黄山殷溪河光唇鱼食性特征的初步研究
2016-09-23刘雪飞余银姣王梦田王智怡严云志
刘雪飞,汪 晗,余银姣,王梦田,王智怡,严云志
(安徽师范大学生命科学学院,安徽芜湖 241000)
黄山殷溪河光唇鱼食性特征的初步研究
刘雪飞,汪晗,余银姣,王梦田,王智怡,严云志
(安徽师范大学生命科学学院,安徽芜湖241000)
基于2014年7月于黄山殷溪河采集的94尾光唇鱼(Acrossocheilusfasciatus)标本,初步研究了该物种的食性特点,分析了其食物组成及其多样性随年龄和全长增长的变化。渔获物的个体全长为17.93~357.72 mm,分为5个年龄组。光唇鱼的食物由陆生和水生植物、陆生昆虫、碎屑以及环节动物、软体动物、甲壳动物、水生昆虫等水生动物组成,其中以水生昆虫、水生植物和碎屑为常见和优势饵料种类。单因素相似性分析结果显示,1~3龄组间的食物类型数无显著差异,但食物多样性指数和宽度指数均存在显著差异。Pearson′s相关分析结果显示,随个体全长增大,食物类型数显著增多而宽度指数显著下降,但食物多样性无显著变化。此外,冗余分析结果显示,光唇鱼的食物组成与个体全长显著相关;随个体全长增大,胃含物中水生昆虫类逐渐减少而甲壳类和陆生昆虫逐渐增多,但水生植物和碎屑的多度与全长无明显关系。
光唇鱼(Acrossocheilusfasciatus);食物组成;食物多样性;殷溪河
光唇鱼(Acrossocheilusfasciatus),隶属鲤形目(Cypriniformes)鲤科(Cyprinidae)鲃亚科(Barbinae)光唇鱼属(Acrossocheilus),是我国特有的小型鱼类,具有一定的食用和观赏价值。主要分布在安徽、福建和浙江等地的山区急流水域,是山区渔业经济的主要品种之一[1]。因其成鱼常栖息于石缝间、体侧具有6条鲜艳的黑色横斑而被地方称为“石斑鱼”[2]。
在安徽省的皖南山区,光唇鱼分布于海拔较高、水流湍急、底质粗糙(如圆石、鹅卵石等)的中小型河流及其支流,是该水域的常见种和优势种[3,4]。近年来,受城镇化发展、河道片段化、水质污染等人类活动的影响,野生光唇鱼资源量日益下降,分布范围逐渐减小,并呈现出明显的低龄化发展趋势[3,4]。有关皖南山区光唇鱼的年龄、生长和繁殖等生物学特征及其环境适应性研究已有报道[4-6],但有关其食性研究尚无报道。
殷溪河是皖南山区青弋江流域的一条河源溪流,发源于中国安徽省黄山市东部的九龙峰自然保护区内。本研究根据2014年7月采集的殷溪河光唇鱼标本,研究了其食性特点,分析了食物组成及其多样性同年龄和个体全长之间的关系,以期为该物种的保护及其资源开发利用提供基础资料。
1 材料与方法
1.1标本采集
2014年7月,以电鱼器采集殷溪河光唇鱼标本94尾;现场用8%福尔马林溶液浸泡后带回实验室用于后续研究。
1.2食性分析
取前肠(咽部至肠道的第一弯曲部),在显微镜下对其内含物进行定性和定量分析。食物类型分为以下5大类:陆生植物、水生植物、陆生动物、水生动物和碎屑;对于水生动物而言,进一步分为4类:水生昆虫类、软体动物类、甲壳类、环节动物。使用压片体积法进行食物的定量分析[7-8]。
1.3年龄估算
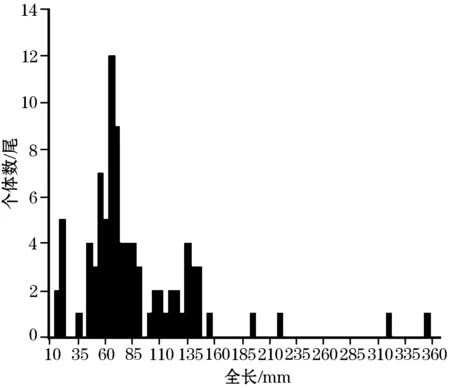
运用长度分布频率法对光唇鱼的年龄组成进行估算;该方法适用于具有寿命短、生长快等生活史特征的小型鱼类的年龄估算[9],已被运用于本研究区域内光唇鱼[10]、尖头鱥(Phoxinusoxycephalus)[11]等鱼类的年龄估算。我们测量了每尾标本的个体全长(精确到0.01 mm),基于测量数据,按每间隔5.0 mm的全长范围内的标本数量构建其长度频率分布柱状图,根据其频率分布的波峰数量来确定其年龄组及其对应的全长范围,进而确定每尾标本的年龄。
1.4数据分析
根据每食物类型在89尾光唇鱼标本(另外的5尾标本空肠)的前肠中的出现次数及相对体积大小,计算每食物类型的出现频率(F,%)、相对多度(P,%)和重要值指数(IVI),以及每尾光唇鱼的食物多样性指数(Shannon index,H′)[12]和食物宽度指数(Levin′s standardized index,B)[13],计算公式如下:
Fi=100×(Si/S);
Pi=100×(Vi/V);
IVIi=Fi×Pi;
H′=-∑(Pi× log2Pi);
B=[(∑Pi2)-1-1] × (n-1)-1。
上式中,Fi、Pi和IVIi分别代表食物类型i的出现频率、相对体积大小和重要值指数,Si和Vi分别代表食物类型i的出现次数(标本数)和体积大小,S和V分别为全部光唇鱼标本数和全部食物的体积大小。
1.5数据统计
鉴于4龄和5龄鱼均仅2尾标本,因此只对1~3龄鱼的食性进行分析。另外,将食物体积大小的数据经log10(x+1)转化后用于分析以降低极端数据的负面权重。
以SPSS 13.0软件包为工具,运用单因素方差分析(One-way ANOVA)分别检验1~3龄组间的食物类型数、多样性指数及其宽度指数的差异;若存在显著差异,再运用post-hoc多重比较进行两两年龄组间差异比较。运用Pearson′s相关分析分别检验食物类型数、多样性指数及宽度指数与个体全长的关系,视P<0.05为差异显著性。
以PRIMER 5.0软件包为工具,运用单因素相似性分析(One-way ANOSIM)检验1~3龄组间食物组成(以各食物类型的相对体积大小来反映)的差异情况,视P<0.05为差异显著,视R值大小判别分离程度:R>0.75,完全分离;0.5 2.1渔获物个体概况 采集的94尾光唇鱼的全长范围为17.93~357.72 mm,体重为0.03~311.08 g。94尾标本中,5尾标本的前肠为空肠,89尾前肠存在内含物。根据其全长频率分布柱状图,89尾有效标本可分为5个年龄组,1~5龄组的全长依次为17.93~22.47 mm(7尾)、39.96~94.80 mm(56尾)、101.67~157.98 mm(22尾)、199.11~223.05 mm(2尾)、321.06~357.72 mm(2尾)(图1)。 图1 光唇鱼渔获物的全长频率分布柱状图Fig.1 Histograms in total length frequency for A.fasciatus 2.2食物组成概况 光唇鱼的食物类型由陆生植物、水生植物、陆生昆虫、碎屑与水生动物(含环节动物、软体动物、甲壳动物、水生昆虫)组成,其中陆生动物均为昆虫类。水生昆虫的出现频率和相对多度均最大,其重要值指数达3954.9,为光唇鱼的优势饵料种类;而水生植物和碎屑的重要值指数,分列第2、3位。陆生昆虫的出现频率较高(>20%),但相对多度较低(<2%),因而重要值指数偏低。此外,陆生植物、环节动物和软体动物均仅见于2尾标本的前肠内含物,甲壳动物仅见于5尾标本前肠中,它们的出现频次均低于10%,同属光唇鱼的非优势饵料种类(表1)。 2.3食物多样性同年龄和全长的关系 运用单因素方差分析(One-way ANOVA)检验了1~3龄组光唇鱼之间的食物类型数、多样性指数及其宽度指数的差异情况,结果显示,1~3龄组间的食物类型数无显著差异(F=2.41,P>0.05),但多样性指数(F=4.90,P<0.05)和宽度指数(F=6.13,P<0.01)均存在显著差异。3龄组的食物多样性指数和宽度指数均低于1龄组和2龄组(表2)。 表1 光唇鱼的食物组成及其出现频率和重量值指数Tab.1 Diet compositions and their frequency of occurrence and importance value index of A.fasciatus 表2 光唇鱼1~3龄组之间的食物多样性变化Tab.2 Variations in diet diversity among different age-groups of A.fasciatus 运用Pearson′s相关分析检验了食物类型数、多样性指数及宽度指数与个体全长的关系,结果显示,随个体全长的增长,食物类型数显著增多(R=0.29,P<0.01),食物宽度指数显著下降(R=-0.36,P<0.01),但食物多样性指数无显著变化(R=-0.16,P>0.05)。考虑到全长同食物类型数和宽度指数之间的相关性,因而计算出其关系式如下:lgND=0.14×lgTL+0.26(R=0.29,n=89);lgB=-0.17×lgTL+0.46(R=-0.36,n=89)。 2.4食物组成同年龄和全长的关系 根据单因素相似性分析(One-way ANOSIM),1~3龄组间的食物组成存在明显分离且差异显著(GlobalR=0.49,P<0.01)。相似性百分比分析(SIMPER)显示,水生植物和水生昆虫的多度变化是造成1~3龄组间食物组成差异的主要贡献食物类型,其贡献值分别大于30%,这两类食物类型在2龄组中的平均多度最高,3龄组次之,1龄组最低。碎屑的多度变化对1~3龄组间食物组成差异的贡献值大于10%,而陆生昆虫和甲壳类的贡献率少于5%,而这三类食物类型仅在2龄组和3龄组中出现,且在3龄组中具有更高的平均多度(表3)。 表3 各食物类型的平均多度(AA)及其对食物组成差异的贡献率(Con)和不相似性(AD)Tab.3 Average abundance of each diet group and their contribution to the dissimilarity in diet composition among 1~3 aging groups of A.fasciatus 运用冗余分析(RDA)检验了光唇鱼食物组成同个体全长的关系,结果显示两者间存在显著性相关(P<0.05),RDA的四个轴共解释了90.4%的食物组成变异。根据RDA双标图,随光唇鱼个体全长的增长,其食物中水生昆虫类逐渐减少、甲壳类和陆生昆虫却逐渐增多,但水生植物和碎屑的多度与全长无明显关系(图2)。 图2 食物组成随个体全长变化的冗余分析双标图Fig.2 RDA biplot examining the correlation between diet composition and total length of A.fasciatus 本研究采用长度频率分布的方法对采集的光唇鱼年龄组进行了划分,结果表明,89尾光唇鱼共分为5个年龄组。运用长度分布频率法划分年龄组,具有操作快捷、可不处死鱼等优点。由于长度频率无法显示长度几乎停止增长的高龄个体的真实年龄[14],该方法虽不适用于年龄结构复杂及个体生长缓慢的鱼类种群的年龄鉴定,但对于年龄结构简单、生长快速或年龄结构复杂但处于生命早期阶段的鱼类种群而言,该方法具有很好的适用性[9]。郭丽丽[10]曾对黄山浦溪河光唇鱼的年龄和生长进行了研究,基于其研究结果,在5—7月份,浦溪河光唇鱼分为4个年龄组,其全长范围分别为0~25 mm(1龄)、25~75 mm(2龄)、75~120 mm(3龄)、120~155 mm(4龄)。本研究区域殷溪河与浦溪河同处于黄山北麓、同为青弋江流域的河源溪流、海拔相近且地理位置接近等,这两条溪流中光唇鱼很可能具有相似的个体生态学特征(如种群年龄结构、生长速度等)。在本研究结果中,殷溪河光唇鱼5个年龄组的全长分布范围(15~25 mm,1龄;25~95 mm,2龄;100~160 mm,3龄;195~225 mm,4龄;320~360 mm,5龄)与郭丽丽[10]的研究结果基本一致,但整体上,各年龄段的个体全长阈值范围稍大于浦溪河光唇鱼,这可能因为郭丽丽[10]是对浦溪河5—7月份的光唇鱼渔获物进行整体分析,而本研究仅针对殷溪河7月份渔获物分析,考虑到5—7月份的时间差,该差异也就不难理解。 本研究表明,殷溪河光唇鱼的食谱较广,主要摄食水生昆虫和水生植物(主要为固着藻类)和部分碎屑,这是其作为皖南山区溪流优势种的适应性结果。光唇鱼主要分布的山区中小型河流及其支流,水流湍急,水温较低,底质粗糙,水体初级生产力较低,而陆地植被盖度较大,雨水将陆地生态系统的营养带入河流中。因此,植物碎屑、以碎屑为食的水生昆虫和水生植物(主要为固着藻类)构成了鱼类主要的食物来源。鱼类对食物的选择由其对饵料的喜好性和饵料的易得性共同决定。在光唇鱼分布的水体中,植物碎屑为最易得饵料,但在其食物组成中,营养较高的水生昆虫和水生植物构成了主体,其比例要远大于植物碎屑,表明光唇鱼较强的摄食能力,体现了其对周围环境很好的适应性。本研究结果与朱仁等[8]、陈凌云等[1]的报道一致。 鱼类的食性往往存在物种间及个体间的差异,食物组成随着水域、季节等时空因素的变化而变化;此外,在鱼类个体的生长过程中,不同大小或不同年龄段的个体对饵料生物资源的利用也存在差异[15-17]。已有研究表明,对于鱼类来说,随着个体体长增加,其口裂增大、摄食能力不断加强,其摄食饵料生物的种类、数量和大小也发生变化,甚至会出现较为明显的食性转化[18-19]。在本研究中,殷溪河光唇鱼各个体长组的食物主要组成均为水生植物和水生昆虫,未发现明显的食性转化。这可能是由于山区溪流水体清急,适合作为早期生长饵料的浮游生物、较大个体饵料的软体动物等不易生长,鱼类饵料来源相对单一。另一方面,随体长的增长,殷溪河光唇鱼的食物类型数显著上升而宽度指数显著下降。食物类型数的上升,表明了光唇鱼摄食能力的不断增强,可以摄食更多类型的食物;宽度指数显著下降,则表明其对饵料的选择性增强。这种选择性的增强,有可能是其对食物的偏好,也有可能是食物竞争所致。随着体长的增加,种群的食物需求增加,与光唇鱼同域分布的宽鳍鱲、麦穗鱼等鱼类的食物组成与光唇鱼有着很高的重叠。就食物组成而言,随鱼体增长,光唇鱼饵料生物中的水生昆虫数量减少但陆生昆虫数量增多这也表明了其摄食能力的不断提高。 本研究的溪流为殷溪河——皖南山区青弋江流域的一条河源溪流,这些溪流的物理栖息地也可能存在较高的季节动荡,考虑到不同时期光唇鱼的食性特征由于外界环境的不同,而具有潜在的季节变化和空间差异,今后有必要进一步深入研究其食性随季节和空间变化的一些特征。 [1]陈凌云,郭水荣,应利萍,等.光唇鱼山区池塘流水养殖试验[J].水产养殖,2014,(6):37-38. [2]陈宜瑜.中国动物志 硬骨鱼纲 鲤形目 (中卷)[M].北京:科学出版社,1998. [3]郭丽丽,严云志,席贻龙.黄山浦溪河光唇鱼的性腺发育周年变化[J].淡水渔业,2008,38(6):8-13. [4]Yan Y Z,Guo L L,Xiang X Y,et al.Age and growth ofAcrossocheilusfasciatus(Barbinae,Cyprinidae) from the Puxi Stream in the Huangshan Mountain[J].China J Freshwat Ecol,2010,25 (1):79-83. [5]Yan Y Z,Guo L L,Xiang X Y,et al.Breeding strategy ofAcrossocheilusfasciatusin the Puxi Stream of the Huangshan Mountain[J].Curr Zool,2009,55 (5):350-356. [6]Yan Y Z,Zhu R,He S,et al.Life-history strategies ofAcrossocheilusfasciatus(Barbinae,Cyprinidae) in the Huishui Stream of the Qingyi watershed,China[J].Ichthyol Res,2012,59(3):202-211. [7]Güntzel A M, Melo I K M, Roche K F, et al. Cladocerans from gut contents of fishes associated to macrophytes from Taquari River Basin,MS,Brazil[J].Acta Limnol Brasilien,2012,24 (1):97-102. [8]朱仁,严云志,孙建建,等.黄山陈村水库河源溪流鱼类群落的食性[J].生态学杂志,2012,31 (2):359-366. [9]Casselman J M.Determination of age and growth//Weatherley A H.The biology of fish growth[M].New York:Academic Press,1987:209-242. [10]郭丽丽.黄山浦溪河光唇鱼的年龄、生长和繁殖的研究[D].合肥:安徽师范大学,2009. [11]Chu L,Ye J,Si C,et al.Age,growth and reproduction of the Chinese minnow,Phoxinusoxycephalusin the Jiulongfeng Nature Reserve of the Huangshan Mountain[J].Acta Hydrobiol Sinic,2015,39(1):29-37. [12]Hulbert S H.The nonconcept of species diversity:A critique and alternative parameters[J].Ecology ,1971,52(4) :577-586. [13]Lessells M C,Krebs R J.Age and breeding performance of European bee-eaters[J].Auk ,1989 ,106 (3):375-382. [14]Albanese B.Life history of the flagfin shiner,Pteronotropissingnipinnis,from a black-water stream in southeastern Mississippi[J].Ecol Freshwat Fish,2000,9(4):219-228. [15]Werner E E,Gilliam J F,Hall D J,et al.An experimental test of the effects of predation risk on habitat use in fish[J].Ecology,2000,64(6):1540-1548. [16]Wootton R J.Ecology of Teleost Fishes[M].London:Chapman and Hall,1990. [17]殷名称.鱼类早期生活史研究与其进展[J].水产学报,1991,15 (4):348-358. [18]刘焕亮,丁守河,杨云龙,等.尼罗罗非鱼摄食器官胚后发育生物学[J].水产学报,1994,18 (1):8-17. [19]Guillemaud T,Streiff R,Santos R S,et al.Microsatellite characterization in the rainbow wrasseCorisjulis(Pisces:Labridae)[J].Mol Ecol,2000,9(5):631-632. (责任编辑:张红林) The primary research of the feeding habits of Acrossocheilus fasciatus in the Yinxi Stream of the Huangshan mountain LIU Xue-fei ,WANG Han ,YU Yin-jiao,WANG Meng-tian,WANG Zhi-yi,YAN Yun-zhi (CollegeofLifeSciences,AnhuiNormalUniversity,Wuhu241000,Anhui,China) Based on the specimens ofAcrossocheilusfasciatuscollected from the Yinxi stream of the Huangshan mountain during July 2014,the feeding habits of this species and the pattern of how diet composition and diversity relate to ages and total-lengths were examined in this study.A total of 94 specimens were collected,ranging from 17.93 mm to 357.72 mm in total length and comprising five age groups.The diet ofA.fasciatuswere mainly composed of eight diet-categories,including terrestrial plant,aquatic plant,terrestrial insect,detritus,aquatic annelid,mollusk,crustacean and aquatic insect,among which the most common and dominant diet-categories involved aquatic plant,aquatic insect and detritus.According to the results of One-way ANOVA,H- and B-indices differed significantly across 1~3 age groups,but the number of diet-categories did not show this significant among-age variation.The results of Pearson′s correlation analysis showed that total length significantly related positively to number of diet-categories but negatively to B-index;however,no significant correlation was observed between total length and H-index.In addition,based on the RDA results,diet composition had significant correlation with total length.Aquatic insects increased but crustacean and terrestrial insect decreased with total length increasing.However,aquatic plant and detritus showed no obvious relation to total length. Acrossocheilusfasciatus;diet composition;diet diversity;the Yinxi Stream 2015-11-06; 2016-05-20 国家级大学生创新创业训练计划(201410370049);国家级星火计划重大项目(2013GA710001) 刘雪飞(1994-),女,专业为生物科学。E-mail:Liuxuefei366@163.com 严云志。E-mail:yanyunzhi7677@126.com S931.1 A 1000-6907-(2016)05-0025-052 结果


3 讨论
