亚损伤状态下枯草芽孢杆菌芽胞萌芽影响因素的研究
2016-09-14伍金娥武汉轻工大学食品科学与工程学院湖北武汉430023
常 超,潘 丽,伍金娥,张 喆(武汉轻工大学食品科学与工程学院,湖北武汉430023)
亚损伤状态下枯草芽孢杆菌芽胞萌芽影响因素的研究
常超,潘丽,伍金娥,张喆
(武汉轻工大学食品科学与工程学院,湖北武汉430023)
为了研究食品中营养要素对处于亚损伤状态枯草芽孢杆菌芽胞的影响,采用分批培养法,利用食品中存在的碳源、氮源、无机盐离子等分别诱导芽胞萌芽,采用正交实验研究了温度、pH、接种量、溶氧量对芽胞萌芽率的影响,分别在OD580下测定其光密度值,得到芽胞萌芽率。结果显示,麦芽糖、乳清蛋白是芽胞萌芽的最佳碳源、氮源;无机盐MgSO4对芽胞萌芽产生显著的诱导作用。以麦芽糖(5 mg/mL),MgSO4(5 mg/mL),乳清蛋白(5 mg/mL)为基础培养基,芽胞的最佳生长条件为:接种量4%、pH7.0、溶氧量70 mL,37℃恒温振荡培养60 min,得芽胞萌芽率最大为15.27%。
枯草芽孢杆菌,亚损伤状态,萌芽,影响因素
枯草芽孢杆菌(Bacillus subtilis)具有代谢旺盛,能分泌脂肪酶、蛋白酶、淀粉酶等,广泛分布于空气、水体、土壤、生物体的体表等,同时也是污染食品的最常见的芽胞菌之一[1]。在寡营养、缺氧、辐照等条件下能产生抗逆性很强的内源芽胞,赋予其具有耐高温、抗干燥、抗辐射以及某些有机溶剂等多种类型的抗逆性特征[2]。亚致死损伤发生在细菌或芽胞暴露在不利的物理化学环境(超出生长范围,但不在致死的范围)后,导致了细胞的功能和结构组织发生了可逆的变化。食品加工中重要手段如低热杀菌、低水分活度、辐射、电脉冲杀菌技术、低pH、加入防腐剂等均可能导致亚致死损伤发生[3]。然而,受损的菌体或芽胞能够在适宜的环境中修复损伤,从而导致食品的污染。一旦枯草芽孢杆菌污染食品会使食品的品质发生变化,如面粉中存在枯草芽孢杆菌,不仅可使发酵的面团液化并伴有黏丝状物质,还会使烤制的面包或饼干的表面出现斑状形态,且产生不良气味;污染肉类,可在其表面可产生黏液并有异味等[2,4]。因此,本研究选择影响枯草芽孢杆菌受损芽胞(80℃下处理30 min)的萌芽因素,深入探讨食品中诱导枯草芽孢杆菌芽胞萌芽的主要因素以及芽胞萌芽的条件,为有效控制和预防枯草芽孢杆菌芽胞对食品品质的影响提供理论依据。由于影响微生物生长的外界因素主要有两大方面,其一为营养物质,其二是许多物理、化学因素,因此本文从这两方面探讨对受损芽孢萌芽影响的作用规律。
1 材料与方法
1.1材料与仪器
菌株枯草芽孢杆菌为武汉轻工大学微生物实验室提供;牛肉膏,蛋白胨,葡萄糖,氯化钠,酵母粉,蔗糖,乳糖,麦芽糖,可溶性淀粉,卵清蛋白,乳清蛋白,牛血清蛋白,硫酸镁(MgSO4·7H2O分析纯),硫酸锰(MnSO4分析纯),氯化钾(KCl分析纯),氯化钙(CaCl2分析纯);10 mmol/L Tris-HCl缓冲溶液称取0.605 g Tris-base,加490 mL蒸馏水溶解后,用5 mol/L HCl和5 mol/L NaOH调节pH为8,然后再定容至500 mL,于121℃高压灭菌20 min,备用。
AL204梅特勒电子分析天平瑞士梅特勒-托利多公司;YM-50全自动灭菌锅上海三申公司;SW-CJ-2FD型超净工作台苏州净化设备有限公司;GHP-9050隔水式恒温箱、电热恒温鼓风干燥箱上海齐欣科学仪器有限公司;PHS-3C精密pH计上海雷磁仪器厂;UPK-I-10T优普系列超纯水器西安优普仪器设备有限公司;A011754紫外可见分光光度计上海现科分光仪器有限公司。
1.2实验方法
1.2.1菌种的处理
1.2.1.1菌种活化培养基牛肉膏5 g,蛋白胨10 g,葡萄糖5 g,氯化钠5 g,水1000 mL,pH7.2;菌种扩大培养基:蛋白胨10 g,酵母粉5 g,氯化钠10 g,水1000 mL,pH7.0。
1.2.1.2菌种活化配制菌种活化培养基后分装,取250 mL锥形瓶装50 mL的培养液,进行121℃高压灭菌15 min后冷却,将枯草芽孢杆菌接种,置于37℃振荡培养箱中培养1 d,进行复苏活化处理。
1.2.1.3菌种扩大培养将菌种活化液以2%的接种量接种至菌种扩大培养基中,置于37℃振荡培养箱中培养72 h。
1.2.1.4制备芽胞液将扩大培养物以6000 r/min,离心10 min。所得菌悬液用无菌蒸馏水洗涤并离心3~4次,再将新的悬浮液加入5 mL的无菌蒸馏水,并在100℃下处理30 min杀死其中的营养细胞,得到的芽胞悬液如上述离心并洗涤4次。最后于4℃下储存芽胞悬液,浓度约为109cfu/mL。
1.2.2芽胞萌芽率的测定萌芽率的测定参考Chedia等[5]的方法进行测定,具体如下:将制备的芽胞悬液稀释成工作液1×106cfu/mL,再次放入80℃的水浴锅中30 min,使芽胞处于亚损伤状态,以2%的接种量将芽胞液接种至含有10 mmol/L Tris-HCl缓冲溶液的250 mL锥形瓶中,每瓶装液量为50 mL,分别添加不同碳源、氮源、无机盐离子。接种后先使用可见分光光计测定0 min的光密度值,然后再将其移至37℃的恒温振荡培养箱中培养60 min,再次测定光密度值。萌芽率表示为萌芽60 min后样液的OD580值与初始值的比值,计算公式如下:式中:ODi:芽胞萌芽0 min时的OD值,即样品的初始OD580值;ODf:芽胞萌芽60 min时的OD值,即试样孵育60 min后的OD580值。
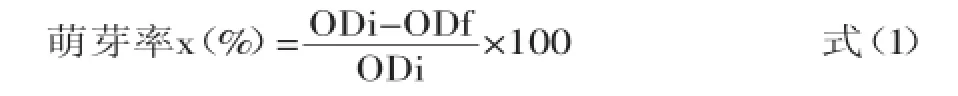
1.2.3营养元素对枯草芽孢杆菌芽胞萌芽的影响
1.2.3.1不同碳源诱导枯草芽孢杆菌芽胞的萌芽将
1.2.2所述受损芽胞添加至50 mL 10 mmol/L Tris-HCl缓冲溶液中,其中氮源为0.1%蛋白胨、无机盐为2 mmol/L NaCl,分别添加麦芽糖、葡萄糖、乳糖、蔗糖、可溶性淀粉,终浓度为5 mg/mL。于37℃培养60 min,测定光密度值的变化,按1.2.2计算萌芽率。
1.2.3.2不同氮源诱导枯草芽孢杆菌芽胞的萌芽将
1.2.2所述受损芽胞添加至50 mL 10 mmol/L Tris-HCl缓冲溶液中,其中碳源为0.1%葡萄糖、无机盐为2 mmol/L NaCl,分别添加牛血清蛋白、乳清蛋白、卵清蛋白、蛋白胨和牛肉膏,终浓度为5 mg/mL。37℃培养60 min,测定光密度值的变化,按1.2.2计算萌芽率。
1.2.3.3不同无机盐离子诱导枯草芽孢杆菌芽胞的萌芽将1.2.2所述受损芽胞添加至50 mL 10 mmol/L Tris-HCl缓冲溶液中,其中碳源为0.1%葡萄糖,氮源为0.1%蛋白胨,分别添加MnSO4、NaCl、CaCl2、MgSO4和KCl,终浓度为5 mg/mL。37℃培养60 min,测定光密度值的变化,按1.2.2计算萌芽率。
1.2.4培养环境条件因素的影响由于温度、pH、接种量、溶氧量对微生物生长的具有直接的影响[2-3],将
1.2.2所述受损芽胞添加至50 mL 10 mmol/L Tris-HCl缓冲溶液中,37℃放置15 min,按表1列出的接种量接种至麦芽糖(5 mg/mL),MgSO4(5 mg/mL),乳清蛋白(5 mg/mL)的基础培养基中,设置温度、pH、接种量、溶氧量各三个水平[6],进行L9(34)正交实验。实验设计见表1,以进一步确定芽胞萌芽的最适条件。按式(1)计算萌芽率。
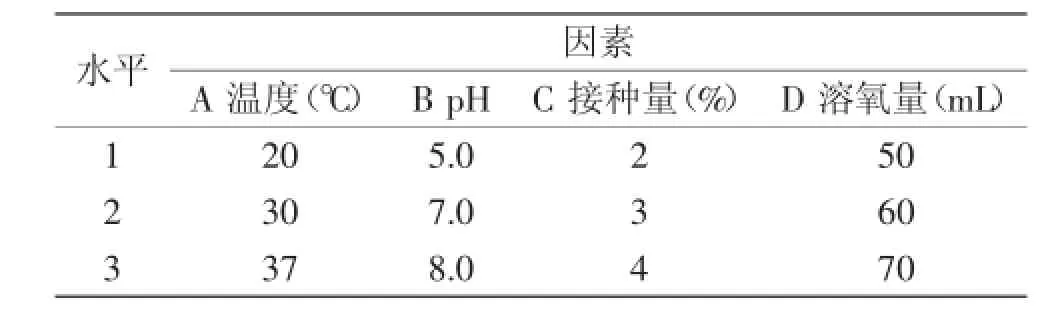
表1 正交实验因素水平表Table 1 Factors and levels of orthogonal experimental design
1.3统计学分析
利用SPSS 17.0软件对所得数据进行统计学分析,实验重复三次,结果均为平均值±标准差表示。
2 结果与分析
2.1营养元素对芽孢萌芽的影响
2.1.1不同碳源因子对芽胞萌芽率的影响从图1可知,枯草芽孢杆菌处于亚损伤状态下的芽胞在不同碳源诱导下其萌芽率表现不同。利用SPSS软件进行多重方差分析可以看出葡萄糖、麦芽糖和乳糖对芽孢萌芽的影响呈显著性差异,而蔗糖与可溶性淀粉之间则无显著性差异。麦芽糖和葡萄糖对芽孢的萌芽率影响高于乳糖,蔗糖和可溶性淀粉对芽胞萌芽率的影响基本一致。
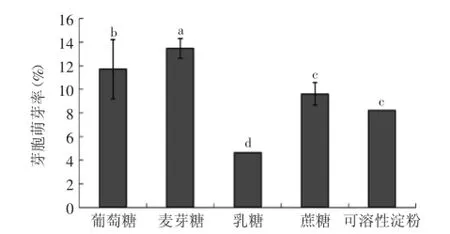
图1 不同碳源对芽胞萌芽率的影响Fig.1 Effects of different carbon sources on spore germination rates
对于芽胞的萌芽,尽管研究表明,芽胞处于80~85℃、5 min可促进芽胞的萌芽,而本实验中采用的条件为80℃、30 min,为一种亚致死应力[3]。当芽胞处于损伤状态时以一种超休眠态,即要经过很长时间才能进入萌芽状态。但将之置于适宜的环境下时,芽胞的激活、萌发、发芽和生长过程中的一系列生化反应会终止芽胞的休眠状态。目前在此方面的研究报道大部分较为深入地探讨了正常状态下芽胞萌芽的营养的因素[7-10]。Paidhungat等[10]研究了营养元素对枯草芽孢杆菌芽胞萌芽的影响,研究表明DPA-Ca (2,6-吡啶二羧酸钙盐)和营养成分导致枯草芽孢杆菌芽胞萌芽显著增加。Pandey等[11]通过对枯草芽孢杆菌单个孢子萌芽的活细胞成像图的分析,总结出枯草芽孢杆菌经热处理后,随着萌芽时间的延长,平均萌芽率也随之增加。Atluri等[12]也发现葡萄糖能诱导枯草芽孢杆菌的萌芽。本实验主要研究的是在食品中存在的常见营养素对受损芽胞的萌芽的影响,结果显示,碳源对枯草芽孢杆菌芽胞的萌芽存在一定的诱导作用,其中以麦芽糖萌芽率最高,该结果与上述文献报道的结果基本一致。
2.1.2不同氮源因子对芽胞萌芽率的影响不同氮源对芽胞萌芽率存在明显诱导作用,在Tris-HCl缓冲溶液里分别添加了牛血清蛋白、乳清蛋白、卵清蛋白、蛋白胨和牛肉膏5种氮源。利用SPSS多重方差分析结果表明卵清蛋白、乳清蛋白分别与同实验的其他四个因素均有显著性差异,而牛血清蛋白、牛肉膏、蛋白胨之间无显著性差异,与卵清蛋白、乳清蛋白两者之间有显著性差异。从图2结果可知,这五种氮源对芽胞萌芽的影响以乳清蛋白萌芽率最高,其次为牛血清蛋白、牛肉膏和蛋白胨,卵清蛋白的效果最差。
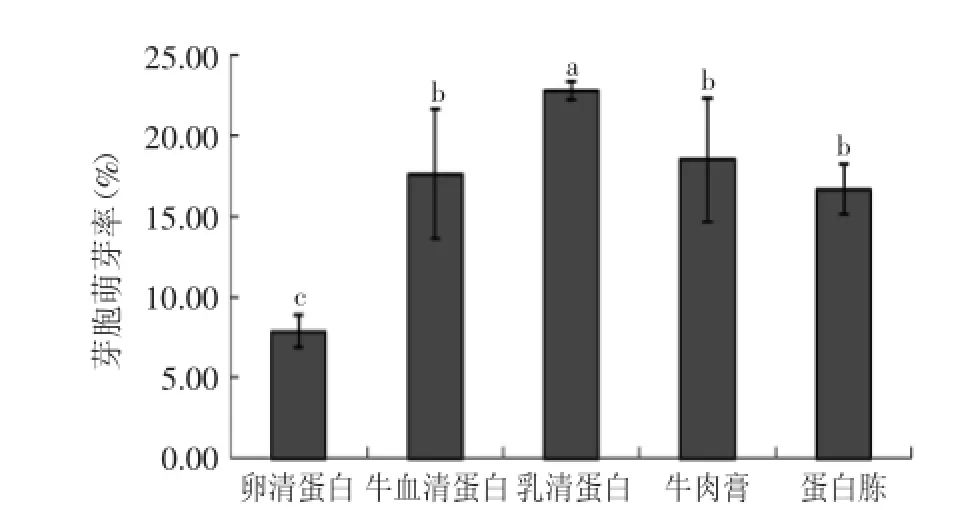
图2 不同氮源对芽胞萌芽率的影响Fig.2 Effects of different nitrogen sources on spore germination rates
2.1.3不同无机盐离子对芽胞萌芽率的影响无机盐不仅仅是细菌细胞繁殖过程中不可或缺的营养因子,也是影响芽胞萌芽率的主要因素。为了研究无机盐对芽胞萌芽率的影响,共选择了5种无机盐对萌芽率进行单一因素研究。图3结果表明,MgSO4对芽胞萌芽率的影响最为显著,MnSO4、NaCl、CaCl2诱导芽胞的萌芽率则较低。SPSS分析结果显示MgSO4和KCl分别与同实验的其他三个因素均有显著性差异,MnSO4、NaCl、CaCl2三者之间无显著性差异,MgSO4与KCl有显著性差异。

图3 不同无机盐对芽胞萌芽率的影响Fig.3 Effects of different inorganic salts on spore germination rates
本研究结果表明,乳清蛋白是芽胞萌芽的最佳氮源;无机盐MgSO4对芽胞萌芽产生显著的诱导作用(14.43%)。究其原因,这些营养成分诱导萌芽可视为一种萌芽的自然途径,萌芽可能始于存在于芽胞衣膜上特定的受体,这些受体可能被某些营养物质结合而被激活[10]。随后,释放出DPA,芽胞的通透性也随之增加,核心部分再次激活CwlJ和SleB水解酶。该水解酶能水解皮质层,此点对扩大其核心的空间显得尤为重要。待皮质消失后,充分水化的核心部分会恢复各种合成酶的活性和合成ATP合成的前体物质3-PGA(3-磷酸甘油酸)。随着SASP(酸溶性芽胞蛋白,该蛋白可防止DNA的损伤)的降解,逐步释放出的DNA,启动RNA,合成菌体生长所需要DNA、RNA物质和蛋白质等物质[13]。
2.2温度、pH、接种量、溶氧量对芽胞萌芽的影响
由正交实验的结果可知,不同条件下枯草芽孢杆菌芽胞萌芽率表现不同,以A3B2C1D3实验组的萌芽率最高,为14.29%。从极差R结果可知,对枯草芽孢杆菌芽胞萌芽率变化最大的是pH,其次是溶氧量、温度,而对芽胞萌芽率的变化最小的为接种量。经正交实验优化得出,导致枯草芽孢杆菌芽胞萌芽的最佳组合是A3B2C3D3,因此对最佳组合进行验证实验。结果表明理论优化的萌芽率为15.27%,枯草芽孢杆菌芽胞萌芽率的最佳方案为A3B2C3D3,即为37℃、pH7.0、最佳接种量为4%、最佳溶氧量是70 mL时得到的萌芽率最高。
从理论上推导孢子萌芽的最适温度范围应该在30~40℃内。Chedia等[5]研究结果表明,在温度高于40℃或低于30℃时孢子萌芽率显著下降,该结果与本实验的结果报道一致。Ciarciaglini(2000)报道枯草芽孢杆菌pSB357的萌芽率在L-丙氨酸诱导下pH为4.0时萌芽率将会完全受到抑制[14]。其机制可能在于,较低pH改变了氨基酸侧链,从而改变其电荷分布、氢键的状态和蛋白质的构象[15-16]。从溶氧量和接种量的结果可观察到,在本实验设计的浓度范围内,溶氧量和接种量越大,萌芽率越高。溶氧量可能与菌体本身的生理生化性质有关。芽胞萌芽的速度通常很快,仅需要数分钟,当芽胞发芽后便恢复成营养体细胞的形态。枯草芽孢杆菌为一类好氧型细菌,其在有氧的情况下,呼吸作用增强,新陈代谢作用加快,更有利于它营养细胞的生长。
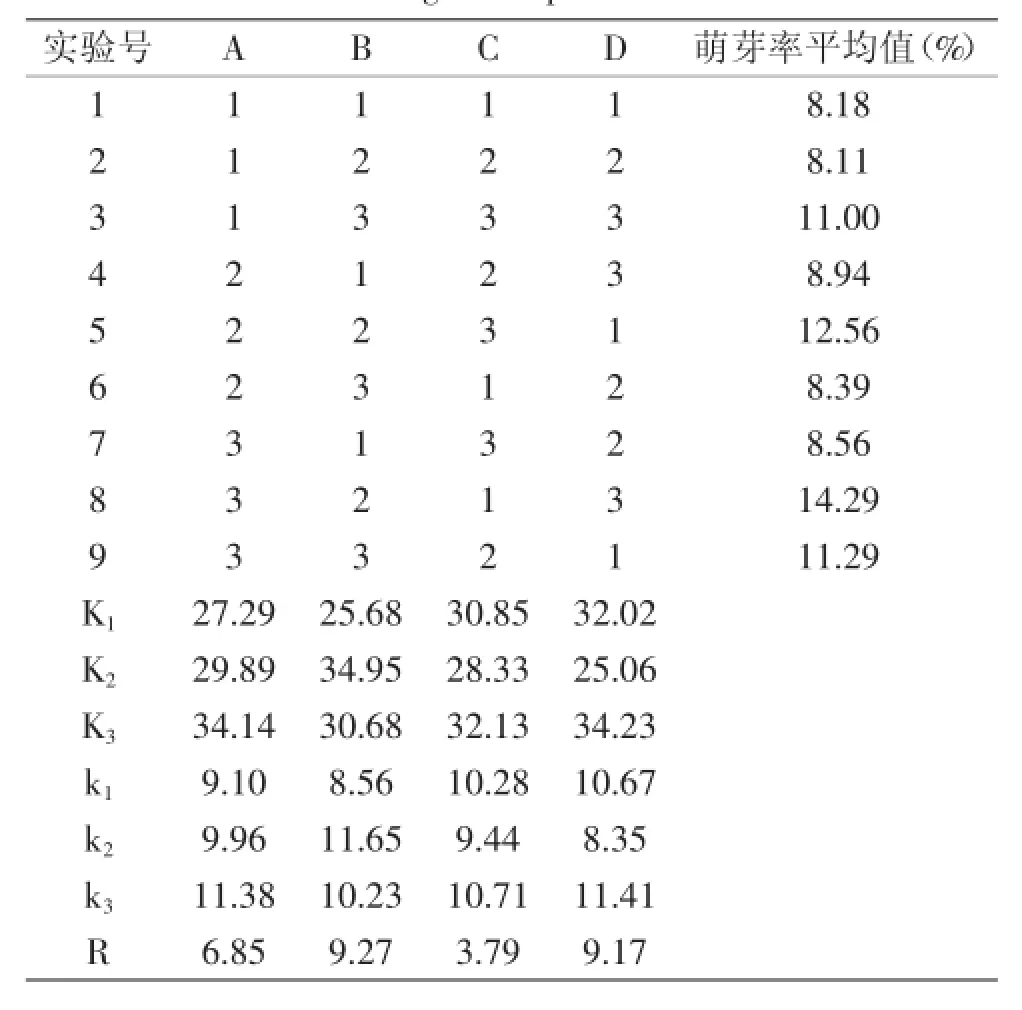
表2 枯草芽孢杆菌芽胞萌芽率正交实验结果Table 2 Results of Bacillus subtilis spore germination rate by orthogonal experiment
3 结论
营养成分、适宜的温度、pH、接种量和溶氧量能刺激芽胞的萌芽,不适宜营养成分浓度、温度、pH、接种量和溶氧量则会改变菌体的形态、代谢能力,甚至导致死亡。综合本实验的结果可知,食品中存在麦芽糖、乳清蛋白是有利于处于亚损伤状态下芽胞萌芽的条件;无机盐MgSO4对芽胞萌芽能产生显著的诱导作用;最佳萌芽温度为37℃、pH7.0、接种量4%、溶氧量则越大越有利于萌芽。该结果对于指导食品工业生产中合理控制芽胞菌的萌芽、生长与繁殖提供了数据支撑。
[1]姜成林,徐丽华.微生物资源开发利用[M].北京:中国轻工业出版社,2001.
[2]何国庆,贾英明.食品微生物学[M].北京:中国农业大学出版社,2002.
[3]雷伊,布恩亚,江汉湖.基础食品微生物学[M].北京:中国轻工出版社,2014.
[4]钱昆,周涛.低温熏煮香肠中腐败菌的分离及鉴定[J].食品工业科技,2008(4):124-126.
[5]Chedia Aouadhi,Hélène Simonin,Abderrazak Maaroufi,et al. Optimizationofnutrient-inducedgerminationofBacillus sporothermodurans spores using response surface methodology[J]. J Food Microbiol,2013,36:320-326.
[6]郭夏丽,狄源宁,王岩.枯草芽孢杆菌产芽胞条件的优化[J].中国土壤与肥料,2012(3):99-107.
[7]白艳红,赵电波,杨公明,等.蛋清溶菌酶对枯草芽孢杆菌芽胞萌芽的影响[J].食品工业科技,2008,29(11):213-215.
[8]姚露燕,张水华,曹昱,等.金属离子浓度对枯草芽孢杆菌芽胞率的影响[J].现在食品科技,2008,8(24):770-772.
[9]张宝善,王君,陈锦屏,等.大蒜汁对枯草芽孢杆菌抑制作用的研究[J].西北植物学报,2009,29(6):1269-1275.
[10]Paidhungat M,Setlow P.Role of Ger proteins in nutrient and nonnutrient triggering of spore germination in Bacillus subtilis [J].Journal of Bacteriology,2000,182(9):2513-2519.
[11]Pandey R,Ter Beek A,Vischer N O,et al.Live cell imaging of germination and outgrowth of individual Bacillus subtilis spores;the effect of heat stress quantitatively analyzed with Spore Tracker[J].PloS one,2013,8(3):e58972.
[12]Atluri S,Ragkousi K,Cortezzo D E,et al.Cooperativity between different nutrient receptors in germination of spores of Bacillus subtilis and reduction of this cooperativity by alterations in the GerB receptor[J].Journal of Bacteriology,2006,188(1):28-36.
[13]Setlow B,Cowan A E,Setlow P.Germination of spores of Bacillussubtiliswithdodecylamine[J].JournalofApplied Microbiology,2003,95(3):637-648.
[14]Ciarciaglini G,Hill P J,Davies K,et al.Germinationinduced bioluminescence,a route to determine the inhibitory effect of a combination preservation treatment on bacterial spores [J].Applied and Environmental Microbiology,2000,66(9):3735-3742.
[15]Broussolle V,Gauillard F,Nguyen C,et al.Diversity of spore germination in response to inosine and L-alanine and its interaction with NaCl and pH in the Bacillus cereus group[J]. Journal of Applied Microbiology,2008,105(4):1081-1090.
[16]Kort R,Brul S.Modelling the effect of sub(lethal)heat treatment of Bacillus subtilis spores on germination rate and outgrowth to exponentially growing vegetative cells[J].Int J Food Microbiol,2008,128:34-40.
Effects of germination factors on Bacillus subtilis spores in the sub(lethal)stage
CHANG Chao,PAN Li,WU Jin-e,ZHANG Zhe (College of Food Science and Engineering,Wuhan Polytechnic University,Wuhan 430023,China)
The effect of food nutritional ingredient on Bacillus subtilis spores in the sub(lethal)stage germination was evaluated,which the impact of carbon,nitrogen,inorganic salts in food on spore germinate was conducted by batch culture method,and temperature,pH,inoculum size,oxygen content was studied by orthogonal experimental design.To quantitatively analyse the effect of these factors,the optical density of OD580was determined to calculate germination rate.The results showed that maltose,whey protein were the best C source and N source of spore germination,respectively.MgSO4had a significant induction effect on spore germination. Based on the best nutrients of maltose(5 mg/mL),MgSO4(5 mg/mL),whey protein(5 mg/mL),optimal growth conditions for spore was that an inoculum of 4%of Bacillus subtilis spores in media with pH7.0 were cultured under 37℃,and the dissolved oxygen was 70 mL,constant shaking for 60 min,resulted in the maximum spore germination rate was 15.27%.
Bacillus subtilis spore;sublethal state;germinate rate;influence factor
TS201.1
A
1002-0306(2016)04-0242-04
10.13386/j.issn1002-0306.2016.04.040
2015-06-12
常超(1976-),男,硕士,副教授,研究方向:食品质量与安全,E-mail:changchao2000@163.com。
湖北省教育厅科技计划项目(D20131701)。
