栀子褐斑病病原菌生物学特性研究△
2016-09-25罗光明董艳凯朱玉野胡燕珍龚雨虹王晓云
罗光明,董艳凯,朱玉野,胡燕珍,龚雨虹,王晓云
(江西中医药大学 药学院,江西 南昌 330004)
·中药农业·
栀子褐斑病病原菌生物学特性研究△
罗光明,董艳凯,朱玉野*,胡燕珍,龚雨虹,王晓云
(江西中医药大学 药学院,江西 南昌 330004)
从栀子植株褐斑病叶片病变部位分离到的病原菌。经致病性测定确定该病原菌为葡萄座腔菌属Botryosphaeria的葡萄座腔菌Botryosphaeriadothidea。本试验研究了病原菌的生物学特性,包括不同温度、光照时间、pH值、碳氮源营养对菌丝生长和子实体萌发的影响,并确定了菌丝和子实体的致死温度。结果表明,菌丝生长的温度范围为10~35 ℃,最佳生长温度为30 ℃,菌丝在pH 4~10范围内均能生长,最适pH为5。适宜菌丝生长的碳、氮源分别为可溶性淀粉和酵母膏。光暗交替条件下更适合菌丝生长。子实体在15~40 ℃范围内均可萌发,最佳温度为25 ℃。在pH 3~11范围内子实体均能萌发,最佳pH为9。子实体萌发的最佳碳、氮源分别为可溶性淀粉和硝酸钙。黑暗条件下适宜菌核萌发。子实体和菌丝的致死温度均为60 ℃。
栀子;褐斑病;生物学特性
栀子为茜草科(Rubiaceae)植物栀子GardeniajasninoidesEllis的干燥成熟果实,始载于《神农本草经》,性味苦寒,具有泻火除烦、清热利尿、凉血解毒等功效,临床上常用于治疗热病心烦、湿热黄疸、血淋涩痛、血热出血、火毒疮疡以及扭挫伤等病证[1]。目前,从栀子属植物中分离鉴定出的化合物主要有环烯醚萜类、多糖类、黄酮类、醇类、有机酸酯类、长链烷烃类、和醛类等[2]。栀子目前正值需求旺季,近期市场货源走动较好,价格不断走高,近年来各药厂相继开发出以栀子为主要原料的新的中成药,从而加大了栀子的药用需求量。同时,栀子还被广泛应用于酿酒、香料制造,也被用以提取天然染料和食用色素,还被开发成香水等产品。国外对栀子的需求量也在呈上升趋势,栀子出口数量不断增大,具有很好的市场前景。
褐斑病是栀子常见的病害之一,全年都可发生,严重时植株落叶、落果或枯死[3]。从而导致栀子产量严重下降。从病变部位分离出的致病菌为葡萄座腔菌,姜晓龙[4]曾对葡萄座腔菌进行系统鉴别,李冬[5]曾对病原菌进行过报道,但至今未见栀子褐斑病病原菌生物学特性的研究报道。为明确栀子褐斑病病原菌生物学特性,了解其所致病害的发生规律和制定有效的防治措施提供理论依据。本试验调查、采集、分离了栀子褐斑病的病原菌,对褐斑病病原菌的生物学特性进行了较为系统的研究。
1 材料与方法
1.1 供试材料
栀子褐斑病病原菌为病变组织经常规组织分离法[6]分离培养得到,确定为葡萄座腔菌Botryosphaeriadothidea。
1.2 生物学特性研究
1.2.1 不同温度对菌丝体生长和子实体萌发的影响 用直径无菌打孔器从生长4 d的菌落边缘打取直径6 mm的菌丝块,接种于PDA平板上,每皿接种1个菌丝块。分别于4、10、15、20、25、30、35、40 ℃条件下黑暗培养,每个处理设置3个重复。96 h后用十字交叉法测量菌落直径。
由培养30 d的成熟菌落挑取子实体,接种于PDA平板上,每皿20粒,4次重复,培养条件与菌丝培养条件相同,接种36 h后观察子实体萌发情况,统计子实体萌发率。
1.2.2 不同pH值对病原菌菌丝体生长和子实体萌发的影响 PDA培养基灭菌后,在无菌条件下用灭菌的1 mol·L-1HCl和1 mol·L-1NaOH溶液调节PDA培养基酸碱度,设置3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0共9个pH梯度处理。打取直径为6 mm菌丝块接种在不同pH值的PDA平板中央,每处理重复3次,置于30 ℃黑暗条件下恒温条件下培养。72 h后采用十字交叉法测量菌落直径。
由培养30 d的成熟菌落挑取子实体,接种于不同pH的PDA平板上,每皿20粒,4次重复,25 ℃恒温培养,36 h观察子实体萌发情况,统计子实体萌发率。
1.2.3 不同碳源、氮源对病原菌菌丝生长和子实体萌发的影响 以查彼克培养基(2.0 g KNO3,1.0 g KH2PO4,0.5 g KCl,0.5 g MgSO4.7H2O,0.01 g FeSO4,30.0 g蔗糖,1000 mL蒸馏水)为基础,用含有同等质量碳元素的其他碳源(肌醇、半乳糖、葡萄糖、麦芽糖、可溶性淀粉、甘露糖、乳糖、木糖醇)取代培养基中的蔗糖,制成含有不同碳源的培养基,制作平板时每1000 mL培养基加入15 g琼脂。打取直径为6 mm的菌丝块接种在不同碳源的培养基。以不加任何碳源为对照,每处理3个重复,置于30 ℃黑暗条件下恒温培养箱培养。96 h后用十字交叉法测量菌落直径并观察菌体生长情况。
由培养30 d的成熟菌落挑取子实体,接种于不同碳源的PDA平板上培养基,每皿20粒,4次重复,25 ℃恒温培养,36 h观察子实体萌发情况,统计子实体萌发率。
以查彼克培养基为基础,用含有相同质量氮元素的不同氮源(NaNO3、Ca(NO3)2、NH4Cl、酵母膏、尿素、KNO3、蛋白胨)代替其中的KNO3,配成含有不同氮源的液体培养基。打取直径为6 mm的菌丝块接种于含不同氮源平板的中央。以不加任何氮源为对照,每处理3个重复,置于30 ℃黑暗条件下恒温培养箱培养。96 h后用十字交叉法测量菌落直径并观察菌体生长情况。
由培养30 d的成熟菌落挑取子实体,接种于不同氮源的PDA平板上培养基,每皿20粒,4次重复,25 ℃恒温培养,36 h观察子实体萌发情况,统计子实体萌发率。
1.2.4 光照对菌丝生长和子实体萌发的影响 将已接种的PDA培养皿分为3组,作如下处理:(1)30 ℃持续光照培养24 h(光照培养箱);(2)30 ℃持续黑暗培养24 h;(3)30 ℃黑暗12 h与光照12 h交替培养24 h。96 h后采用十字交叉法测量菌落直径。
由培养30 d的成熟菌落挑取子实体,光照时间设定同菌丝,每皿20粒,4次重复,25 ℃恒温培养,36 h观察子实体萌发情况,统计子实体萌发率。
1.2.5 菌丝体与子实体的致死温度测定 菌丝体的致死温度测定:挑取已在液体培养中生长成熟的菌丝块,置于同等大小的已灭菌试管中,加入10 ml无菌水。分别于40、45、50、55、60、65 ℃恒温水浴10 min,用流动水快速冷却试管。取出菌丝接种于PDA平板上30 ℃黑暗条件下培养,2 d后观察并记录菌丝生长情况。每个处理设置4次重复。设未经水浴处理的菌丝块作空白对照。子实体的致死温度测定:收集PDA培养基上萌发初期的成熟子实体。分别置于40、45、50、55、60、65 ℃恒温水浴锅中加热10 min,用冷水迅速冷却试管。将子实体转入PDA平板中,每皿4粒,25 ℃恒温培养。48 h后根据子实体生长情况确定其致死温度。
1.2.6 数据统计与分析 用GraphPad Prism软件分析温度、pH值、不同碳源、氮源和光照时间对病原菌菌丝生长和子实体萌发的影响并作图。用spss19.0比较不同处理的差异显著性。
2 结果与分析
2.1 温度对病原菌菌丝体生长及子实体萌发的影响
图1(A)表明,葡萄座腔菌对温度的适应范围较广。在10、15、20、25、30、35下培养96 h后的菌落直径分别为1.16、3.34、5.72、6.39、7.78、1.63 cm,4 ℃和40 ℃未见菌丝生长,菌落在不同温度的扩展速度不同,适宜温度为20~30 ℃,此温度范围内菌丝生长致密,20 ℃以下或35 ℃以上菌落生长明显趋缓。4 ℃和40 ℃条件下菌落直径无显著性差异(P>0.05),其余各组之间均存在显著性差异(P<0.05)。子实体萌发率测定结果表明,不同温度下子实体萌发差异显著,10~40 ℃范围内子实体均可萌发,4 ℃条件下子实体不能萌发。25 ℃最为适宜,子实体萌发率为78.75%,见图1(B),此温度下子座生长也较为迅速。4 ℃与10 ℃,10 ℃与15 ℃,20 ℃与30 ℃、35 ℃、40 ℃的子实体萌发率无显著性差异(P>0.05),其余不同温度之间的子实体萌发率存在显著性差异(P<0.05)
2.2 不同pH值对病原菌菌丝体生长和子实体萌发的影响
葡萄座腔菌对酸碱度的适应范围较广,在PH 4-10范围内均能生长,PH3和PH11环境下均不能生长,适宜菌落扩展的PH范围为PH4~7,在此范围内菌落生长迅速,菌丝生长致密,长势繁茂。最适P为5,在此条件下生长72 h后的菌落直径为5.27 cm。PH3和PH11、PH4~7、PH9和PH10条件下菌落直径无显著性差异(P>0.05),其余各组之间两两均存在显著性差异(P<0.05)。在PH 3~11范围内子实体均能萌发,碱性条件下更适宜子实体萌发,pH9时子实体萌发率最高,为62.5%,显著高于其他PH值,在PH8~10范围内子实体萌发率较高,见图2。PH3、PH4、PH5、PH11之间,PH6、PH7、PH10之间,PH8、PH10之间的子实体萌发率无显著性差异(P>0.05),其余各不同PH之间的子实体萌发率存在显著性差异(P<0.05)。

图1 温度对菌丝体生长(A)和 子实体萌发(B)的影响

图2 pH对菌丝体生长(A)和子实体萌发(B)的影响
2.3 不同碳源、氮源对病原菌菌丝生长和子实体萌发的影响
图3结果表明,葡萄座腔菌对不同氮源利用差异显著,以可溶性淀粉为碳源时菌丝生长最为迅速,培养96 h之后的菌落直径为7.47 cm,菌落较为致密,与其他碳源处理存在显著差异。其次为蔗糖、甘露糖、葡萄糖、乳糖、麦芽糖,对木糖醇、半乳糖、肌醇的利用较差。以木糖醇为碳源时菌丝生长最慢,菌落稀疏,生长状况差。以蔗糖为碳源时,菌落厚度最为致密(见表1),但菌落生长速度较为缓慢,综合考虑可溶性淀粉为最适碳源。乳糖、葡萄糖、麦芽糖之间,肌醇、木糖醇、半乳糖之间,蔗糖、甘露糖之间菌落直径不存在显著性差异(P>0.05),其余不同炭源之间的菌落直径均存在显著性差异(P<0.05)。

图3 不同碳源对菌丝体生长(图A)和 子实体萌发(图B)的影响
子实体萌发对9种碳源利用差异显著,可溶性淀粉和葡萄糖利用最好,子实体萌发率分别为62.5%、61.25%。其次为蔗糖、麦芽糖、半乳糖、木糖醇。乳糖、肌醇、甘露糖对子实体萌发的效果较差。乳糖、肌醇、甘露糖、空白之间,蔗糖、木糖醇、半乳糖、麦芽糖之间,葡萄糖、淀粉之间的子实体萌发率无显著性差异(P>0.05),其余各碳源之间的子实体萌发率均存在显著性差异(P<0.05)。
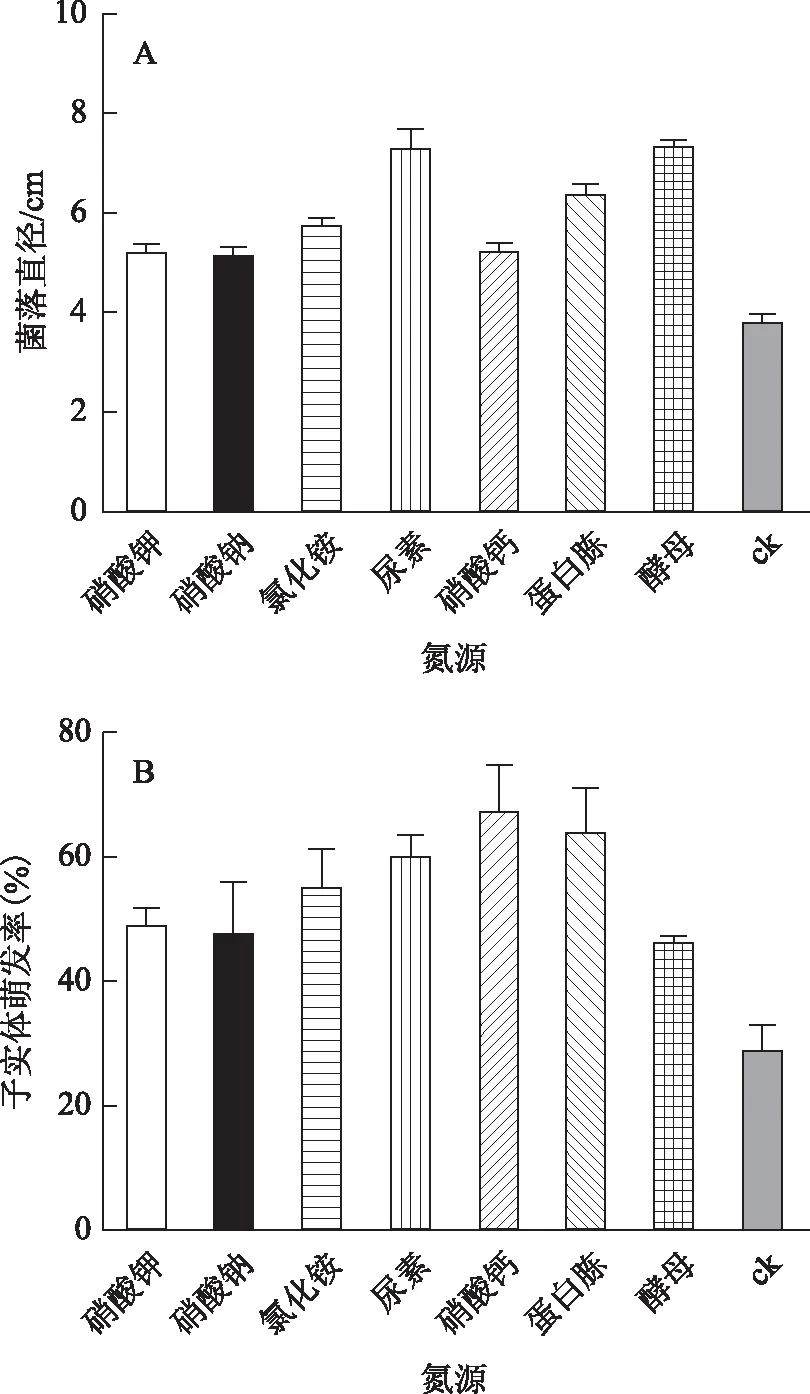
图4 不同氮源对菌丝体生长(图A)和 子实体萌发(图B)的影响
葡萄座腔菌对供试的氮源均能利用,但对不同氮源的利用率存在显著差异。图4结果显示,供试菌株在尿素和酵母膏作为氮源的培养基生长速度最快,培养96 h之后的菌落直径分别为7.32 cm和7.34 cm,硝酸钾、硝酸钠、氯化铵、硝酸钙之间,尿素、酵母膏,氯化铵、蛋白胨之间菌落直径无显著差异(P>0.05),其余各氮源之间菌落直径存在显著性差异(P<0.05)。以酵母膏作为氮源的菌落更为致密,因此酵母膏为最适氮源,其次为尿素。无氮源存在时菌落厚度很小。以硝酸钙作为氮源时子实体萌发率最高,达到67.5%。其次为蛋白胨,子实体萌发率可达到63.75%,硝酸钠作为氮源时子实体的萌发率最低,只有47.5%。空白组与其他各氮源之间的子实体萌发率均存在显著性差异(P<0.05)。硝酸钙与硝酸钾、硝酸钠、酵母,蛋白胨与酵母之间的子实体萌发率存在显著性差异(P<0.05),其余各氮源之间的子实体萌发率无显著性差异(P>0.05)。

表1 不同碳、氮源对菌丝生长的影响
注:“+”表示菌落生长稀薄,菌落厚度依次有“+”、“++”、 “+++”。
2.4 光照对菌丝生长和子实体萌发的影响
试验结果表明,不同光照时间对菌丝生长的影响不明显,黑暗条件菌丝生长速度略高于光暗交替和全光照条件。黑暗条件和光暗交替处理的菌落直径无显著性差异(P>0.05),其余各组之间存在显著性差异(P<0.05)。从子实体萌发率来看,黑暗条件最优,光照条件和光暗交替子实体萌发率均显著降低,分别为23.75%和35%,见图5。不同光照时间的子实体萌发率之间均存在显著性差异(P<0.05)。
2.5 菌丝体与子实体的致死温度
结果表明,经40~55 ℃处理10 min之后的子实体和菌丝均能正常生长,菌丝60 ℃水浴10 min后完全致死,经60 ℃水浴10 min之后的子实体接种到PDA培养基之后不再生长,故子实体和菌丝的致死温度均为60 ℃水浴10 min。
3 结果与讨论
目前有关栀子褐斑病病原菌生物学特性方面的系统资料相对较少,因此,进行病原菌的生物学特性试验有重要的意义。本试验结果表明,葡萄座腔菌可不同程度的利用多种碳源和氮源,最适宜子实体萌发的碳源和氮源分别为淀粉和硝酸钙,最适宜

图5 不同光照时间对菌丝体生长(图A)和 子实体萌发(图B)的影响

处理温度(℃)菌丝生长情况子实体生长情况ck++++++++40℃++++++++45℃++++++++50℃++++++++55℃++++++++60℃--------65℃--------
注:“+”表示生长;“-”表示不生长。
菌丝生长的碳源和氮源分别是可溶性淀粉和酵母膏,施用过多氮肥既会造成草坪草徒长,又会加剧根腐病的发生,因此,少施氮肥对控制该病害的发生有显著作用。光照试验可以得出,黑暗条件有利于病害的蔓延,光照条件病害发生较慢。病原菌在10~35 ℃均能生长,20~30 ℃为菌丝生长的适宜范围,与董一丹[7]报道的褐斑病通常在较温暖(>25 ℃)、湿度较大的夏天发生相一致。菌丝和子实体的致死温度均为60 ℃处理10min,可能原因为其寄主及生长环境不同或是菌群分化等原因,使得病原菌某些生物学特性也具有一定的差异,尚待深入研究[8]。
葡萄座腔菌生长的最适pH为5,说明偏酸性土壤上的草坪发病重于碱性土壤,改善土壤酸碱度可以有效预防该病害的发生。
在生产上应采取综合的管理和防治措施,病害发生前后,积极做好预防和防治工作等,将病害发生程度降低到最低水平。本试验的研究结果对于有针对性地拟定病害防治措施具有一定的参考意义。
[1] 刘益华,李晶,林曼婷,等.栀子有效成分栀子苷的现代研究进展[J].中国药学杂志.2012,47(6):406-409.
[2] 颜升,董艳凯,陈健,等.中药栀子的研究现状[J].安徽农业科学.2013,41(18):7759-7760,7772.
[3] 李国元,邓青云.栀子主要病害发生特点及综合防治[J].湖北林业科技.2003(3):24-26.
[4] 姜晓龙,金磊磊,陈辉辉,等.1株侵染蓖麻的葡萄座腔菌菌株鉴定[J].江苏农业科学.2015,43(2):319-322.
[5] 李冬,刘兴平.栀子病害调查及病原鉴定初报[J].江西林业科技.2009,(5):31-32.
[6] 古丽君,徐秉良,李彬,等.草坪禾草根腐病病原菌生物学特性研究[J].草业学报,2012,21(3):93-98.
[7] 董一丹.高羊茅褐斑病的病原学及防治技术研究[D].武汉:华中农工业大学.2006.
[8] 陈海波.草坪草褐斑病病原鉴定及化学防治研究[D].兰州:甘肃农业大学.2001.
StudyonBiologicalCharacteristicsGardeniaLeafSpot
LUO Guangming,DONG Yankai,ZHU Yuye*,HU Yanzhen,GONG Yuhong,WANG Xiaoyun
(JiangxiUniversityoftraditionalChinesemedicinecollegeofpharmacy,Nanchang330004,China)
Leaf spot lesions pathogens were isolated from gardenia plants,which were identified asBotryosphaeriadothidea.The experiment studied the biological characteristics of pathogens,including different temperature,illumination time,pH,carbon and nitrogen source for mycelial growth and sclerotia nutrition germination and determine the mycelium and sclerotia lethal temperature.The results showed that mycelial growth temperature ranged from 10~35 ℃,the optimum growth temperature was 30 ℃,the mycelium could grow within pH 4 to pH 10,the optimum pH value was pH 5.Suitable carbon and nitrogen sources for mycelial growth were soluble starch and yeast extract.Alternating light and dark condition was more suitable for mycelial growth.Sclerotia could germinate at the range of 15 ℃ to 40 ℃,and the optimum temperature was 25 ℃.In sclerotia could germinate at the range of pH 3 to pH 11,and the optimum pH value is pH 9.Soluble starch and calcium nitrate were the optimum carbon and nitrogen sources for the germination of sclerotia.Under dark conditions it was suitable for military and germination.The lethal temperatures of sclerotia and mycelium are 60 ℃.
Gardenia;biological characteristics;leaf spot
10.13313/j.issn.1673-4890.2016.10.016
2015-10-22)
国家公益性行业科研专项资助项目(201507002)。
*
朱玉野,讲师,研究方向:药用植物;E-mail:zyy711@126.com
