21种野生百合亲缘关系的ISSR分析
2016-10-28罗建让肖菲菲张延龙牛立新
侯 珺, 罗建让, 肖菲菲, 张延龙, 牛立新
( 西北农林科技大学 风景园林艺术学院, 陕西 杨凌 712100 )
21种野生百合亲缘关系的ISSR分析
侯珺, 罗建让, 肖菲菲, 张延龙*, 牛立新
( 西北农林科技大学 风景园林艺术学院, 陕西 杨凌 712100 )
该研究选用7条ISSR标记引物对百合属21种野生百合的亲缘关系进行分析,使用POPGEN1.32和MEGA5.1数据处理软件分析数据。结果表明:7条引物扩增出149条条带,其中136条为多态性条带,多态性条带比率为91.06%,平均有效等位点数为1.762 4,平均Nei’s基因多样度为0.421 4,平均Shanon信息指数0.608 5。遗传距离变化范围为0.307 5~0.887 3,紫脊百合和尖被百合的遗传距离最大,均值为0.887 3,表明21份材料中二者亲缘关系最远;紫脊百合和宜昌百合遗传距离最小,均值为0.307 5,表明二者亲缘关系最近。聚类结果与形态学分类大致吻合,21个供试材料可被分成5个类群,大理百合、宝兴百合、卷丹百合、紫斑百合、乳头百合、川百合、兰州百合、山丹百合、绿花百合和毛百合为第Ⅰ类群;紫脊百合、宜昌百合、岷江百合、通江百合和淡黄花百合为第Ⅱ类群;野百合和百合为第Ⅲ类群;青岛百合为第Ⅳ类群;第Ⅴ类群包括玫红百合、有斑百合和尖被百合。毛百合与卷瓣组百合亲缘关系较近,钟花组百合与卷瓣组百合存在基因交流,说明不能仅仅依靠形态学对野生百合进行分类。聚类分析结果中野百合与百合单独聚为一类,这说明是否具叶柄对野生百合的分类是一个重要的形态学特性。ISSR分子标记适合用于百合属植物的亲缘关系分析。
百合属, 野生百合, ISSR, 亲缘关系, 树状图
百合属(Lilium)为百合科百合族的一个多年生属,有90余种,大多分布在北半球的温带和寒带地区,少数分布在热带高海拔山区,中国为世界百合起源中心,约46种18个变种(赵祥云等,1999),分布于27个省市自治区(陈心启等,1980)。百合属植物具多年生地下鳞茎,花色多样,药用及观赏价值极高。虽然野生百合种质资源丰富,但对百合属的属下类群划分、起源、进化等问题仍争议颇多。现国际最常采用的分类系统是Comber(1949)依据15个形态特征建立的,他将百合属分为7组。《中国植物志》将百合属植物划分为4个组(陈心启等,1980)。Nishikawa et al(1999)最早对百合属进行分子系统学研究,发现国际分类系统的毛百合组与卷瓣组亲缘关系较近,国际分类系统中属于毛百合组的毛百合被划分在中国分类系统中的钟花组,由此可见毛百合的系统位置较复杂。我国百合属分组中属于百合组的野百合和百合,在Comber的分组中被划分在具叶柄组(sect.Archelirion)。中国是百合属植物的自然分布中心之一,因此研究中国百合属植物亲缘关系对于探讨百合属起源、进化、迁移以及建立科学的分类系统至关重要。目前国内百合系统研究多以形态学(向地英等,2006)、染色体特征(张艺萍等,2014)、花粉形态(顾欣等,2013)为依据。利用分子标记研究百合亲缘关系多见于栽培品种,原生种的分子标记有RAPD,AFLP,SRAP分析,较全面的针对野生种系统学的研究较少。
简单重复序列间扩散多态性(ISSR)是由加拿大蒙特利尔大学Ziekiewicz et al(1994)基于SSR标记创建来的一种新型分子标记技术,其基本原理就是在SSR的3′或5′端加锚1~4个随机核苷酸作为引物,对两侧具有反向排列SSR的一段DNA序列进行扩增,然后进行电泳、染色,根据所读取谱带的有无及相对位置差异,来分析不同样品间 ISSR标记的多态性。由于 ISSR标记技术结合了RAPD和SSR技术的优点,实验操作简便快捷、重复性高、稳定性好、成本较低、所揭示的遗传多态性十分丰富,因此,ISSR技术已被广泛应用于植物育种、植物遗传多样性、系统发育、品种鉴定、基因定位、遗传作图等研究中(Tsumura et al,1996;Qian et al,2011;Pradeep et al,2002;Devarumath et al,2002;Guo et al,2011)。 本研究将此项技术应用于中国百合属的亲缘关系分析,首次对我国百合属四个组21种野生类群进行研究,希望为百合属系统学研究及资源合理利用提供科学依据,并对分类上存在争议的种的归属进行确定。
1 材料与方法
1.1 材料
供试的野生百合材料有21个种或变种,来自陕西、甘肃、云南、四川和山东省(表1),由西北农林科技大学牛立新教授鉴定,种植于西北农林科技大学百合资源温室内。
1.2 方法
1.2.1 DNA提取采用Biospin植物基因组DNA提取试剂盒提取21份植物材料的基因组DNA。用1%的琼脂糖凝胶电泳检测DNA的纯度与浓度,稀释至80 ng·μL-1于-20 ℃保存。

表 1 供试21种野生百合采集地
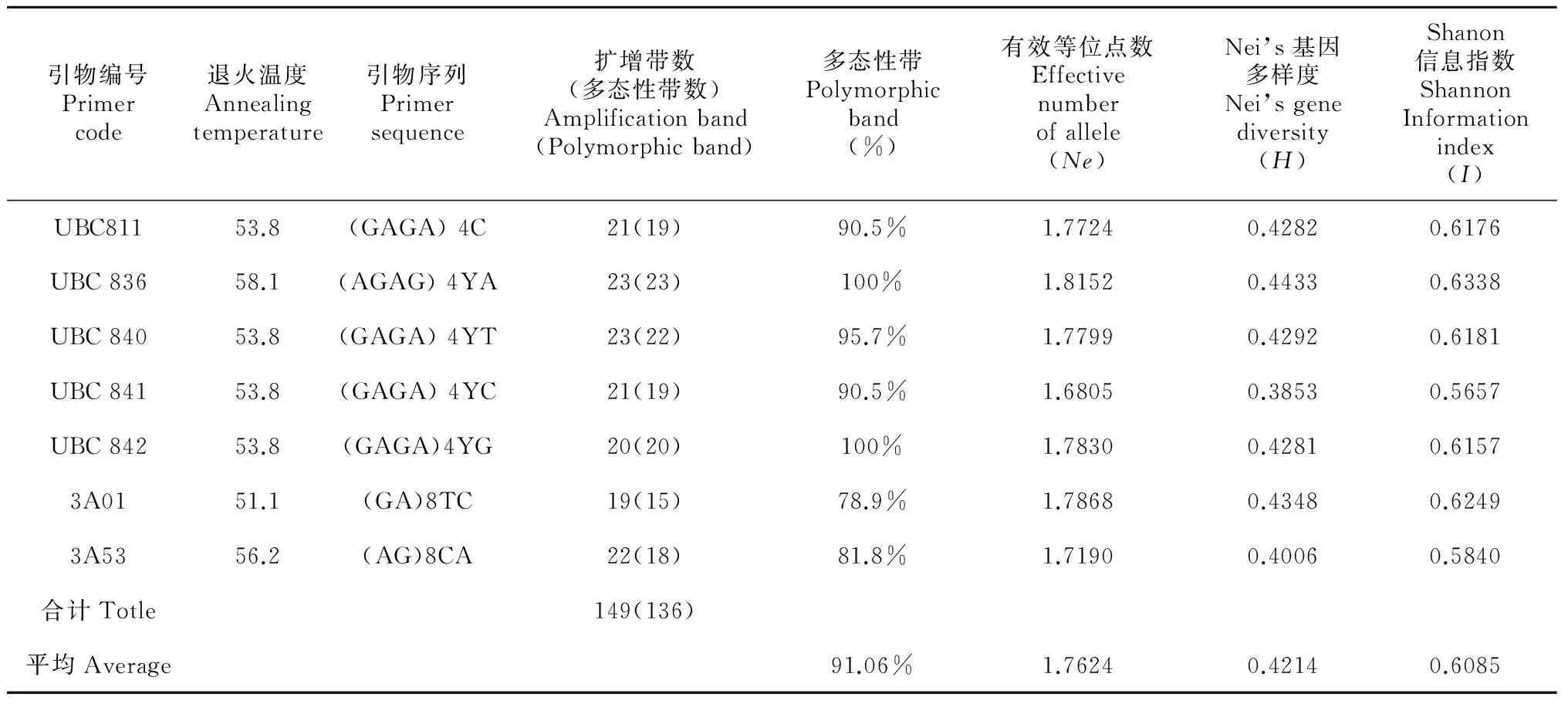
表 2 7条ISSR引物的退火温度及其扩增结果
Y = (C, T)
1.2.2 引物筛选及优化体系建立基于早期ISSR标记在木兰亚属植物应用中的研究报道,本研究引物参照加拿大哥伦比亚大学(UBC)公布的序列和日本Yamagishi et al(2002)在研究百合所采用的3A引物序列,由上海生工生物工程技术服务有限公司合成,利用Eppendorf梯度PCR扩增仪(Mastercycler pro型)确定引物最佳退火温度和筛选引物,最终从163条引物中筛选出7条稳定且重复性好的ISSR引物。利用单因素设计,对影响PCR反应的模板浓度、引物浓度、循环次数等进行优化,建立适合野生百合ISSR-PCR最佳反应体系。
PCR总反应体系为20 μL,含10 μL 2 ×TaqPCR Master Mix,RNase-free water 8 μL ,DNA模板80 ng,10 μmol·L-1引物1 μL。PCR扩增反应程序为94 ℃预变性5 min,94 ℃变性40 s,51.1~58.1 ℃(因引物不同而异)退火45 s,72 ℃ 延伸1.5 min,39个循环,最后72 ℃延伸8 min,4 ℃ 保存。PCR产物在6%聚丙烯酰胺凝胶上电泳,银染检测。
1.2.3 数据统计分析根据扩增产物电泳图谱,对清晰且易于辨认的条带,在相同迁移位置上有带标为“1”,无带标为“0”,构建出样品分子标记0、1矩阵。应用POPGEN 1.32软件,采用SM相似系数法计算21份野生百合材料的多态性条带百分比(PPB)、有效等位基因树(Ne)、Nei’s基因多样度(H)、Shanon信息指数(I)、和遗传距离。应用MEGA5.1软件,采用邻接法(Neighbor-joining)进行聚类分析,得出野生百合亲缘关系聚类图。
2 结果与分析
2.1 ISSR扩增结果
选用野生百合四个组的四种野生百合材料,分别为轮叶组的青岛百合、钟花组的玫红百合、卷瓣组的卷丹百合和百合组的宜昌百合,筛选ISSR引物,从169条引物中筛选出7条扩增效果较好的引物,用于野生百合材料的ISSR扩增。多态性条带的分子量主要分布在250~750 bp之间,扩增结果及多态性信息见表2,共得到清晰条带149条,多态性条带136条,多态性条带比率为91.06%。经重复试验发现 ISSR标记稳定性较强,多态性条带丰富,利用7条引物能有效地将所有样品进行区分,因而ISSR可用于野生百合种质的鉴定。其中引物UBC842对 21 份百合材料的扩增结果见图 1 。
2.2 亲缘关系分析
21份供试材料的遗传距离变化范围为0.307 5~0.887 3。ISSR引物组合平均有效等位基因数、Nei’s 基因多样度和 Shannon 信息指数均值分别为1.762 4,0.421 4和0.608 5(表2)。表2表明供试材料多态性高,遗传变异丰富。数据统计结果显示,紫脊百合和尖被百合的遗传距离最大,均值为0.8873,表明21份野生百合材料中二者亲缘关系最远;紫脊百合和宜昌百合遗传距离最小,均值为0.3075,表明二者亲缘关系最近(表3)。

图 1 引物UBC842对21个野生百合样品扩增的凝胶电泳图谱 1. 有斑百合; 2. 尖被百合; 3. 野百合; 4. 山丹百合; 5. 绿花百合; 6. 通江百合; 7. 川百合; 8. 淡黄花百合; 9. 紫脊百合; 10. 宜昌百合; 11. 岷江百合; 12. 百合; 13. 玫红百合; 14. 兰州百合; 15. 青岛百合; 16. 乳头百合; 17. 大理百合; 18. 紫斑百合; 19. 毛百合; 20. 宝兴百合; 21. 卷丹百合。Fig. 1 Electrophoresis result of amplification of 21 wild species of Lilium using primer U842 1. L. concolor var. pulchellum; 2. L. lophophorum; 3. L. brownie; 4. L. pumilum; 5. L. fargesii; 6. L. sargentiae; 7. L. davidii; 8. L. sulphureum; 9. L. leucanthum var. centifolium; 10. L. leucanthum; 11. L. regale; 12. L. brownii var. viridulum; 13. L. amoenum; 14. L. davidii var. unicolor; 15. L. tsingtauense; 16. L. papilliferum; 17. L. taliense; 18. L. nepalense; 19. L. dauricum; 20. L. duchartrei; 21. L. lancifolium.

2102000.53061900.49380.40181800.44670.58190.55591700.43530.44670.48180.48181600.49380.54320.55590.62220.56881500.72300.65000.59520.66420.65000.62221400.62220.49380.50590.43530.51820.55590.50591300.51820.58190.69310.65000.65000.58190.62220.67851200.49380.43530.62220.59520.60860.55590.59520.60860.63601100.67850.50590.47000.58190.58190.56880.54320.53060.54320.54321000.43530.59520.63600.59520.63600.66420.56880.56880.60860.51820.4938900.30750.45830.59520.75380.42400.63600.55590.49380.47000.58190.59520.5952800.51820.59520.40180.63600.73830.58190.62220.59520.63600.58190.70800.63600.6360700.56880.48180.72300.48180.49380.58190.38010.58190.50590.47000.54320.50590.67850.5688600.60860.44670.55590.60860.53060.59520.63600.56880.66420.69310.62220.42400.63600.54320.6785500.54320.54320.50590.56880.65000.56880.60860.59520.50590.54320.51820.55590.50590.54320.58190.5559400.42400.60860.60860.54320.58190.63600.53060.59520.66420.56880.75380.55590.54320.51820.48180.70800.4938300.55590.54320.55590.72300.56880.53060.53060.55590.47000.58190.56880.66420.69310.62220.59520.58190.51820.6500200.78550.75380.65000.72300.78550.65000.88730.66420.63600.65000.58190.62220.72300.63600.80180.80180.60860.67850.6500100.41280.81830.75380.62220.75380.58190.59520.75380.63600.60860.70800.53060.62220.69310.63600.67850.80180.66420.70800.8018123456789101112131415161718192021
注: 表内野生百合顺序同图1。
Note: see Figure 1 for the sequence ofLilium.

图 2 21种野生百合聚类图Fig. 2 Cluster analysis of 21 wild species of Lilium based on ISSR data using UPGMA
2.3 聚类分析
根据ISSR扩增结果,得到21份野生百合材料的亲缘关系树状图(图2)。由图2可见, 21个供试材料可分成5类:Ⅰ类包括卷瓣组的大理百合、宝兴百合、卷丹百合、紫斑百合、乳头百合、川百合、兰州百合、山丹百合和绿花百合,钟花组的毛百合;Ⅱ类包括百合组的紫脊百合、宜昌百合、岷江百合、通江百合和淡黄花百合;Ⅲ类包括百合组的野百合、百合;Ⅳ类有轮叶组的青岛百合;Ⅴ类有钟花组的玫红百合、有斑百合和尖被百合。
3 讨论与结论
本研究采用7个ISSR标记引物对原产中国的21个百合野生种基因组DNA进行扩增,得到清晰条带149条,多态性条带136条,多态性比率为91.06%。统计结果显示供试材料间的遗传距离较大,这说明我国野生百合种间差异明显、基因资源丰富。由此可见,ISSR标记适合于百合属植物亲缘关系分析,是一种有效、可靠的分子标记。
21份材料的遗传距离变化范围为0.307 5~0.887 3。数据统计结果显示,紫脊百合和尖被百合的遗传距离最大,均值为0.887 3,说明二者亲缘关系最远;紫脊百合为宜昌百合的变种,紫脊百合和宜昌百合遗传距离最小,均值为0.307 5,说明二者亲缘关系最近。兰州百合为川百合变种,二者遗传距离为0.380 1,百合为野百合变种,二者遗传距离为0.47,遗传距离均较小,种间遗传距离多大于0.47,可见变种与原种亲缘关系比种间亲缘关系近。
通过对21种野生百合的聚类分析,可以将供试材料分成5类。其中卷瓣组全部聚为一类,百合组大多数种聚为一类,轮叶组自成一类,钟花组大多数聚为一类,聚类结果大致上与形态学分类一致。聚类结果中第Ⅰ类中,大多数野生种花被片反卷程度大,与其他类形态学上差异明显;第Ⅱ类中花呈喇叭状,花被先端向外反卷,程度小于卷瓣组反卷程度;第Ⅲ类与第Ⅱ类花的性状一致,区别在于叶子具叶柄;第Ⅳ类为轮叶组的青岛百合,其叶片轮生,与其他类群叶片着生方式大不相同;第Ⅴ类野生百合花呈钟状,花瓣先端微弯。由此可见,花被片的反卷以及叶子的性状都是野生百合分类重要的依据。
传统分类中,毛百合被划分在钟花组,本研究中毛百合与卷瓣组所有种聚为一类。根据国际野生百合形态学分组,属于中国野生百合钟花组(sect.Lophophorum)的毛百合被放入一个中国分组中没有的毛百合组(sect.Daurolirion)中,在Michael & Harris(2002)的百合进化树图中,钟花组的毛百合与卷瓣组的卷丹、山丹划在一个进化树分支上,Lee(2011)通过DNA测序研究朝鲜百合属植物进化分支,聚类结果第6分支将毛百合组与大多数的卷瓣组百合聚为一类,说明毛百合组与卷瓣组百合之间亲缘关系较近。这与 Dubouzet & Shinoda(1999)研究结果相同。智利等(2011)在对中国23个野生种通过SRAP分子标记对其进行遗传多样性研究表明,毛百合与卷瓣组聚为一类。由此可见,钟花组的毛百合与卷瓣组聚为一类,是符合野生百合形态学和分子水平分类的,证实了Nishikawa et al(1999)的发现,国际分类系统的毛百合组与卷瓣组亲缘关系较近,钟花组与卷瓣组之间存在基因交流。毛百合虽未能与同样花冠的种聚在一起,而与花被反卷的卷瓣组聚在一起,说明毛百合系统位置可能比较复杂,有待随后继续研究。
在中国野生百合分组中,野百合和百合属于百合组,而在Comber(1949)的野生百合分组中,野百合和百合被放在具叶柄组(sect.Archelirion)中,中国分组中无具叶柄组。本研究聚类结果显示,野百合与百合单独聚为一类,因此支持了国际野生百合形态学上的分组,并且说明是否具叶柄对野生百合的分类是一个重要的形态学性状。Nishikawa et al(1999)研究证实很难依据形态特征对百合属进行分类,因为有些种虽然亲缘关系较远,但其形态特征尤其是花型却非常相似,本研究种毛百合、野百合、百合的聚类形式与传统形态分类结果不尽相同,一定程度上印证了此观点 。
本研究证实了ISSR分子标记技术能较好地从分子水平揭示野生百合种间的亲缘关系,肯定了形态学分类的科学性,并发现毛百合与卷瓣组百合的亲缘关系相近,钟花组与卷瓣组野生百合存在基因交流。我国野生百合种类丰富、生态类型多样,曾为百合育种做出了突出贡献。如20世纪初,利用野生百合(岷江百合)的抗逆性,濒临灭绝的欧洲百合得以拯救。现今栽培百合杂种系中的亚洲百合杂种系、麝香百合杂种系、喇叭百合杂种系和白花百合杂种系等均有我国原生种参与杂交而成。因此,进一步加强我国百合遗传多样性与亲缘关系研究,对于野生资源科学保护和有效利用具有重要意义 。
CHEN XQ,XU JM,LIANG SY, et al, 1980. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press: 116. [陈心启,许介眉,梁松筠, 等, 1980. 中国植物志 [M]. 北京: 科学出版社: 116.]
COMBER HF, 1949. A new classification of the genusLilium[M]. Lily Yearbook, Royale Horticulture Society(London), 15: 86-105.
DEVARUMATH RM, NANDY S, RANI V, et al, 2002. RAPD, ISSR and RFLP fingerprintings as useful markers to evaluate genetic integrity of micropropagated plants of three diploid and triploid elite tea clones representingCamelliasinensis(China type)andC.assamicassp.Assamica(Assam-India type) [J]. Plant Cell Rep, 21: 166-173.
DUBOUZET JG, SHINODA K, 1999. Phylogenetic analysis of the internal transcribed spacer region of JapaneseLiliumspecies [J]. Theor Appl Genet, 98: 954-960.
GU X, ZHANG YL, NIU LX, et al, 2013. Pollen morphology observation of 15 wild lilies from four provinces in Western China [J]. Acta Hortic Sin, 40(7): 1 389-1 398. [顾欣, 张延龙, 牛立新, 等, 2013. 中国西部四省15种野生百合花粉形态研究 [J]. 园艺学报, 40(7): 1 389-1 398.]
GUO WH, JEONG JH, KIM ZS, et al, 2011. Genetic diversity ofLiliumtsingtauensein China and Korea revealed by ISSR markers and morphological characters [J]. Biochem Syst Ecol, 39(4-6): 352-360.
LEE CS, KIM SC, YEAU SH,et al, 2011. Major lineages of the genusLilium(Liliaceae)based on nrDNA sequences, with special emphasis on the Korean species [J]. J Plant Biol, 54: 159-171.
MICHAEL JB, HARRIS H, 2002. The gardener’s guide to growing lilies [M]. Oregon: Timber Press: 1-160.
NISHIKAWA T, OKAZAKI K, UCHINO T, et al, 1999. A molecular phylogeny ofLiliumin the internal transcribed spacer region in nuclear ribosomal DNA [J]. J Mol Evol, 49: 238-249.
PRADEEP RM, SARLA N, SIDDIQ EA, et al, 2002. Inter simple sequence repeat(ISSR) polymorphism and its application in plant breeding [J]. Euphytica, 128: 9-17.
QIAN W, GE S, HONG DY,et al, 2001. Genetic variation within and among populations of a wild riceOryzagranulatafrom China detected by RAPD and ISSR markers [J]. Theor Appl Genet, 102: 440-449.
TSUMURA Y, OHBA K, STRAUS SH, et al, 1996. Diversity and inheritance of inter-simple sequence repeat polymorphisms in Douglas-fir (Pseudotsuga menziesii) and sugi (Cryptom eria japonica) [J]. Theor Appl Genet, 92: 40-45.
XIANG DY, ZHANG YL,HAO RJ, et al, 2006. Study on the characters description ofLiliumfrom Qinba Mountain and its adjacent area [J]. Chin Agric Sci Bull,22(10): 97-100. [向地英, 张延龙,郝瑞杰, 等, 2006. 秦巴山区及毗邻地区野生百合性状描述 [J]. 中国农学通报,22(10): 97-100.]
YAMAGISHI M, ABE H , NAKANO M, et al, 2002. PCR-based molecular markers in Asiatic hybrid lily [J]. Sci Hortic-Amsterdam, 96: 225-234.
ZHANG YP, WANG JH, WU LF, et al, 2014. The karyotype of wild and horticultural varieties of lily [J]. J Fujian Agric For Univ, 43(4): 370-373. [张艺萍, 王继华, 吴丽芳,等, 2014. 百合属野生种和园艺品种的染色体核型 [J]. 福建农林大学学报·自然科学版, 43(4): 370-373.]
ZHAO XY,WANG SD,CHEN XL,et al, 1999.Lilium[M]. Beijing:China Agriculture Press:1. [赵祥云,王树栋,陈新露,等, 1999. 百合 [M]. 北京:中国农业出版社:1.]
ZHI L, TENG ZH, LI XY, et al, 2011. Phylogenetic relationship analysis of 23 wild species ofLiliumby SRAP markers [J]. J Agric Biotechnol, 19(4): 677-684. [智利, 滕中华, 李先源, 等, 2011. 23种野生百合遗传关系的SRAP分析 [J]. 农业生物技术学报, 19(4): 677-684.]ZIEKIEWICZ E, RAFALSKI A, LABUDA D, et al,1994. Genome fingerprinting by simple sequence repeat(SSR)-anchored polymerase chain reaction amplification [J]. Genomics, 20: 176-183.
Genetic relationship analysis of 21 wild species ofLiliumby ISSR markers
HOU Jun, LUO Jian-Rang, XIAO Fei-Fei, ZHANG Yan-long*, NIU Li-Xin
(CollegeofLandscapeArchitectureandArts,NorthwestA&FUniversity, Yangling 712100, China )
To probe the infrageneric phylogenetic relationship ofLilium, seven inter-simple sequence repeat (ISSR) primers were chosen to analyze genetic diversity of 21 wild species ofLilium. POPGENE1.32 and MEGA5.1 were used for data analysis. Results showed that a total of 149 clear DNA bands were amplified, 136 of which were polymorphic, the proportion was 91.06%. The average value of effective number of alleles was 1.762 4. The average value of Nei’s gene diversity was 0.421 4, and the average value of Shannon's Information index was 0.608 5. The genetic distance ranged from 0.307 5 to 0.887 3. The genetic distance betweenL.leucanthumandL.leucanthumvar.centifoliumwas a maximum of selected 21 parts of wild lily material, average value 0.887 3, which indicated that the genetic relationship between them was the fartherest. Whereas the genetic distance betweenL.leucanthumvar.centifoliumandL.lophophorumwas a minimum of selected 21 parts of wild lily material, average value 0.307 5, which indicated genetic relationship between them was the closest. The clustering results were basically consistent with the morphology classification. Twenty-one wildLiliumspecies were classified into five clustering groups by neighbor joining method.L.taliense,L.duchartrei,L.lancifolium,L.nepalense,L.papilliferum,L.davidii,L.davidiivar.unicolor,L.pumilum,L.fargesiiandL.dauricumwere clustered in the first group. AndL.leucanthumBaker var.centifolium,L.leucanthum,L.nepalense,L.regale,L.sargentiaebelong to the second group.L.brownie,L.browniivar.viridulumwere clustered in the third group.L.tsingtauenseformed a distinct group andL.lophophorum,L.amoenum,L.concolorvar.pulchellumwere clustered in the fifth group.L.dauricumhas close genetic relationship with sect. Sinomartagon, and there were gene exchanges existed between Sect. Lophophorum and Sect. Sinomartagon. This indicated that it was not accurate to classify the wildLiliumall depend on the morphology.L.browniivar. viridulum andL.browniewere clustered into one individual group in cluster analysis results. This result showed that whether it has petioles was an important morphological character in the classification of wild lilies. And all the results indicated that ISSR markers were suited to study the phylogenetic relationship of wild lilies.
Lilium, wild lilies, ISSR, genetic relationship, tree plot
10.11931/guihaia.gxzw201504028
2015-04-19
2015-05-19
国家林业局重点科研项目(2006-73);陕西省林业厅项目(陕林计字 [2011]70号);国家自然科学基金(305021110);西北农林科技大学基本科研业务费专项(Z109021002) [Supported by Key Scientific Research Project of State Forestry Administration (2006-73); Project of Shaanxi Forestry Department ( [2011]70); National Natural Science Foundation of China (305021110); Foundamental Scientific Research Fund of Northwest A & F University (Z109021002)]。
侯珺(1990-),女,陕西渭南人,硕士,主要从事野生百合遗传多样性研究,(E-mail)346144734@qq.com。*
张延龙,教授,主要从事园林植物资源与育种研究,(E-mail) zhangyanlong@nwsuaf.edu.cn。
Q949
A
1000-3142(2016)09-1032-07
侯珺, 罗建让, 肖菲菲, 等. 21种野生百合亲缘关系的ISSR分析 [J]. 广西植物, 2016, 36(9):1032-1038
HOU J, LUO JR, XIAO FF,et al. Genetic relationship analysis of 21 wild species ofLiliumby ISSR markers [J]. Guihaia, 2016, 36(9):1032-1038
