等。这与栲树(Castanopsisfargesii)和金花茶等植物类似(柴胜丰等, 2009; 陈波等, 2003)。白桦(Betulaplatyphylla)老龄个体生殖构件的减少是由于生殖枝减少,而不是每个生殖枝上的花和果实数量的减少造成(祖元刚等, 2000)。本研究则发现香果树树龄超过110 a后,其花枝和花数量仍然有所增加,但果实数量在减少,说明香果树可能存在自适应,随着树龄增大,单花枝产花数量最大为86.28 ± 32.36,部分花无法获得足够营养物质,而导致单花枝产果量较少。这一点在树冠不同层位和方向上可体现出来,香果树生殖构件的生长发育与光照有关(郭连金等, 2011)。光照条件较好的树冠上层和南侧花枝和花数量达到最大,但单花枝产果实的数量却在树冠中层和东侧最大,这可能是由于单花枝产花量大,部分花营养不良,无法形成果实所致。
4.3 生殖构件的数量与开花物候的关系
珍稀濒危植物有性生殖过程中一般均存在花多果少格局(刘易鑫, 2011;盛海燕, 2004),而座果率低可能受资源限制或(和)花粉限制(Burd, 1994)。香果树花粉量较大,单花约为256752粒花粉,活力可达97 %(程喜梅, 2008),因此花粉质量不是限制座果率低的原因。本研究由树龄大的香果树个体单花枝数量、花朵数量多,而果实数量少以及光线最好的树冠上层和南侧单花枝结果量较小,这可能是由于大量花朵消耗植株过多营养物质而无法提供果实正常发育所需要的营养物质,导致果实数量较少。对香果树的树龄、物候指标和生殖构件数量进行相关性分析,结果表明香果树树龄除了与始花期呈极显著负相关外,与单花花期、花期持续时间、花枝数、花朵数、座果率均存在极显著正相关;花期持续时间、花枝数、花朵数和果实数之间均存在极显著正相关关系。这表明香果树生殖成功受开花物候的显著影响。这与沙冬青(李新蓉等, 2006)、长柄双花木(肖宜安等, 2005)、准噶尔无叶豆(Eremospartonsongoricum)(马文宝等, 2008)等的研究结果相似。
4.4 濒危植物香果树的保护措施
自然生境中香果树生长20 a后才能首次开花,之后随着其树龄的增加,香果树开花结果的数量逐渐增加,但野外调查发现,在其生境中存在砍伐及破坏香果树的现象,故应严格控制对20 a以上的香果树进行破坏或砍伐。光照显著影响香果树生殖构件的生长发育(郭连金等, 2011)。本研究发现香果树树冠的下层和树冠的北侧着生的花枝、花朵和果实的数量最少,因此,对香果树附近的高大乔木进行疏伐和间伐以改善香果树树冠下层及树冠北侧的光照条件将有利于其生殖构件的生长发育。沙冬青和长柄双花木等濒危植物开花同步性较高,大多为集中开花模式,这有利于吸引传粉者,进而容易实现生殖成功(肖宜安等, 2005; 李新蓉等, 2006),而香果树尽管开花量较大,但访花昆虫种类却较少,且访花频率很低,由其于传粉昆虫以中华蜜蜂、雄蜂以及土蜂为多(程喜梅, 2008)。因此,进行人工养蜂,提高传粉者的数量,将有可能提高香果树的结实率,并有利于其种群的自然更新。
BIAN CM, JIN ZX,LI JM, 2005. Characteristics of reproductive modules of endangeredHeptacodiummiconioides[J]. Acta Bot Boreal-Occident Sin, 25(4): 756-760. [边才苗, 金则新,李钧敏, 2005. 濒危植物七子花的生殖构件特征 [J]. 西北植物学报, 25(4): 756-760.]
BLIONIS GJ, HALLEY JM, VOKOU D, 2001. Flowering phenology of Campanula on Mt Olympos, Greece [J]. Ecography, 24(6): 696-706.
BURD M, 1994. Bateman’s principle and plant reproduction: the role of pollen limitation in fruit and seed set [J]. The Bot Review, 60(1): 83-139.
BUTT N, SEABROOK L, MARON M, et al, 2015. Cascading effects of climate extremes on vertebrate fauna through changes to low-latitude tree flowering and fruiting phenology [J]. Glob Chang Biol, DOI: 10.1111/gcb.12869.
CHAI SF, WEI X, JIANG YS, et al, 2009. The flowering phenology and characteristics of reproductive modules of endangered plantCamellianitidissima[J]. J Trop Subtrop Bot, 17(1): 5-11. [柴胜丰, 韦霄, 蒋运生, 等, 2009. 濒危植物金花茶开花物候和生殖构件特征 [J]. 热带亚热带植物学报, 17(1): 5-11.]
CHEN B, DA LJ, SONG YC. Flowering phenology and floral distribution ofCastanopsisfargesiiin tiantong, Zhejiang Province [J]. Acta Phytoecol Sin, 27(2):249-255. [陈波, 达良俊, 宋永昌. 常绿阔叶树种栲树开花物候动态及花的空间配置 [J]. 植物生态学报, 27(2):249-255.]CHENG XM, 2008. Study on the pollination biology of the state key protected speciesEmmenopteryshenryiOliv. [D]. Zhengzhou: Henan Agricultural University: 1. [程喜梅, 2008. 国家重点保护植物香果树传粉生物学研究 [D]. 郑州: 河南农业大学: 1.]
DU JC, JING DL, CHEN FJ, et al, 2010. 6 Kinds of isozymes after long-term subculture ofEmmenopteryshenryiOliv. [J]. Agric Sci Technol, 11(5):22-27.
EHRLÉN J, 2015. Selection on flowering time in a life-cycle context [J]. Oikos, 124(1): 92-101.
FU LG, JIN JM, 1992. Chinese plant red book-rare and endangered plants (Vol. 1) [M]. Beijing: Science Press: 568-569. [傅立国, 金鉴明, 1992. 中国植物红皮书——稀有濒危植物(第一册) [M]. 北京: 科学出版社: 568-569.]
GEBER MA, GRIFFEN LR, 2003. Inheritance and natural selection on functional traits [J]. Intern J Plant Sci, 164(S3): S21-S42.
GUO LJ, LIN GW, XU WH, et al, 2011. Characteristics of reproductive modules ofEmmenopteryshenryinatural population in Wuyi Mountain [J]. J NW For Univ, 26(4): 18-22. [郭连金, 林国卫, 徐卫红, 等, 2011. 武夷山香果树自然种群生殖构件特性研究 [J]. 西北林学院学报, 26(4): 18-22.]
GUO LJ, 2009. Population structure and dynamics of the endangered plantEmmenopteryshenryi[J]. J Wuhan Bot Res, 27 (5): 509-514. [郭连金, 2009. 濒危植物香果树 (Emmenopteryshenryi) 种群结构与动态 [J]. 武汉植物学研究, 27 (5): 509-514.]HAMMAD I, TIENDERENIENDEREN PH, 1997. Natural variation in flowering time among populations of the annual cruciferArabidopsisthaliana[J]. Plant Species Biol, 12(1): 15-23.HE M, CHEN SH, MA CQ, et al, 2014. Flowering phenology and pollination characteristic ofAdonisamurensis[J]. Pratac Sci, 31(3):431-237. [何淼, 陈士惠, 马翠青, 等, 2014. 野生及引种侧金盏花的开花物候与传粉特性 [J]. 草业科学, 31(3):431-237.]
HENDRY AP, DAY T, 2005. Population structure attributable to reproductive time: isolation by time and adaptation by time [J]. Mol Ecol, 14(4): 901-916.JIAO PP, LI ZJ, 2007. Flowering phenology of an endangered plantAmmopiptanthusnanus[J]. Acta Bot Boreal-Occident Sin, 27(8): 1 683-1 689. [焦培培, 李志军, 2007. 濒危植物矮沙冬青开花物候研究 [J]. 西北植物学报, 27(8): 1 683-1 689.]
KANG HJ, CHEN ZL, ZHOU YH,et al, 2011. Seed germination and dynamics of seedling growth ofEmmenopteryshenryi[J]. J Cent S Univ For Technol, 31(1): 32-37. [康华靖, 陈子林, 周钰鸿,等, 2011. 濒危植物香果树种子萌发及幼苗生长动态的比较 [J]. 中南林业科技大学学报, 31(1): 32-37.]LI JM, JIN ZX, 2008. Genetic structure of endangeredEmmenopteryshenryiOliv. based on ISSR polymorphism and implications for its conservation [J]. Genetica, 133(3): 227-234.
REN CY, ZHANG T, LI RY, 2015. Responses of growing period and phonological duration of woody plants to climate warming in southwestern Shandong Province [J]. J Meteorol Environ, 431(1),: 25-3190-95. [任崇勇,张婷,李瑞英, 2015. 鲁西南草本植物生长季及物候期持续日数对气候变化的响应 [J]. 气候变化研究快报气象与环境学报, 431(1),: 25-3190-95.]
LI TH, ZHOU YX, DUAN XP, et al, 2004. Physiological characteristics of the dormancy and light-sensitive germination ofEmmenopteryshenyiseeds [J]. J Centr S For Univ, 24(2): 82-84. [李铁华, 周佑勋, 段小平, 等, 2004. 香果树种子休眠和萌发的生理特性 [J]. 中南林学院学报, 24(2): 82-84.]
LI XR, TAN DY, GUO J, 2006. Comparison of flowering phenology of two species ofAmmopiptanthus(Fabaceae) underexsituconservation in the Turpan Eremophytes Bo-tanical Garden, Xinjiang [J]. Biodivers Sci, 14(3): 241-249. [李新蓉, 谭敦炎, 郭江, 2006. 迁地保护条件下两种沙冬青的开花物候比较研究 [J]. 生物多样性, 14(3): 241-249.]
LIU YX, 2011. Preliminary study on plant reproductive ecology and bioactive components ofChimonanthusgrammatusM. C. Liu [D]. Nanchang: Jiangxi Normal University: 27-29. [刘易鑫, 2011. 突托蜡梅繁殖生态学与生物活性成分初步研究 [D]. 南昌: 江西师范大学: 27-29.]
LOVETT DJ, 1980. Experimental manipulation of patterns of resource allocation in the growth cycle and reproduction ofSmyrniumolusatrumL. [J]. Biol J Linn Soc, 13: 155-166.
LUO R, GUO JJ, 2010. Plant flowering time: natural variation in the field and its role in determining genetic differentiation [J]. Chin Bull Bot, 5 (1): 109-118. [罗睿, 郭建军, 2010. 植物开花时间: 自然变异与遗传分化 [J]. 植物学报, 5 (1): 109-118.]MA WB, SHI X, ZHANG DY, et al, 2008. Flowering phenology and reproductive features of the rare plantEremospartonsongoricumin
(Continueonpage1111)desertzone, Xinjiang, China [J]. J Plant Ecol, 32(4): 760-767. [马文宝, 施翔, 张道远, 等, 2008. 准噶尔无叶豆的开花物候与生殖特征 [J]. 植物生态学报, 32(4): 760-767.]
(Continuefrompage1100)
MANUEL C, MOLLES J, 2002. Ecology, concept and applications 2nd ed. NewYork: McGraw-Hill Companies: 186-254.
OLLERTON J, LACK A J, 1992. Flowering phenology: an example of relaxation of natural selection? [J]. Trends Ecol Evol, 7(8): 274-276.
SHENG HY, 2004. Ecological strategy of endangered plantChangiumsmyrnioidesWolff [D]. Hangzhou: Zhejiang University: 29-32. [盛海燕, 2004. 濒危植物明党参的生态策略研究 [D]. 杭州: 浙江大学: 29-32.]
WILLSON MF, 1983. Plant reproductive ecology [M]. New York: John Wiley & Sons:1-44.
WU LY, TANG QR, YIN H, et al. Flower bud differentiation mechanism of ornamental plants [J]. Lett Biotechnol, 18(6):1 064-1 067. [吴莉英, 唐前瑞, 尹恒, 等. 观赏植物花芽分化研究进展 [J]. 生物技术通讯, 18(6):1 064-1 067.]XIAO YA, HE P, HU WH, et al. 2005. Temporal and spatial dynamics of reproductive modules of endangered plantDisanthuscercidifoliusvar.longipesnatural populations [J]. Chin J Appl Ecol, 16(7):1 200-1 204. [肖宜安, 何平, 胡文海, 等. 2005. 濒危植物长柄双花木自然种群生殖构件的时空动态 [J]. 应用生态学报, 16(7):1 200-1 204.]
ZHANG DY, 2004. Plant life-history evolution and reproductive ecology [M]. Beijing: Science Press: 43-61, 97-156. [张大勇, 2004. 植物生活史进化与繁殖生态学 [M]. 北京: 科学出版社: 43-61, 97-156.]
ZHANG WB,JIN ZX, 2008. Flowering phenology and pollination success of an endangered plantSinocalycanthusChinensis[J]. Acta Ecol Sin, 28(8): 4 037-4 046. [张文标,金则新, 2008. 濒危植物夏蜡梅(Sinocalycanthuschinensis)的开花物候与传粉成功 [J]. 生态学报, 28(8): 4 037-4 046.]
ZHANG WH, ZU YG, LIU GB, 2002. Population ecological characteristics and analysis on endangered cause of ten endangered plant species [J]. Acta Ecol Sin, 22(9): 1 512-1 520. [张文辉, 祖元刚, 刘国彬, 2002. 十种濒危植物种群生态学特征及致危因素分析 [J]. 生态学报, 22(9): 1 512-1 520.]ZU YG, YUAN XY, 2000. The blooming and production of reproductive modules in relation to tree age and their position within crowns inBetulaplatyphylla[J]. Acta Ecol Sin, 20(4): 673-677. [祖元刚, 袁晓颖, 2000. 白桦的开花时间及生殖构件的数量与树龄和树冠层次的关系 [J]. 生态学报, 20(4): 673-677.]
Flowering phenology and reproductive module characteristics of an endangered plantEmmenopteryshenryiof different age classes
GUO Lian-Jin1*, LI Mei1, LIN Sheng2, XIE Peng2
( 1.ShangraoNormalUniversity, Shangrao 334000, China; 2.AdministrativeBureauofWuyiMountainNationalNatureReserve, Wuyishan 354300, China )
Using fixed sample observation methods, we studied the flowering phenology and reproductive characteristics ofEmmenopteryshenryiof different ages, and analyzed the correlations between phenological indices and reproductive modular were in this paper. The results showed that the florescence ofE.henryisingle flower was 5-9 d, with an average of 6.79 d. With the increasing ages of theE.henryimother trees, they grown exponentially. For theE.henryitrees of different ages, the first-flowering date was quite different, but the flower fading date was almost the same. As the trees getting old, the first-flowering date came earlier, and therefore the florescence was getting longer. The first-flowering date of 20-50 years old trees was August 9th, whose florescence was 36 d, which was the shortest in all theE.henryimother trees different ages. In comparison, the first-flowering date of 110-140 years old trees was July 15th, whose florescence was 61 d. For a singleE.henryimother tree, the number of flowers and flower branches, as well as the total fruits amounts significantly increased with the age of the mother tree increased; the number of flowers showed the same changing trend as above as the mother tree getting old; however the number of fruits per branch increased firstly and then decreased as the age of the mother tree increased; the number of flower branches per tree and number of flowers per branch represented the same changing trend on the canopy, the order of which was south > east or west > north of the canopy, upper > middle > bottom crown, while the fruit number per branch showed different trend, the order of which was east > south > west > north of the canopy, middle > bottom > upper crown. The common phenomenon of theE.henryiwas that flowering number was extremely larger than that of the fruits. The correlation analysis between the flowering phenology indices and the number of reproductive module showed that, the florescence was extremely significantly related with the number of flower branches, total flower and fruit amounts respectively; the first-flowering date was significantly negatively correlated with the number of the productive module, while the phonological indices represent significant correlations with the age ofE.henryitree.
protective plant, reproductive module, flowering phenology, florescence of single flower, reproductive success
10.11931/guihaia.gxzw201505012
2015-05-19
2015-08-11
国家自然科学基金(31360145) [Supported by the National Natural Science Foundation of China (31360145)]。
郭连金(1976- ),男,山东滨州人,硕士,副教授,研究方向为濒危植物保护,(E-mail) guolianjin@163.com。
Q945
A
1000-3142(2016)09-1093-08
郭连金, 李梅, 林盛, 等. 武夷山不同龄级香果树开花物候及其生殖特性 [J]. 广西植物, 2016, 36(9):1093-1100
GUO LJ, LI M, LIN S, et al. Flowering phenology and reproductive module characteristics of an endangered plantEmmenopteryshenryiof different age classes [J]. Guihaia, 2016, 36(9):1093-1100
