七种中国淫羊藿属药用植物细胞分类学研究
2016-10-28严福林何顺志徐文芬黎忠鸿
严福林, 何顺志*, 徐文芬, 黎忠鸿
( 1. 贵阳中医学院 药学院, 贵阳 550002; 2. 贵州省人民医院, 贵阳 550002 )
七种中国淫羊藿属药用植物细胞分类学研究
严福林1, 何顺志1*, 徐文芬1, 黎忠鸿2
( 1. 贵阳中医学院 药学院, 贵阳 550002; 2. 贵州省人民医院, 贵阳 550002 )
该研究应用常规压片法对7种中国淫羊藿属药用植物进行了核型分析,为淫羊藿属药用植物的分类及选育等提供理论基础。结果表明:7种淫羊藿属植物体细胞染色体数目均为2n=2x=12,基数x=6;水城淫羊藿、腺毛淫羊藿、保靖淫羊藿、拟巫山淫羊藿、天全淫羊藿5个种均有4对m染色体和2对sm染色体,核型公式均为2n=2x=12=8m(2SAT)+4sm,其中水城淫羊藿、腺毛淫羊藿的核型分析为首次报道;镇坪淫羊藿、偏斜淫羊藿具有3对m染色体和3对sm染色体,核型公式为2n=2x=12=6m(2SAT)+6sm。拟巫山淫羊藿第2对染色体具随体,其余物种第1对染色体均有随体;腺毛淫羊藿、保靖淫羊藿、拟巫山淫羊藿、镇坪淫羊藿、偏斜淫羊藿、天全淫羊藿均属于Stebbins 2A类型,水城淫羊藿为Stebbins 1A类型。结合文献综合分析发现,该属植物不同种间的核型高度一致,核型稳定,种间核型差异小,核型结构与地理分布、形态特征相关性较小。因此,淫羊藿属是一个在核型进化上较为保守的植物类群,用核型分析对该属植物属下进行种类鉴定、遗传演化十分困难,即使是属内远缘物种,其核型系统学的意义有限,但在属上的分类级别有较大的参考价值。
淫羊藿属, 细胞分类学, 核型分析
淫羊藿属(Epimdium)药用植物来源于小檗科(Beiberdaceae),为多年生草本植物。该属建于1694年,1700年用拉丁文Epimdium命名,Morren & Decaisne于1834年进行首次系统分类(Stearn & William,2002)。淫羊藿具有补肾阳、强筋骨、祛风湿、助阳益精等功效,用于阳痿遗精,筋骨痿软,风湿痹痛,麻木拘挛等症。临床上用于治疗肿瘤、骨质疏松症、类风湿性关节炎,不孕不育、心理性勃起功能障碍和心肌梗死等。淫羊藿的药用最早记载于《神农本草经》,列为中品,是重要的传统中药之一(国家中医药管理局《中华本草》编委会,2005)。近年研究发现(陈国峰等,2001),淫羊藿对于改善心血管系统功能、提高性机能、增强免疫能力、抗骨质疏松和延缓衰老等均表现出生理活性,还具有抑菌抗炎、镇咳祛痰、抗抑郁等作用,深受国内外研究者关注。2010版《中国药典》(国家药典委员会,2010)收载:淫羊藿、箭叶淫羊藿、柔毛淫羊藿、朝鲜淫羊藿和巫山淫羊藿5个种作为中药材使用,其余种类在其产地均作为淫羊藿药材使用。目前,以淫羊藿作主药的中成药有仙灵骨葆列产品、安神补脑液、肾宝归真液、脑灵素片、冠心康片、心病宁、痹康胶囊、抗骨增生颗粒、复方玄驹胶囊等20多个品种,生药用量呈逐年上升趋势。
淫羊藿属植物种类繁多,截至目前,淫羊藿属全球有约59个种,中国分布约有48种(郭宝林和肖培根,1993)。该属植物分布地域性较强,生长环境多样,形态相近。加上近年来一些新种的不断发现,使得该属植物种间关系变得更为复杂,这给该属植物的分类鉴定带来了一定难度。系统进行淫羊藿属植物的核型分析,将有益于该属的系统学研究。
1 材料与方法
1.1 材料
所有实验材料均取自其原产地,由贵阳中医学院药学院何顺志教授鉴定,活体材料种植于贵州省同济堂制药股份有限公司淫羊藿种质资源圃,凭证标本均存于贵阳中医学院标本室(HGCM)。供试材料具体情况详见表1。
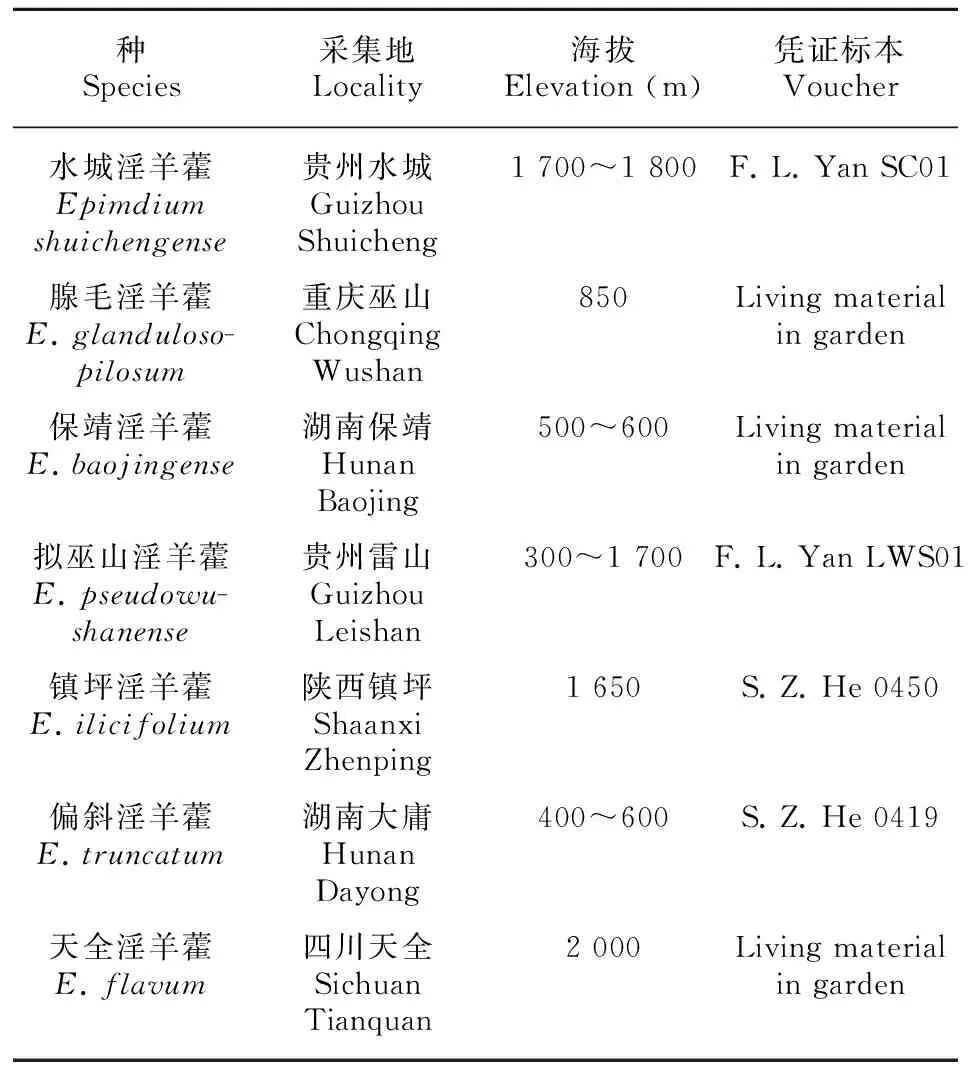
表 1 七种淫羊藿属植物材料来源
1.2 方法
春季取移植后不同植株生长旺盛的幼嫩根尖,清水洗净后置于a-溴钠溶液中预处理5~6 h,转入新鲜配制的卡诺氏固定液(乙醇∶冰醋酸=3∶1)中固定12 h后,用清水清洗,于1 mol·L-1HCl溶液中60 ℃恒温解离10 min,用改良品红染色45 min,常规方法压片。用带有摄像头的奥林巴斯(BX53)显微镜对着丝点清晰、分散良好的中期分裂细胞进行显微拍照,每个类群选取30个以上中期分裂细胞进行染色体计数,并选出5个细胞进行染色体长度测量和计算。永久片用加拿大树胶封存。
核型分析参照李懋学和陈瑞阳(1985)的标准,染色体类型按Levan et al (1964)标准命名,核型不对称程度用不对称系数As.K.表示(As. K.=染色体长臂总长度/全组染色体总长度×100%),百分比越大,核型不对称程度越高,核型类别按Stebbins(1971)的分类标准。
2 结果与分析
对7种淫羊藿植物的根尖有丝分裂中期细胞进行染色体计数和核型分析,结果发现:7种植物均为二倍体,2n=2x=12,基数x=6,第一对染色体都具有随体。
2.1 水城淫羊藿 E. shuichengense
水城淫羊藿是20世纪90年代中期何顺志教授在贵州水城发现的药用植物新种。目前仅在贵州的水城发现有分布,生于海拔1 700~1 800 m的山谷灌丛中。该种主要形态鉴别要点:植株高13~30 cm。叶上面绿色光滑无毛,背面被有白色柔毛。花大,花瓣黄色,短距形,根茎伸长达15 cm以上而不同于小花类群其他种。此种在当地作淫羊藿药用。
核型公式:2n=2x=12=8m(2SAT)+4sm,其中1、2、3、4号4对中部着丝粒染色体(m),占全部染色体的66.67%,且1号具有明显的中间随体;5、6号2对为近中着丝粒染色体(sm),占全部染色体的33.33%。染色体相对长度变异范围在13.18~18.56之间,平均值为16.67。最长染色体/最短染色体=1.41<2,臂比的变异幅度为1.01~1.97,平均臂比值为1.50,核型不对称系数As.K.=58.74%。整个染色体组中,臂比值大于2的染色体百分比为0.00%,因此,水城淫羊藿核型属于Stebbins 1A型。详见表2与表3,图1:A1,B1,C1。
2.2 腺毛淫羊藿 E. glandulosopilosum
腺毛淫羊藿发现于20世纪90年代初,目前仅在重庆市的巫山海拔约850 m发现有分布。主要形态鉴别要点:地下根状茎短粗,呈不规则结节状或瘤状;茎被腺毛和长柔毛,节上尤密。单叶基生和茎生;基生叶阔卵形。总状花序具8~24朵花,花大,黄色,花瓣角状距。该种总状花序,叶柄、花序轴及花梗均被腺毛而不同于大花类群其他种。
核型公式:2n=2x=12=4m(2SAT)+8sm,其中1、6号为中部着丝粒染色体(m),占全部染色体的33.33%,1号具有明显的中间随体;2、3、4、5号为近中着丝粒染色体(sm),占全部染色体的66.67%。染色体相对长度变异范围在11.44~19.72之间。最长染色体/最短染色体=1.72<2。臂比的变异幅度为1.08~2.53,平均臂比为1.84,核型不对称系数As. K.=63.60%。整个染色体组中,臂比值大于2的染色体百分比为50.00%。因此,腺毛淫羊藿核型属于Stebbins 2A型。详见图1:A2,B2,C2和表2,表3。
2.3 保靖淫羊藿 E. baojingense
20世纪80年代初发现的植物新种,目前仅在湖南保靖、大庸、永顺发现有分布,生于海拔500~600 m的林下或沟边。该种叶为一至三回羽状复叶,小叶有小齿;总状花序顶生或与叶对生;内轮的花瓣状,常有颜色;花瓣4,蜜腺状,有距;雄蕊4,离生;蒴果背裂。其性状特征近似恩施淫羊藿,但本种叶背无毛、无白粉而易于区别。
核型公式:2n=2x=12=8m(2SAT)+4sm,其中1、2、3、6号为中部着丝粒染色体(m),占全部染色体的66.67%,且1号染色体短臂上具有明显的中间随体;4、5号为近中着丝粒染色体(sm),占全部染色体的33.33%。染色体相对长度变异范围在10.04~19.2之间。最长染色体/最短染色体=1.92<2。臂比的变异幅度为1.04~1.92,平均臂比值为1.59,核型不对称系数As.K.=59.48%。整个染色体组中,臂比值大于2的染色体百分比为33.33%。因此,保靖淫羊藿核型属于Stebbins的2A型。详见表2,表3和图1:A3,B3,C3。
2.4 拟巫山淫羊藿E. pseudowushanense
该种分布于四川、湖北、广西、贵州、重庆、陕西等地区,生于林下、灌丛中、草丛中或石缝中,海拔300~1 700 m。花大, 具长距, 但不明显长于内轮萼片, 花茎具两叶对生;聚伞状圆锥花序;三出复叶, 小叶披针形或狭披针形。该种近似绿药淫羊藿,但本种叶背被绵毛或无毛,内轮萼片阔椭圆形,花药黄色。
核型公式:2n=2x=12=8m(2SAT)+4sm,其中1、2、3、4号为中部着丝粒染色体(m),占全部染色体的66.67%,且1号具有明显的中间随体;5、6号为近中着丝粒染色体(sm),占全部染色体的33.33%。染色体相对长度变异范围在13.12~20.51之间。最长染色体/最短染色体=1.56<2。臂比的变异幅度为1.04~2.49,平均臂比值为1.57,核型不对称系数As.K.=59.13%。整个染色体组中,臂比值大于2的染色体百分比为16.67%, 因此, 拟巫山淫羊藿核型属于Stebbins的2A型。详见图2:A4,B4,C4和表2,表3。
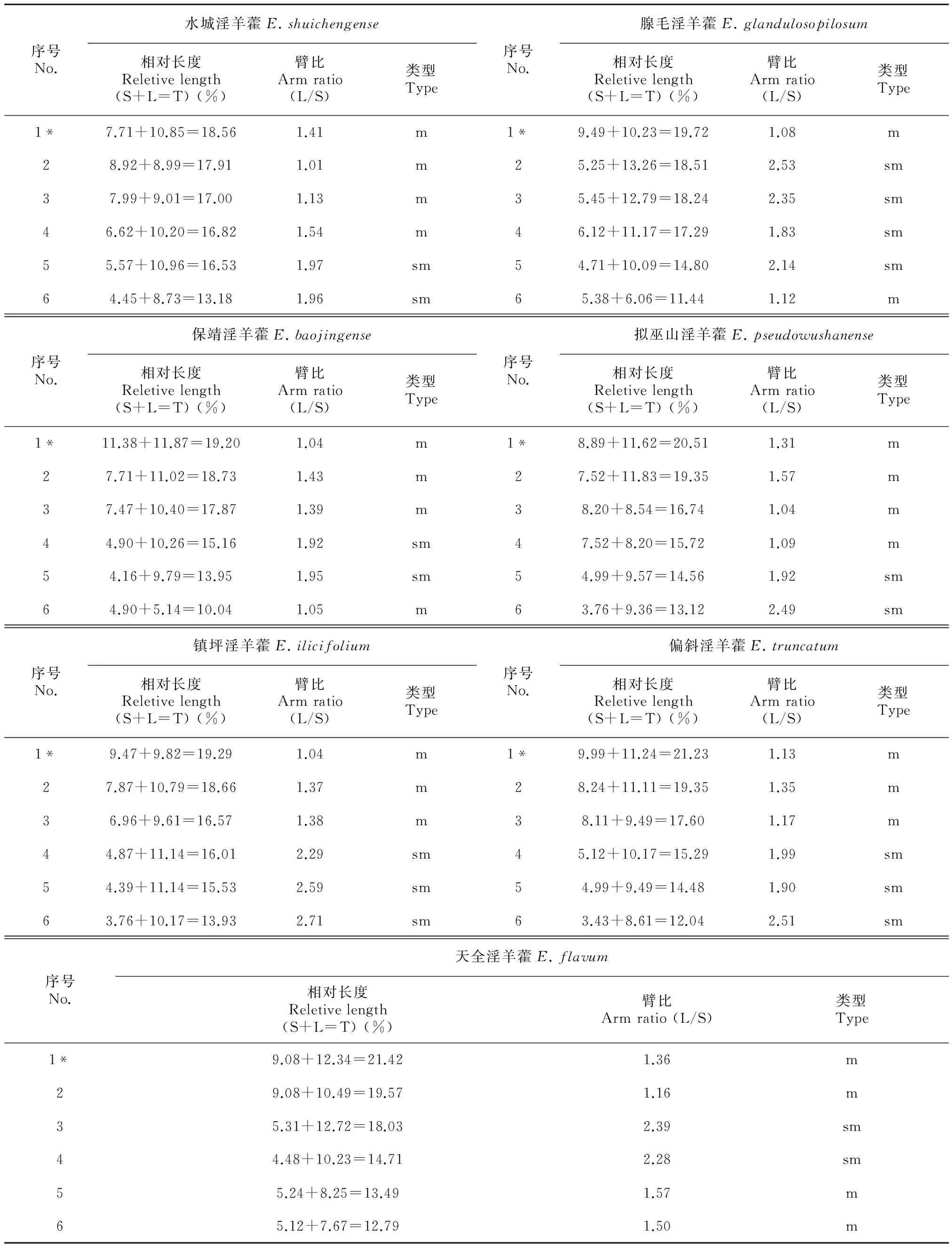
表 2 七种中国淫羊藿属药用植物核型分析参数(*=随体染色体)
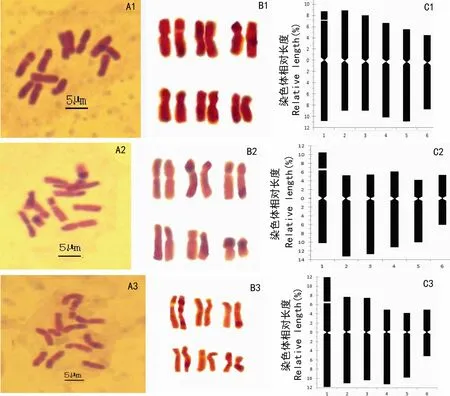
图 1 七种淫羊藿染色体数目(A)、核型(B)及核型模式(C) A1,B1,C1. 水城淫羊藿; A2,B2,C2. 腺毛淫羊藿; A3,B3,C3. 保靖淫羊藿。Fig. 1 Chromosome number (A), karyotype (B) and karyotypes (C) of seven species of Chinese EpimediumA1, B1, C1. E. shuichengense; A2, B2, C2. E. glandulosopilosum; A3, B3, C3. E. baojingense.

图 2 七种淫羊藿染色体数目(A)、核型(B)及核型模式(C) A4,B4,C4. 拟巫山淫羊藿; A5,B5,C5. 镇坪淫羊藿; A6,B6,C6. 偏斜淫羊藿; A7,B7,C7. 天全淫羊藿。Fig. 2 Chromosome number (A), karyotype (B) and karyotypes (C) of seven species of Chinese EpimediumA4, B4, C4. E. pseudowushanense; A5, B5, C5. E. ilicifolium; A6, B6, C6. E. truncatum; A7, B7, C7. E. flavum.
2.5 镇坪淫羊藿 E. ilicifolium
20世纪90年代末期发现的植物新种,仅在分布于陕西的镇坪,海拔约1 650 m。该种近似粗毛淫羊藿,花大而少,具长距, 明显大于内轮萼片,不超过10朵;花茎具两叶对生;多单叶,革质。本种小叶披针形,外轮萼片先端缺裂而易于区别。
核型公式:2n=2x=12=6m(2SAT)+6sm,其中1、2、3号为中部着丝粒染色体(m),占全部染色体的50.00%,且1号具有明显的中间随体;4、5、6号为近中着丝粒染色体(sm)。染色体相对长度变异范围在13.93~19.29之间。最长染色体/最短染色体=1.38<2。臂比的变异幅度为1.04~2.71,平均臂比值为1.90,核型不对称系数As.K.=62.67%。整个染色体组中,臂比值大于2的染色体百分比为50.00%,因此,镇坪淫羊藿核型属于Stebbins 2A型。详见图2:A5,B5,C5和表2,表3。
2.6 偏斜淫羊藿 E. truncatum
20世纪90年代初发现的药用植物新种,目前仅在湖南的保靖、大庸发现有分布,生于海拔400~600 m的山坡灌丛中;该种小叶基部平截,不呈心形而不同于小花类群其他种。
核型公式:2n=2x=12=6m(2SAT)+6sm,其中1、2、3号为中部着丝粒染色体(m),占全部染色体的50.00%,且1号具有明显的中间随体;4、5、6号为近中着丝粒染色体(sm)。染色体相对长度变异范围在12.04~21.23之间。最长染色体/最短染色体=1.76<2。臂比的变异幅度为1.13~2.51,平均臂比值为1.68,核型不对称系数As.K.=60.11%。整个染色体组中,臂比值大于2的染色体百分比为16.67%,因此,偏斜淫羊藿核型属于Stebbins 2A型。详见图2:A6,B6,C6和表2,表3。
2.7 天全淫羊藿 E. flavum
分布于四川天全二郎山,生于海拔2 000 m。本种极近宝兴淫羊藿,但本种总状花序;内轮萼片长约11 mm;花淡黄色,易于区别。
核型公式:2n=2x=12=8m(2SAT)+4sm,其中1、2、5、6号为中部着丝粒染色体(m),占全部染色体的66.67%,且1号具有明显的中间随体;3、4号为近中着丝粒染色体(sm)。染色体相对长度变异范围在12.79~21.42之间。最长染色体/最短染色体=1.67<2。臂比的变异幅度为1.16~2.39,平均臂比值为1.71,核型不对称系数As.K.=61.70%。整个染色体组中,臂比值大于2的染色体百分比为33.33%,因此,天全淫羊藿核型属于Stebbins的2A型。详见图2:A7,B7,C7和表2,表3。
3 讨论
全球淫羊藿属的染色体核型及带型均有一定数量的文献报道,但报道结果存在不统一现象(盛茂银和陈庆富,2007a,b,2010;Kuroki, 1967,1970;张燕君等,2008)。目前尚未有全面系统的研究报道。本研究对淫羊藿属药用植物7个种的核型进行了分析,E.shuichengense、E.glandulosopilosum核型分析为首次报道。7个淫羊藿种类根尖细胞染色体数目均为2n=2x=12,基数x=6;除E.glandulosopilosum第二对染色体具有中间随体外,其余六种第1对染色体均有中间随体;染色体类型均为m、sm两种,核型不对称系数变异幅度为58.74%~63.60%,较为集中,变异幅度为1.38~1.76,因此,淫羊藿属染色体核型较为对称。水城淫羊藿、腺毛淫羊藿、保靖淫羊藿、拟巫山淫羊藿、天全淫羊藿5个种均有4对m染色体和2对sm染色体,核型公式均为2n=2x=12=8m(2SAT)+4sm;镇坪淫羊藿、偏斜淫羊藿具有3对m染色体和3对sm染色体,核型公式为2n=2x=12=6m(2SAT)+6sm。腺毛淫羊藿、保靖淫羊藿、拟巫山淫羊藿、镇坪淫羊藿、偏斜淫羊藿、天全淫羊藿均属于Stebbins 2A类型,水城淫羊藿为Stebbins 1A类型。这与Kuroki(1967,1970)、盛茂银和陈庆富(2007a,b,2010)、张艳君等(2008)的研究报道结果一致。本研究中,经过反复验证发现淫羊藿属植物水城淫羊藿的核型为1A型。但多数淫羊藿属植物核型集中表现为2A型,由此得出淫羊藿属是一个在核型上进化上较为保守的植物类群。
据小檗科山荷叶(Diphylleia)、八角莲(Dysos~ma)、足叶草(Podophyllum)、桃儿七(Sinopodo~phyllum)4个属植物的核型资料报道,(Li,1986;马绍宾和胡志浩,1996; Koga et al,1991)核型公式分别为2n=2x=8m(SAT)+2st+2t、2n=2x=8m(2SAT)+2st(SAT)+2t、2n=2x=6m(4SAT)+2sm+2st+2t和2n=2x=8m(2SAT)+2st(2SAT)+2t,与淫羊藿属植物的核型有较大差异,反映出淫羊藿属与其他属间的亲缘关系较远,与孢粉学、分子标记、同工酶及化学分类学等研究结果一致(郭宝林,1999;郭宝林等,2008;盛茂银和陈庆富,2007;何顺志和郭宝林,1997)。
综合分析发现,盛茂银等(2010)对18种西南地区产淫羊藿属植物进行了核型分析,除E.yingjiangense(印江淫羊藿) 2n=4x=24外,其余种类均为2n=2x=12。淫羊藿属植物核型有1A, 2A级 3A类型,但主要为2A型。Kuroki(1967,1970)对日本产的所有淫羊藿属种类进行核型分析发现,所研究的种类均为2n=2x=12,2A型。 张燕君等(2008)对中国产21种淫羊藿属植物进行核型分析发现,所研究的种类均为2n=2x=12,2A型。这说明淫羊藿属植物不同种间的核型高度一致,核型稳定,种间核型差异小。因此,应用核型分析对淫羊藿属植物属下进行种类鉴定、遗传演化十分困难,即使是属内远缘物种,其核型系统学的意义有限,但在属上的分类级别有较大参考价值。
CHEN GF,LIU F, 2001. Advance in study onEpimedium[J]. J Shaanxi Trad Chin Med,22(10):624-625. [陈国峰,刘峰, 2001. 淫羊藿属研究进展 [J]. 陕西中医,22(10):624-625.]
CHINA’S STATE PHARMACOPOEIA COMMITTEE, 2010. The first part of Chinese pharmacopoeia. 2010 ed: Vol. 1 [S]. Beijing:Chinese Medical Science and Technology Press: 306-308. [国家药典委员会, 2010. 中国药典2010版一部 [S]. 北京:中国医药科技出版社: 306-308.]GUO BL,XIAO PG, 1993. ChineseEpimediumspecies of medicinal plants belonging to the geographical distribution of research [J]. Chin Trad Herb Drugs,16: 14-17. [郭宝林,肖培根, 1993. 中国淫羊藿属植物种类与分布研究 [J]. 中药材,16: 14-17.]
GUO BL, 1999. Molecular systematic research onEpimediumL. [D]. Beijing: China Union Medical University. [郭宝林, 1999. 淫羊藿属植物的分子系统学研究 [D]. 北京: 中国协和医科大学.]
GUO BL,PEI LK,XIAO PG, 2008. Study on significance of flavonoides in classification ofEpimedium[J]. J Syst Evol,6:874-885. [郭宝林,裴利宽,肖培根, 2008. 淫羊藿属植物黄酮类化合物的分类学意义 [J]. 植物分类学报,6:874-885.]
HE SZ,GUO BL, 1997.Epimediumgenera of guizhou medicinal plants pollen morphology of scanning electron microscope [J].
(Continueonpage1022)Chin J Chin Mat Med,5:12-14. [何顺志,郭宝林, 1997. 贵州产淫羊藿属药用植物花粉形态的扫描电镜观察 [J]. 中国中药杂志,5:12-14.]
(Continuefrompage1045)
KOGA S,SHOYAMA Y,NISHIOK A, 1991. Studies onEpimediumspecies:flavonol glycosides and isozymes [J]. Biochem Syst Ecol,19(4):315-318.KUROKI Y, 1970. Chromosome study in four species ofBerberdaceae[J]. Memoir Ehime Univ Sect,Series B,6(4):215-221.KUROKI Y, 1967. Chromosome study in seven species ofBerberdaceae[ J]. Memoir Ehime Univ Sect,Se-ries B,5(3):175-181.LEVAN A,FREDGE K,SANGCERG AA, 1964. Normenclature for centromeric position on chromosomes [J]. Hereditas,52:201-220.
LI MX,CHEN RY, 1985. The problem on karyotype analysis standardization [J]. J Wuhan Bot Res,3(4):297-302. [李懋学,陈瑞阳, 1985. 核型分析标准化的问题 [J]. 武汉植物学报,3(4):297-302.]
LI LC, 1986. Study on the karyotypes and evolution ofDysosmapleiantha with its relatives [J]. Acta Bot Yunnan,8(4):451-457. [李林初, 1986. 八角莲及其近缘的核型和演化的研究 [J]. 云南植物研究,8(4):451-457.]
MA SB, HU ZH, 1996. Akaryotypic study onpo-dophylloideae(Berberidaceae) [J]. Acta Bot Yunnan,18(3):325-330. [马绍宾, 胡志浩, 1996. 小檗科鬼臼亚科植物的核型研究 [J]. 云南植物研究,18(3):325-330.]SHENG MY,WANG LJ,TIAN XJ, 2010. Karyomorphology of eighteen species of genusEpimedium(Berberidaceae) and its phylogenetic implications [J]. Genet Res Crop Evol,57(8):1 165. [盛茂银,王林娇,田兴军, 2010. 18种淫羊藿属(小檗科)核型及其系统发育的影响 [J]. 植物遗传和资源学报,57(8):1 165.]
STEARN, WILLIAM T, 2002. The genusEpimediumand other herbaceous Berberidaceae [M]. Portland:Timber Press.
STEBBINS GI, 1971. Chromosome evolution in higher plants [M]. London:Edward Anronold Ltd: 87-123.
SHENG MY,CHEN QF, 2007a. Karyotype analysis of sixEpimediumspecies native to Guizhou,China [J]. Guihaia,29:440-443. [盛茂银,陈庆富, 2007a. 六种贵州淫羊藿属植物核型分析 [J]. 广西植物,29:440-443.]
SHENG MY, CHEN QF, 2007b. The POD isozyme analysis of sixEpimediumspecies native to Guizhou,China [J]. J Guizhou Norm Univ:Nat Sci Ed,1:5-8. [盛茂银, 陈庆富, 2007b. 6个黔产淫羊藿属植物的过氧化物酶同工酶分析 [J]. 贵州师范大学学报·自然科学版,1:5-8.]
SHENG MY,CHEN QF, 2010. Karyomorphology of twelve species inEpimedium(Berberidaceae) [J]. Acta Bot Yunnan,29:309-315. [盛茂银,陈庆富, 2010. 12种淫羊藿属植物核型分析 [J]. 云南植物研究,29:309-315.]
THE STATE ADMINISTRATION OF TRADITIONAL CHINESE MEDICINE “CHINESE MATERIA MEDICA: EDITORIAL BOARD, 2005. Materia medica. Miao medicine volume [M]. Beijing:Science Press. [国家中医药管理局《中华本草》编委会, 2005. 中华本草·苗药卷 [M]. 北京:科学出版社.]ZHANG KT,WANG PL, 1983. Study on the pollen morphology of the familyBerberidaceae[J]. Acta Phytotax Sin,21(2):130-141.
ZHANG YJ,HAI SD,AI PM,et al, 2008. Karyomorphology ofEpimedium(Berberidaceae)and its phylo-genetic implications [J]. Caryologia: 613.
Cytotaxonomy of seven species ofEpimediummedicinal plants from Chinese
YAN Fu-Lin1, HE Shun-Zhi1*, XU Wen-Fen1, LI Zhong-Hong2
( 1.DepartmentofPharmacy,GuiyangCollegeofTraditionalChineseMedicine, Guiyang 550002, China;2.People’sHospitalofGuizhouProvince, Guiyang 550002, China )
In order to provide theoretical basis classification and seed breeding research for the medicinal plants ofEpimedium, we studied seven species of medicinal plants of ChineseEpimediumkaryotype, using the conventional pressed discmethod, on the basis of related and summary of phase information consult of karyotype analysis. The results showed that the somatic chromosome numbers were 2n=2x=12, the cardinal numbes were x=6. TheE.shuichengense,E.glandulosopilosum,E.baojingense,E.wushanenseandE.flavumhad four pairs of m chromosome and two pairs of sm chromosomes, and karyotype for mul as were 2n=2x=12=8m(2SAT)+4sm, among them, analysis onE.shuichengenseandE.glandulosopilosumkaryotype was reported for the first time;E.ilicifolium,E.Truncatum2n=2x=12=6m(2SAT)+6sm. The second pair of chromosomes ofE.glandulosopilosumwere with the middle satellite, and the first pair chromosome of other species were with the middle satellite.E.glandulosopilosum,E.baojingense,E.wushanense,E.ilicifolium,E.TruncatumandE.flavumbelonged to “2A” of Stebbins,E.shuichengensebelonged to “1A” of Stebbins. The results showed that the differentiation degree ofEpimediumkaryotype was lower, the correlation between geographical distribution, morphological characteristics and karyotype were smaller, and karyotype structure and geographical distribution, morphological characteristics correlation was smaller. Therefore,Epimediumwas a more conservative in karyotype evolution of taxa. It was very difficult to identify the barrenwort subordinate species and genetic evolution using the karyotype analysis, even within distant species, its karyotype systematics had limited significance, but in the classification level of genera, karyotype analysis still had larger reference value.
Epimedium, cytotaxonomy, karyotype
10.11931/guihaia.gxzw201501029
2015-01-22
2015-04-22
国家自然科学基金(81160506);贵州省中药现代化项目[黔科合社字(2010)5050号][Supported by the National Natural Science Foundation of China(81160506); Guizhou Modernization of Traditional Chinese Medicine(2010)5050]。
严福林(1986-),男,贵州贵阳人,硕士,助教,主要从事中药及民族药资源分类鉴定、栽培等研究,(E-mail)362048796@qq.com。*
何顺志,教授,研究生导师,主要从事中药及民族药资源、分类鉴定与质量控制等研究,(E-mail)hesz8899@126.com。
Q942
A
1000-3142(2016)09-1039-07
严福林, 何顺志, 徐文芬, 等. 七种中国淫羊藿属药用植物细胞分类学研究[J]. 广西植物, 2016, 36(9):1039-1045
YAN FL, HE SZ, XU WF, et al. Cytotaxonomy of seven species ofEpimediummedicinal plants from Chinese[J]. Guihaia, 2016, 36(9):1039-1045
