亚热带毛竹林生态系统碳通量与土壤呼吸研究
2016-10-26张敏霞陈晓峰黄鹤凤舒海燕吴梦玲浙江农林大学林业与生物技术学院浙江临安00南京大学国际地球系统科学研究所南京2009西南大学生命科学学院三峡库区生态环境教育部重点实验室重庆40075
张敏霞,江 洪,2*,陈晓峰,黄鹤凤,舒海燕,吴梦玲(.浙江农林大学林业与生物技术学院,浙江临安00;2.南京大学国际地球系统科学研究所,南京2009;.西南大学生命科学学院三峡库区生态环境教育部重点实验室,重庆40075)
亚热带毛竹林生态系统碳通量与土壤呼吸研究
张敏霞1,江洪1,2*,陈晓峰1,黄鹤凤1,舒海燕3,吴梦玲1
(1.浙江农林大学林业与生物技术学院,浙江临安311300;2.南京大学国际地球系统科学研究所,南京210093;3.西南大学生命科学学院三峡库区生态环境教育部重点实验室,重庆400715)
以浙江省安吉县毛竹林(Phyllostachys edulis)生态系统为研究对象,利用开路涡度相关系统和LI-Cor 8150自动观测系统,分析2014年毛竹林生态系统碳通量和土壤呼吸速率变化特征及其影响因子。结果表明,毛竹林土壤呼吸速率日变化为单峰曲线,最高值出现在14:00~16:00,最低值出现在06:00;净生态系统交换量(NEE)存在明显日变化特征,变化趋势为“双峰曲线”,峰值分别出现在10:00和12:00;而生态系统呼吸(RE)和土壤呼吸速率呈相同的年变化趋势,为“单峰型”,夏季高、冬季低,且均对温度变化较敏感。毛竹林土壤呼吸速率和生态系统呼吸量(RE)主要受高于20℃大气温度和5 cm土壤温度影响,与水分相关关系不显著。
亚热带;毛竹林生态系统;碳通量;土壤呼吸
网络出版时间2016-6-17 15:21:48[URL]http://www.cnki.net/kcms/detail/23.1391.S.20160617.1521.006.htm l
Zhang Minxia,Jiang Hong,Chen Xiaofeng,et al.Study on the carbon flux and soil respiration of Phyllostachys edulis forest ecosystem in subtropic China[J].Journal of Northeast Agricultural University,2016,47(6):41-50.(in Chinese with English abstract)
植物通过光合作用固定大气CO2,并通过自身呼吸作用及土壤微生物呼吸、凋落物分解、森林采伐等途径将储存碳重新释放到大气中[1]。土壤-植被-大气间净碳交换(Net ecosystem exchange,NEE)及其对环境因子响应研究是理解陆地生态系统碳循环过程关键[2]。陆地生态系统碳通量作为影响碳循环关键因素,已成为全球变暖研究核心内容之一[3]。陆地生态系统中土壤是最大碳库(约含1 600 Pg),约为大气中碳的2倍,是活植物体碳库3倍,仅次于总初级生产力,是陆地生态系统碳循环重要组成部分[4-5]。因此,全球土壤呼吸的微小变化将对大气CO2浓度产生巨大影响[6-8]。土壤呼吸作为影响全球气候变化的关键生态过程,已成为全球碳循环研究核心问题[9],但目前我国土壤碳储量估算尚不确定,因此了解和掌握土壤碳储量转化过程和机理,是陆地生态系统碳循环研究重要前提[10]。
毛竹是我国重要经济竹种,生长快、产量高、用途广、固碳能力强,是我国重要森林资源之一[11],主要分布在热带和亚热带,毛竹是我国面积最大竹种[12-13]。因毛竹有高生态和社会经济多功能性,尤其是其巨大固碳潜力,在减缓气候变化中起重要作用,近年受到广泛关注[13]。周国模等研究发现,毛竹固碳能力远高于其他林木,毛竹林每年固碳量是杉木林、马尾松林和热带雨林2~4倍[14]。因此,毛竹生态系统在区域和全球CO2平衡及气候稳定方面作用重要。
土壤呼吸作为毛竹林生态系统碳循环重要环节,通过生态系统总呼吸量影响毛竹林生态系统碳通量,因此本文通过研究安吉毛竹林生态系统碳通量及土壤呼吸变化特征,一定程度上可从土壤呼吸角度解释毛竹林碳通量变化过程,估测亚热带毛竹林生态系统土壤呼吸对该生态系统碳通量影响,为定量把握影响亚热带毛竹林生态系统碳循环环境控制机制,准确估算土壤碳通量变化提供理论依据。
1 研究地区与研究方法
1.1研究地区概况
试验区位于浙江省湖州市安吉县山川乡,森林面积4 251 hm2,其中竹林面积2 155 hm2,占森林总面积50.7%,毛竹林面积1 693 hm2,占竹林总面积78.6%[15]。地理位置30°28'34.5''N,119°40' 25.7''E,属亚热带季风气候,全年雨水充沛,温暖湿润,四季分明,雨热同期。年降水量761~ 1 780mm,主要集中在6~8月,12月最少,具有明显双峰型降雨特点,各年份梅雨量差异较大[16]。年平均气温16.6℃,7月最高,1月最低。年日照1 980~2 050 h,年无霜期203~226 d。试验区内毛竹林为人工纯林,林分密度4 500株·hm-2,植物类型以毛竹为主,林下有少量马尾松、杉木及极少灌木和草本;土壤类型为黄壤。毛竹节间短,壁厚,出笋期为3~5月,且连年出笋能力较强,6月份为大量展叶期,9月份为生长季[15-16]。
1.2研究方法
1.2.1CO2通量测定
观测林地微气象观测塔高40m,开路涡度相关系统(OPEC)探头安装在距地面38m处(约3倍冠层高度),由三维超声风速仪(CAST3,Campbell Inc.,USA)和开路CO2/H2O分析仪(Li-7500,Li-Cor Inc.,USA)组成,原始采样频率10 Hz,通过数据采集器(CR1000,Campbell Inc.,USA)存储数据,并在线计算存储30min CO2通量(Fc)等相关物理量。常规气象观测系统包括7层风速(010C,metone,USA)、7层大气温度和湿度(HMP45C,Vaisala,Helsinki, Finland),安装高度分别为1、7和38m;2个SI-111红外温度计分别置于2m和23m处,用于采集地表和冠层温度;净辐射仪(CNR4,Kipp&Zonen)传感器安装高度38m,用于采集上行/下行长波/短波辐射、净辐射数据。常规气象观测系统数据采样频率为5 Hz,通过数据采集器(CR1000、Camp⁃bell Inc.、USA)每30min自动记录平均风速、湿度、温度、气压、净辐射等常规气象信息。本研究中空气温度、空气湿度与饱和水汽压差选用7m高冠层气象数据。仪器设备详情参照文献[16-17]。
关于植被与大气间CO2通量计算,利用三维超声风速仪测定三维风速,利用红外线CO2/H2O气体分析仪测定CO2密度脉动,将所有10 Hz原始数据利用数据采集器记录并储存。采用EdiRe(Eddy Reprocessing)软件进行协方差运算,并作二次坐标轴旋转、WPL(Wdbb-Pearman-Leuning)修正等,最终将10 Hz原始数据转化为30min数据[17-19]。即植被与大气间净生态系统CO2交换量(NEE,mgCO2·m-2·s-1)定义为:

式中,Fc(mgCO2·m-2·s-1)为CO2湍流通量,分析通量塔数据直接得到;Fs(mgCO2·m-2·s-1)为观测高度以下碳储存通量;h为观测高度(40m);Δc为高度h处前后两次相邻时间测定CO2浓度差值(mgCO2·m-3);Δt为前后两次测定时间间隔(s)。当CO2从生态系统排放大气中时,定义NEE符号为正,当CO2从大气进入到生态系统时,定义NEE为负。利用涡度相关法仅能直接获取NEE,而生态系统呼吸(Re)、生态系统总生产力(GEP)和大气间净生态系统CO2交换量之间关系用下列方程描述[17,

式中,RE为生态系统呼吸量(Ecosystem respi⁃ration,RE),GEE总生态系统交换量(Gross ecosys⁃tem exchange,GEE)。
1.2.2土壤呼吸测定方法
采用美国LI-COR公司生产LI-Cor 8150开路式土壤碳通量自动测量系统(LI-COR,USA)测定土壤呼吸。通量塔观测的毛竹林样地内设置1个10m×10m标准样地,随机选取3个2m×2m小样方,安置PVC土壤环(直径20 cm,高10 cm),每个土壤环露出地表3~5 cm。为减小对土壤扰动,提前24 h将土壤环置于样方内,去除环内植物活体,确保测试环周围不漏气,测定期间保持土壤环位置不变。同时,土壤温度和湿度由该测量系统携带探针同步测定,探针插入土表5 cm深处,分别测得相应位置土壤温度和土壤体积含水量。测定时间为2014年1~12月,每月中旬选取1周时间,24 h昼夜不间断测量土壤呼吸,仪器设备详情参考文献[21]。
1.3数据处理
数据经Excel 2007筛选后SPSS 13.0软件处理分析,通过Excel2007绘图。土壤CO2通量值均为相应时段多次测量所取平均值。运用统计分析软件IBM SPSS Statistics 21.0对数据作显著性分析,方差齐性显著性检验采用LSD法(α=0.05),单因素方差分析(one way ANOVE)检验土壤呼吸速率动态变化显著性。采用指数回归模型分析土壤呼吸速率与土壤温度和大气温度关系,一般线性模型分析土壤呼吸速率与土壤含水量及其他环境因子关系。
2 结果与分析
2.1环境因子变化
研究地属典型亚热带季风气候,因此受季风气候影响,环境因子表现明显季节变化规律。由图1(A)可知,大气和土壤温度均呈单峰型变化趋势,土壤温度随大气温度升高而逐渐升高,夏季达全年最高温,冬季最低。其中大气平均温度为(26.51±1.42)℃,土壤温度为(23.59±0.92)℃,大气和土壤温度差异不显著(P<0.05)。大气湿度变化呈倒“W”型变化趋势,降雨多的月份空气湿度相对较大(见图1B)。图1(C)为2013年毛竹林生态系统降雨量和地下5 cm土壤含水量季节变化规律。由图1(C)可知,该地区全年雨水充沛,夏季降雨集中。观测期内观测林地年降雨总量为1 508.5mm,其中6月份发生1次强降雨,月总降雨量为394.5mm,占夏季总降雨量64.75%;土壤含水量在18.61~30.71m3·m-3波动,与降雨量呈相同变化趋势,其大小随降雨量变化,但不同月份差异较大,无明显变化规律,受温度和降雨量共同影响。

图1 安吉毛竹林生态系统环境因子变化特征Fig.1 Variation of environmental factors of of Phyllostachys edulis forest ecosystem in Anji country
2.2亚热带毛竹林生态系统碳通量和土壤呼吸速率日平均变化特征
李雅红等研究发现,毛竹林土壤呼吸速率呈明显日变化特征[22]。图2(A)将一年中春季(3~5月)、夏季(6~8月)、秋季(9~11月)、冬季(12月至翌年2月)每个季节3个月平均日变化再次平均,得出4个季节毛竹林生态系统土壤呼吸速率日变化曲线,毛竹林土壤呼吸速率日变幅小,整体表现平缓单峰型变化趋势,峰值出现在14:00~16:00,之后又逐渐降低,直到18:00后土壤呼吸速率趋于平缓,基本保持不变。而最小土壤呼吸速率值出现在06:00左右。
受到季节性变化,土壤有机碳含量和植物物候期等因素影响,土壤呼吸日变化在不同季节表现不同模式,本试验春、夏、秋、冬4个季节土壤呼吸日动态变化范围分别为:2.24~2.75、5.10~ 6.29、3.21~3.73和0.63~1.10μmol·m-2·s-1,变化依次为:夏季>秋季>春季>冬季。
同样4个季节每个月碳通量相同时刻数据作平均处理,得到4个季节毛竹林生态系统碳通量日变化曲线,碳通量为正值表示生态系统排放CO2,负值表示生态系统进行光合作用吸收CO2,因此正值越大表明生态系统呼吸作用越强烈,释放CO2越多。由图2(B)可知,毛竹林碳通量具有明显日变化特征,变化趋势为“双峰曲线”,峰值分别出现在10:00和12:00左右,夜间表现为碳释放,白天表现为碳吸收,早晨碳通量由正转负时间段为6:00~ 8:00,傍晚由负转正时间段为17:00~18:00,且夏季由正转负出现时间较早,由负转正出现时间较晚,而冬季净碳交换量时间均短于夏季。4个季节白天均为碳吸收,夜间均为碳排放,即生态系统吸收CO2量大于释放CO2量。
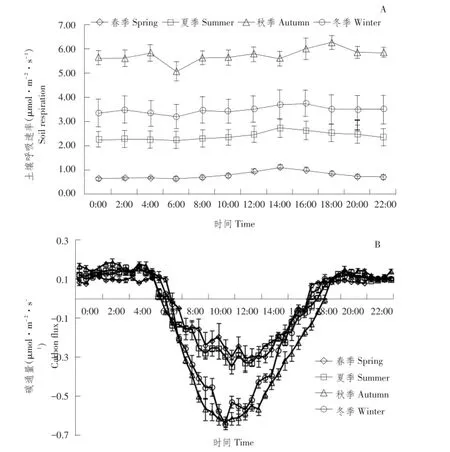
图2 不同月份土壤呼吸和碳通量平均日变化Fig.2 Diurnal variations of monthly-mean soil respiration and carbon flux
2.3亚热带毛竹林生态系土壤呼吸速率和生态系统呼吸(RE)年变化特征
毛竹林土壤呼吸速率变化和林内植物生长密切相关,呈现一定季节变化规律,如图3(A)所示,2014年毛竹林生态系统土壤呼吸速率年变化和大气温度、土壤5 cm温度变化趋势一致,呈“单峰型”,3月份气温逐渐回升,植物生长活跃土壤呼吸速率开始增大,夏季水热条件较好,毛竹生长旺季,因此土壤呼吸速率也达到全年最大值,其中8月份最高为6.67μmol·m-2·s-1,成为全年土壤呼吸速率转折点,之后进入秋季,温度逐渐降低,伴随毛竹生理能力逐渐减弱,土壤呼吸速率开始下降。受环境因子影响土壤呼吸速率最低值出现在1月份为0.6μmol·m-2·s-1,夏季高于冬季。而生态系统呼吸变化趋势和土壤呼吸速率变化趋势一致,也呈单峰型变化趋势,夏季最大,冬季最小。由此可见,毛竹林生态系统土壤呼吸速率极大影响生态系统呼吸。
2.4土壤呼吸速率和环境因子关系
土壤5 cm温度、土壤体积含水量、大气温度、湿度等均是导致土壤呼吸变化的重要环境因子[21]。为明确影响安吉毛竹林生态系统土壤呼吸速率主要环境因子,对土壤呼吸速率和5 cm土壤温度、大气温度、地下5 cm土壤含水量、大气湿度作相关性分析。由图4可知,土壤呼吸速率随5 cm土壤温度和大气温度升高而呈指数增长,其增长关系可用指数模型(y=aebx)描述,即温度越高,土壤呼吸速率越大,当5 cm土壤温度和大气温度均大于20℃时,变化趋势更明显,土壤呼吸速率和土壤5 cm温度、大气温度相关性较好,均达极显著(P<0.01),相关系数分别为0.94和0.87,土壤5 cm温度和大气温度对土壤呼吸影响分别可解释土壤呼吸的93%和87%。同时线性拟合分析土壤呼吸速率与地下5 cm含水量及大气湿度相关性。结果表明,土壤呼吸速率季节性变异与降雨量和大气湿度关系不大,相关系数仅为0.023和0.026,相关性不显著(P>0.05)。
2.5生态系统呼吸(RE)和环境因子关系
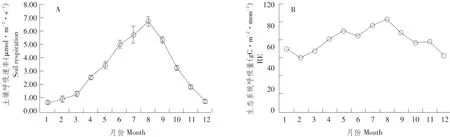
图3 土壤呼吸速率和生态系统呼吸量季节变化Fig.3 Seasonal variation of soil respiration and ecosystem respiration
由图5可知,大气和土壤温度与生态系统呼吸相关性达极显著(P<0.01),大气温度和土壤温度与生态系统呼吸相关系数分别为0.87和0.86,5 cm土壤含水量和生态系统呼吸呈显著正相关,即土壤含水量升高将促进生态系统呼吸,而空气湿度与生态系统呼吸相关性不显著(P>0.05),表明研究期温度是控制生态系统呼吸主要环境因子,大气湿度并非生态系统呼吸限制因子。
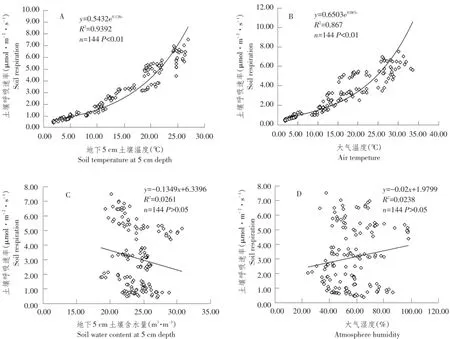
图4 土壤呼吸速率和环境因子关系Fig.4 Relationship between soil respiration and environmental factors
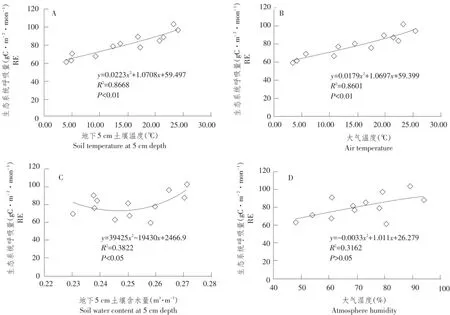
图5 生态系统呼吸量和环境因子关系Fig.5 Relationship between ecosystem respiration and environmental factors
2.6NEE、RE与Rs相关性比较
本文利用月尺度土壤呼吸速率分别与生态系统净碳交换和生态系统呼吸拟合,由图6(A)可知,随土壤呼吸速率不断升高,生态系统净碳交换逐渐增大(数值上表现为负方向增大),达到一定值时,土壤呼吸速率继续增大,但净碳交换不再增大,并有减小趋势(负方向减小),即生态系统净碳交换减少。
由图6(B)可知,土壤呼吸速率增大,将直接导致生态系统呼吸增大,即随毛竹林生态系统土壤呼吸速率增加,生态系统呼吸量不断增加,而生态系统净碳交换将逐渐降低。
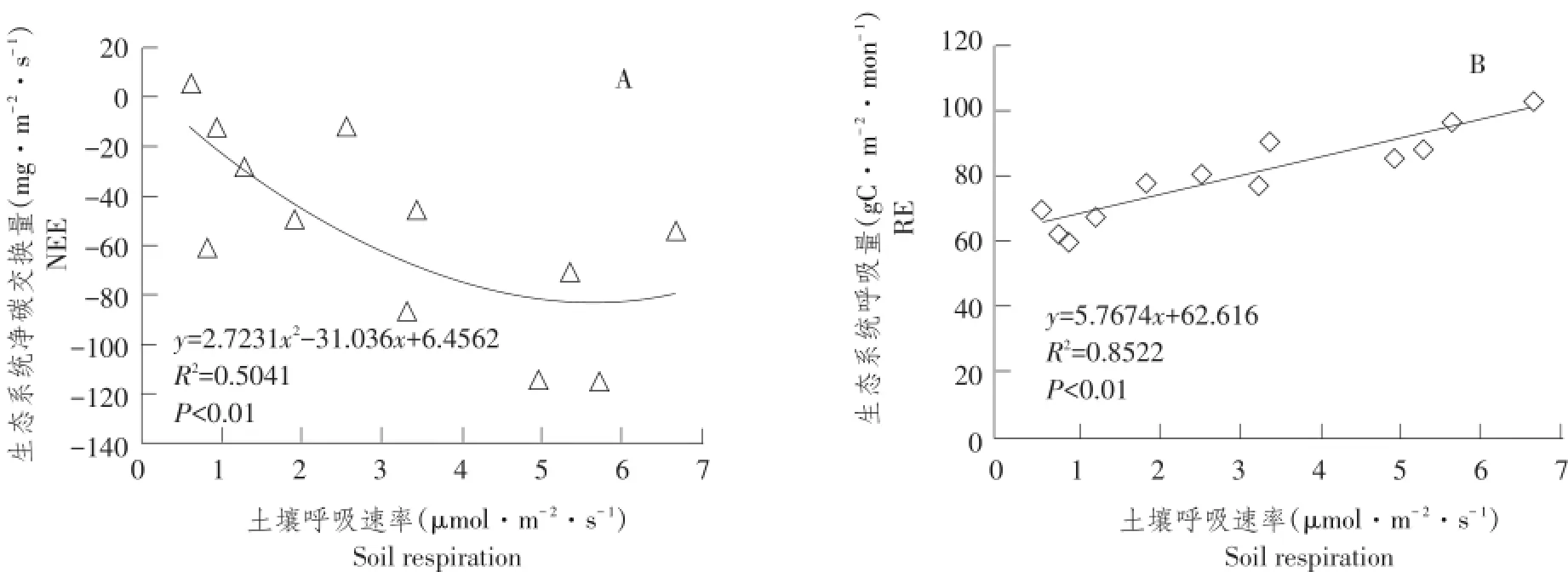
图6 生态系统碳通量和土壤呼吸速率关系Fig.6 Relationship between soil respiration and ecosystem carbon flux
3 讨论
3.1生态系统碳通量和土壤呼吸速率日变化规律
安吉毛竹林土壤呼吸速率具有明显日变化特征。总体而言,土壤呼吸日变化波动不大,变化趋势呈“单峰曲线”,最大值出现时间为14:00~ 16:00,滞后温度最大值出现时间。主要由于地表温度随大气温度升高逐渐升高,但地表温度升高需要一定时间[23]。研究期,四个季节土壤呼吸速率大小依次为:夏季>秋季>春季>冬季,春季和夏季植物生长旺盛,根系活动活跃,呼吸产生量增加,微生物代谢活动加强,促使异养呼吸成分增加。冬季无论温度、湿度和生物生长量,均低于春夏季,这些因子共同作用降低呼吸速率[23]。本研究土壤呼吸速率季节变化结果与李雅红等西天目山毛竹林土壤呼吸研究结果一致[22]。
同土壤呼吸速率一样,研究期毛竹林NEE月平均日变化具有明显日动态。碳吸收时间段为6:00~ 18:00,夏季NEE由正转负时间最早,由负转正时间最晚,而冬季由正转负时间最晚,由负转正时间均是最早,主要是因为夏季日照长,生态系统光合作用时间长,而冬季光照时间短于夏季。研究期生态系统白天均表现为碳汇,夜间表现为碳源。白天植物光合作用大量吸收CO2,NEE为负值,夜间毛竹光合作用停止,生态系统只进行呼吸作用释放CO2,NEE均为正值。4个季节NEE大小依次为:夏季>秋季>春季>冬季,NEE大小季节变化变化规律反应生态系统生物学特性。
3.2土壤呼吸速率和生态系统呼吸年变化规律
本研究还发现生态系统呼吸量和土壤呼吸速率季节变化规律与该地区温度季节变化规律一致,春季随温度升高土壤呼吸速率逐渐升高,夏季达全年最大值,秋季开始降低,冬季为全年最低值。主要是因为春季和夏季植物生长,根系活动活跃,呼吸产生量增加,微生物代谢活动加强,促使异养呼吸成分增加。冬季无论温度、湿度和生物生长量,均低于春夏季,这些因子共同作用降低呼吸速率[24]。
3.3环境因子对毛竹林土壤呼吸速率影响
大量研究表明,在一定范围内,温度升高将加速植物和微生物新陈代谢活动,促进生态系统碳排放,是影响生态系统呼吸主要环境因子[2]。土壤温度对土壤呼吸影响主要是通过增强微生物活性,加速土壤有机质分解增加土壤CO2浓度,温度升高可影响植物生长和生理活动从而加强根系呼吸作用[25]。本研究发现毛竹林生态系统呼吸和土壤呼吸速率月变化模式,与土壤温度和大气温度密切相关,这与吴蒙[25]、刘源月[26]、Song[27]等研究结果一致。土壤温度和大气温度与土壤呼吸速率均呈指数相关,土壤呼吸速率随温度升高而升高,且土壤温度和大气温度分别可解释土壤呼吸速率变化94%和87%,从决定系数R2看,土壤呼吸速率与5 cm土壤温度相关性最好,主要因为土壤呼吸是一个受生物和非生物因素控制的复杂过程,处于5 cm土壤表层生物和非生物因素如植物根系、微生物等活动最旺盛,因此对土壤呼吸速率贡献较大,导致土壤呼吸随土壤5 cm温度变化一致[22]。
李熙波等研究认为,在众多环境因子中土壤温度和水分是土壤呼吸重要影响因素[28],土壤水对土壤形成过程、剖面发育和物质与能量运转有重要影响,是毛竹生长和生存物质基础[18]。龚斌等在中亚热带森林土壤呼吸日变化及其与土壤温湿度关系报道中指出,毛竹林土壤呼吸速率和土壤温度之间无相互关系,而土壤呼吸速率随含水量增加而降低[29]。本研究发现土壤含水量和湿度与土壤呼吸速率相关性不显著,因为本研究地处亚热带,全年雨水充沛,土壤水分较饱和,水分是影响毛竹林土壤呼吸速率限制因子,因此土壤湿度与土壤呼吸相关性不显著,这与李雅红[22]、王国兵[30]等研究结果一致。存在差异可能由地区、试验和采样条件不同造成。
3.4环境因子对毛竹林生态系统呼吸量(RE)影响
乔明阳等认为土壤温度与湿度是影响生态系统呼吸关键因子[31]。本文前面分别探讨温度和水分等环境因子对生态系统呼吸量影响,结果发现,生态系统呼吸与大气温度和5 cm土壤温度均达极显著,生态系统呼吸月变化和温度变化趋势一致,温度变化分别可解释生态系统呼吸变化87%和86%。孙小祥等曾指出,在一定范围内,温度升高将加速植物和微生物新陈代谢活动,促进生态系统碳排放,影响生态系统呼吸[2]。由此可见,温度升高,将促进毛竹林生态系统呼吸作用。安吉毛竹林生态系统中温度对土壤呼吸速率影响与陆地生态系统呼吸相一致,但5 cm土壤含水量和土壤湿度对土壤呼吸速率影响与生态系统呼吸不一致,土壤呼吸速率对生态系统呼吸影响达显著水平,大气湿度对生态系统呼吸影响更明显。
3.5土壤呼吸对生态系统呼吸和碳交换量影响
土壤呼吸是全球森林生态系统重要碳通量,土壤呼吸速率变化将影响全球气候格局分布[18,25],本研究通过拟合分析土壤呼吸速率和生态系统呼吸及生态系统碳交换关系,发现毛竹林土壤呼吸速率增加,直接导致生态系统呼吸和净碳交换增加,但生态系统呼吸随土壤呼吸速率增加基本呈线性升高,而土壤呼吸速率过快增长,使生态系统碳交换减小。
4 结论
①亚热带毛竹林生态系统碳通量和土壤呼吸速率具有明显日变化及季节性变化规律,变化趋势均呈“单峰”,土壤呼吸速率日变化范围为:0.63~ 6.29μmol·m-2·s-1,日最大值出现在下午14: 00~16:00,年变化范围为:2.49~29.7μmol·m-2· s-1,不同季节碳通量和土壤呼吸速率日最大值出现时间不同,主要是由于环境因子等季节性变化引起。
②在影响土壤呼吸速率和生态系统呼吸量众多环境因子中,温度是最主要限制因子,5 cm土壤温度和大气温度与土壤呼吸速率相关系数分别为94%和87%,均达极显著,5 cm土壤温度和大气温度对生态系统呼吸量影响也达极显著,相关系数分别为87%和86%,生态系统呼吸与土壤含水量相关达显著水平,但与大气湿度相关性不显著,说明温度是影响该生态系统土壤呼吸和碳通量主要因子。
③土壤呼吸速率和生态系统呼吸量均随温度升高而逐渐升高,土壤温度与土壤呼吸速率相关关系可用指数模型y=aebx表示。
④毛竹林生态系统呼吸随毛竹林土壤温度升高而逐渐增加,毛竹土壤呼吸速率升高将直接导致生态系统呼吸增加;生态系统净碳交换也随土壤呼吸速率增加而增加,当土壤呼吸速率超过一定值时,生态系统净碳交换随土壤呼吸速率增加而下降。
[1]周丽艳,贾丙瑞,周广胜,等.中国北方针叶林生长季碳交换及其调控机制[J].应用生态学报,2010,21(10):2449-2456.
[2]孙小祥,常志州,杨桂山,等.长三角地区稻麦轮作生态系统净碳交换及其环境影响因子[J].中国生态农业学报,2015,23(7): 803-811.
[3]范连连,谢继萍,马健,等.准噶尔盆地南缘草本层碳通量及土壤呼吸的变化特征[J].生态学杂志,2013,32(10):2567-2573.
[4]路亚坤,刘加珍,陈永金,等.东平湖湖滨带农田土壤碳通量的动态变化[J].贵州农业科学,2013,41(5):104-108.
[5]王国兵,郝岩松,王兵,等.土地利用方式的改变对土壤呼吸及土壤微生物生物量的影响[J].北京林业大学学报,2006,12 (2):73-79.
[6]Song X Z,Yuan H Y,Mark OK,et al.Soil CO2flux dynamics in the two main plantation forest types in subtropical China[J].Sci⁃ence of the Total Environment2013,444:363-368.
[7]冯文婷,邹晓明,沙丽清,等.哀牢山中山湿性常绿阔叶林土壤呼吸季节和昼夜变化特征及影响因子比较[J].植物生态学报, 2008,32(1):31-39.
[8]杨兰芳,蔡祖聪.不同生长期盆栽大豆的土壤呼吸昼夜变化及其影响因子[J].生态学报,2004,24(12):2955-2960.
[9]高翔,郝卫平,顾峰雪,等.降雨对旱作春玉米农田土壤呼吸动态的影响[J].生态学报,2012,32(24):7883-7893.
[10]许信旺,潘根兴,侯鹏程.不同土地利用对表层土壤有机碳密度的影响[J].水土保持学报,2005,19(6):193-200.
[11]黄真娟,江洪.安吉毛竹春秋季光合特性及碳同化量[J].福建林业科技,2014,41(4):1-6.
[12]Song X Z,Zhou GM,Jiang H,etal.Carbon sequestration by Chi⁃nese bamboo forests,and their ecological benefits:Assessment of potential,problems,and future challenges[J].Environ Rev,2011, 19(1):418-428.
[13]黄鹤凤.浙西北典型竹林土壤呼吸特征及其环境因子的研究[D].临安:浙江农林大学,2015.
[14]周国模,吴家森,姜培坤.不同管理模式对毛竹林碳贮量的影响[J].北京林业大学学报,2006,28(6):51-55.
[15]刘玉莉,江洪,周国模,等.安吉毛竹林水汽通量变化特征及其与环境因子的关系[J].生态学报,2014,34(17):4900-4909.
[16]杨爽.浙江安吉毛竹林生态系统CO2通量观测研究[D].临安:浙江农林大学,2012.
[17]孙成,江洪,周国模,等.我国亚热带毛竹林CO2通量的变异特征[J].应用生态学报,2013,24(10):2717-2724.
[18]吴家森,胡睦荫,蔡庭付,等.毛竹生长与土壤环境[J].竹子研究汇刊,2006,25(2):3-6.
[19]陈晓峰,江洪,牛晓栋,等.季节性高温和干旱对亚热带毛竹林碳通量的影响[J].应用生态学报,2016,27(23):335-344.
[20]陈晓峰,江洪,孙文文,等.安吉毛竹林生长季CO2浓度的时空特征[J].生态学杂志,2016,35(5):1-8.
[21]黄鹤凤,江洪,谭有靖,等.不同经营措施下太湖源雷竹林土壤呼吸的研究[J].浙江林业科技,2014,34(5):74-80.
[22]李雅红,江洪,原焕英,等.西天目山毛竹林土壤呼吸特征及其影响因子[J].生态学报,2010,30(17):4590-4597.
[23]黄湘,陈亚宁,李卫红,等.塔里木河中下游柽柳群落土壤碳通量及其影响因子分析[J].环境科学,2006,27(10):1934-1940.
[24]何都良,李涵,王亚萍.北亚热带落叶阔叶林土壤呼吸时间变化特征[J].2015,52(5):1674-7070.
[25]吴蒙,马姜明,梁士楚,等.桂林市尧山桉树及马尾松林春、夏两季土壤碳通量特征[J].水土保持通报,2015,35(1):303-310.
[26]刘源月,江洪,邱忠平,等.亚热带毛竹林生态系统土壤呼吸[J].西南交通大学学报,2009,44(4):590-594.
[27]Song X Z,Peng CH,Zhao ZG,et al.Quantification of soil respi⁃ration in forest ecosystems across China[J].Atmospheric Environ⁃ment,2014,94(1):546-551.
[28]李熙波,杨玉盛,曾宏达,等.亚热带沟叶结缕草草坪土壤呼吸[J].生态学报2011,31(8):2096-2105.
[29]龚斌,王风玉,张继平,等.中亚热带森林土壤呼吸日变化及其与土壤温湿度的关系[J].生态环境学报,2013,22(8):1275-1281.
[30]王国兵,唐燕飞,阮宏华,等.次生栎林与火炬松人工林土壤呼吸的季节变异及其主要影响因子[J].生态学报,29(2):966-975.
[31]乔明阳,沈程程,石洪华,等.海岛森林土壤碳通量日动态变化特征——以北长山岛为例[J].海洋环境科学,2015,34(3):377-383.
Study on the carbon flux and soil respiration of Phyllostachys edulis forest ecosystem in subtropic China/ZHANG Minxia1,JIANG Hong1,CHEN Xiaofeng1,
HUANG Hefeng1,SHU Haiyan2,WU Mengling
1
(1.School of Forestry and Bio-Technology,Zhejiang Agriculture and Forestry University,Lin'an Zhejiang 311300,China;2.International Earth System Scientific Institute,Nanjing University,Nanjing 210093,China;3.Ministry of Education Key Laboratory of Eco-environments of Three Gorges Reservoir Region,School of Life Science,South West University,Chongqing 400715,China)
Based on the eddy covariance system(EC)and LI-8150 automatic monitoring system,this study observed the variation characteristics and impact factors of carbon flux and soil respiration of Phyllostachys edulis forest ecosystem in 2014 inAnji country,Zhejiang Province.The results showed that soil respiration(RS)appeared a single diurnal pattern with the peaks at 14:00-16:00 and the trough at 06:00.The daily variation of net ecosystem exchange(NEE)also showed an obvious bimodal shape with the peaks appeared in the 10:00 and 12:00,respectively.The seasonal variation of ecosystem respiration(RE)and RS showed the same unimodal pattern with the maximum in summer and the minimum in winter,and were all sensitive to changes in temperature.The yearly RE and RS strongly depended on the air temperature above 20℃and soil temperature at 5 cm depth,but both did not have significant correlation with the soil water.
subtropical forests;Phyllostachys edulis forest ecosystem;carbon flux;soil respiration
Q945.79;S345
A
1005-9369(2016)06-0041-10
2016-03-22
国家自然科学基金项目(61190114,41171324);国家“973”重点基础研究发展规划项目(2011CB302705);上海市科技兴农推广项目[沪农科推字(2013)第5-8号]
张敏霞(1989-),女,硕士研究生,研究方向为植物生理生态和土壤呼吸。E-mail:864276694@qq.com
江洪,教授,博士生导师,研究方向为植物生态学。E-mail:jianghong_china@hotmail.com
