弱酸胁迫提升富硒产朊假丝酵母性能及其作用机理
2016-09-14王冬华王大慧卫功元苏州大学医学部基础医学与生物科学学院江苏苏州215123
王冬华,王大慧,卫功元(苏州大学医学部基础医学与生物科学学院,江苏苏州215123)
弱酸胁迫提升富硒产朊假丝酵母性能及其作用机理
王冬华,王大慧,卫功元
(苏州大学医学部基础医学与生物科学学院,江苏苏州215123)
以一株耐亚硒酸钠的产朊假丝酵母为研究对象,考察了该菌株在不同pH环境下的富硒和谷胱甘肽(GSH)合成能力及抗氧化性能。结果表明,在弱酸胁迫(pH3.5)条件下,富硒产朊假丝酵母胞内有机硒和GSH含量均达到最高水平,分别为1.08 mg/g和18.48 mg/g。通过对酵母胞内γ-谷氨酰半胱氨酸合成酶、过氧化氢酶、超氧化物歧化酶的活性以及丙二醛含量进行测定,发现弱酸胁迫不仅有利于增强GSH的合成能力,也有助于提高酵母细胞的抗氧化能力。该研究结果为发酵法制备高性能富硒产朊假丝酵母提供了可行的技术参考。
富硒酵母,酸胁迫,有机硒含量,谷胱甘肽,产朊假丝酵母
硒是人体必需的微量元素之一,缺硒将导致相关疾病的发生,而通过饮食合理补硒则可以起到预防作用[1]。硒的补充途径有多种,其中富硒酵母因富含B族维生素、食用安全以及生物利用率高等优点而成为补硒食品的首选[2-4]。以往,酿酒酵母一直作为富硒的主要微生物而得到广泛关注[5],然而,近年来利用产朊假丝酵母(Candida utilis)进行有机硒的富集为富硒酵母的制备和应用提供了新的方法和手段[6]。作为一种GRAS微生物,产朊假丝酵母比酿酒酵母具有更宽广的营养谱及更高的谷胱甘肽(GSH)合成能力,因而在富硒制品中将具有更大的应用潜力[7-8]。
在利用发酵法制备富硒产朊假丝酵母的过程中,生物量、胞内有机硒含量是体现其发酵水平的重要技术指标,一般可以通过菌株选育和发酵条件优化等方法来提高其性能[6,9-10]。然而,若能提高增加胞内GSH含量,则对于提升富硒产朊假丝酵母的性能将是大有裨益的[11]。为此,作者所在的团队采用了包括添加氨基酸[12-13]和酸胁迫[14]在内的各种手段实现了高性能富硒产朊假丝酵母的制备。在酸胁迫过程中,产朊假丝酵母将以过量合成GSH的形式来抵抗和适应外界的恶劣环境[15],而这一特点正好可以用来提高富硒酵母的性能。
酸胁迫常见于食品发酵工业中,不少微生物对外界的酸胁迫环境具有一定的抵抗和适应能力[16]。在前期工作中,作者在研究酸胁迫在富硒酵母制备中的作用时发现,高浓度无机硒对产朊假丝酵母的
生长具有抑制作用,所以采用分批补加无机硒[11]或在细胞生长结束时添加无机硒的方式[14]。但是,有研究结果表明,酵母高效富硒是伴随着细胞的快速生长而进行的[17],因此有必要对酸胁迫条件下酵母细胞在生长时的富硒过程进行研究。为此,本文在实验室选育获得的一株耐高浓度无机硒的产朊假丝酵母的基础上,以富硒酵母的高性能(高有机硒含量和高GSH含量)为目标,考察酸胁迫的作用及其机理,研究结果将为高性能富硒产朊假丝酵母的高效制备提供可行的技术参考。
1 材料与方法
1.1 材料与仪器
产朊假丝酵母(C.utilis Se-1301) 为一株耐高浓度亚硒酸钠菌株,是在实验室原有C.utilis SZU 07-01的基础上,通过在含有20 mg/L亚硒酸钠的平板上驯化选育获得的突变株;GSH、还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)和谷胱甘肽还原酶等生工生物工程(上海)股份有限公司;各类检测试剂盒 南京建成生物工程研究所;0.22 μm水系微孔滤膜 生工生物工程(上海)股份有限公司。
BIOTECH-5BG全自动发酵罐 上海保兴生物设备工程有限公司;T6新世纪紫外可见分光光度计北京普析通用仪器有限责任公司;LD5-2A离心机 北京京立离心机有限公司;SW-CJ-2A超净工作台 吴江市龙宏净化设备有限公司;HZ-2010K摇床 上海欣蕊自动化设备有限公司;LDZX-50KB灭菌锅 上海申安医疗器械厂。
1.2 实验方法
1.2.1 种子培养 斜面种子活化4 h后接入种子培养基(葡萄糖20 g/L,蛋白胨10 g/L,酵母粉10 g/L,pH6.0),摇床中培养20 h,温度30℃,摇床转速200 r/min。
1.2.2 不同pH条件下产朊假丝酵母分批培养 发酵罐中装有3 L发酵培养基(葡萄糖30 g/L,硫酸铵8 g/L,磷酸二氢钾3 g/L,硫酸镁0.25 g/L,L-蛋氨酸10 mmol/L,pH5.5),按照10%(v/v)的接种量向其中接入种子,在400 r/min和27℃条件下培养30 h。培养液pH将分别恒定控制在2.5、3.5、4.5、5.5和6.5,pH采用梅特勒电极在位监测,通过自动流加3 mol/L H2SO4或3 mol/L NaOH进行调节,以维持pH变化在±0.02范围内。在不同培养时间定时取样,样品经处理后检测细胞生长和GSH合成情况。
1.2.3 不同pH条件下富硒产朊假丝酵母分批培养
种子接种和发酵条件同分批培养操作。在接种前将1 g/L亚硒酸钠溶液(经滤膜过滤除菌)加入发酵培养基中,至总浓度15 mg/L。培养液pH将分别恒定控制在2.5、3.5、4.5、5.5和6.5,在不同培养时间定时取样,样品经处理后检测细胞生长和GSH合成情况,在发酵结束时(30 h)测定胞内总有机硒含量。
1.2.4 不同pH条件下富硒产朊假丝酵母性能比较在富硒产朊假丝酵母分批培养过程中,定时取样,检测9、15、21 h时的γ-谷氨酰半胱氨酸合成酶活性,测定15、21 h时的过氧化氢酶和超氧化物歧化酶活性以及胞内丙二醛含量,比较不同pH条件下各酶活大小,总结弱酸胁迫提升富硒产朊假丝酵母性能的内在机理。
1.3 测定方法
1.3.1 酵母生物量的测定 以细胞干重(DCW)表示酵母生物量,取20 mL发酵液,在3500 r/min下离心10 min,蒸馏水洗涤3次,湿菌体在70℃下烘干至恒重。
1.3.2 胞内GSH的提取、测定及胞内GSH含量(IGC)的计算 参见文献[9]。
1.3.3 无机硒和胞内总硒的测定、胞内有机硒含量(ISeC)的计算 参见文献[11]。
1.3.4 γ-谷氨酰半胱氨酸合成酶(γ-GCS)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)的活性和胞内丙二醛(MDA)含量的测定 采用试剂盒进行测定,操作方法和步骤按照说明书进行。根据实验的具体要求,分别在产朊假丝酵母快速生长期(9 h)、GSH快速合成期(15 h)以及酵母细胞生长稳定期(21 h)对γ-GCS、CAT 和SOD酶活以及胞内MDA含量的变化进行测定。
1.4 数据处理与分析
所有摇瓶实验数据均为三组独立实验样品的平均值,所有分批发酵实验数据均为两次检测结果的平均值。实验数据的可信度采用t-检验进行分析,当p≤0.05时认为有显著影响。统计学分析软件为Statistical Analysis System(SAS 9.0)。
2 结果与讨论
2.1 不同pH条件下产朊假丝酵母分批培养结果

图1 不同pH条件下产朊假丝酵母细胞生长与GSH合成量变化Fig.1 Cell growth of C.utilis Se-1301 and GSH biosynthesis under different pHs
考察了突变株C.utilis Se-1301在不同pH(2.5、3.5、4.5、5.5、6.5)条件下的分批培养情况,并以此作为以下实验的对照。图1显示了突变株在不富集硒时的细胞生长和GSH合成情况,结果表明,该酵母细胞
能在较宽的pH范围内正常生长并合成GSH。除了pH2.5以外,其他pH条件下酵母细胞的生长情况基本一致,最大细胞干重没有明显差异。然而,不同pH条件下的GSH合成结果显示,GSH产量在pH3.5~5.5范围内均维持在较高的水平。值得一提的是,只有当pH5.5时细胞才能快速地合成最大量的GSH,18 h时胞内GSH含量达到最大值18.27 mg/g。与文献[10]中的C.utilis SZU 07-01分批培养结果相比,图1所示的突变株具有更高的GSH合成能力。
2.2 不同pH条件下酵母细胞生长和富硒情况
2.2.1 分批发酵过程参数 在分批培养的0 h向发酵培养基中添加15 mg/L亚硒酸钠,考察C.utilis Se-1301在不同pH(2.5、3.5、4.5、5.5、6.5)条件下细胞生长和GSH合成的变化情况,结果见图2。可以看出,酵母细胞在亚硒酸钠存在的情况下也可以在较宽的pH范围(3.5~6.5)内正常生长,只有在pH2.5时细胞生长速度受到了抑制,最终细胞干重也明显低于其他pH环境下的结果。相应地,GSH合成受pH的影响与细胞生长规律较为类似。
产朊假丝酵母在富硒环境条件下,不同pH条件下的细胞干重(图2)与对照条件(图1)相比虽略有下降,但差异不大,显示出突变株具有良好的适应高浓度无机硒并正常生长的能力。针对GSH合成,虽然最终GSH最大合成量与对照(图1)相比较为接近,但GSH合成速度在富硒过程中则稍微滞后于对照,说明酵母胞内与GSH合成有关的代谢速率受到了无机硒富集的影响。
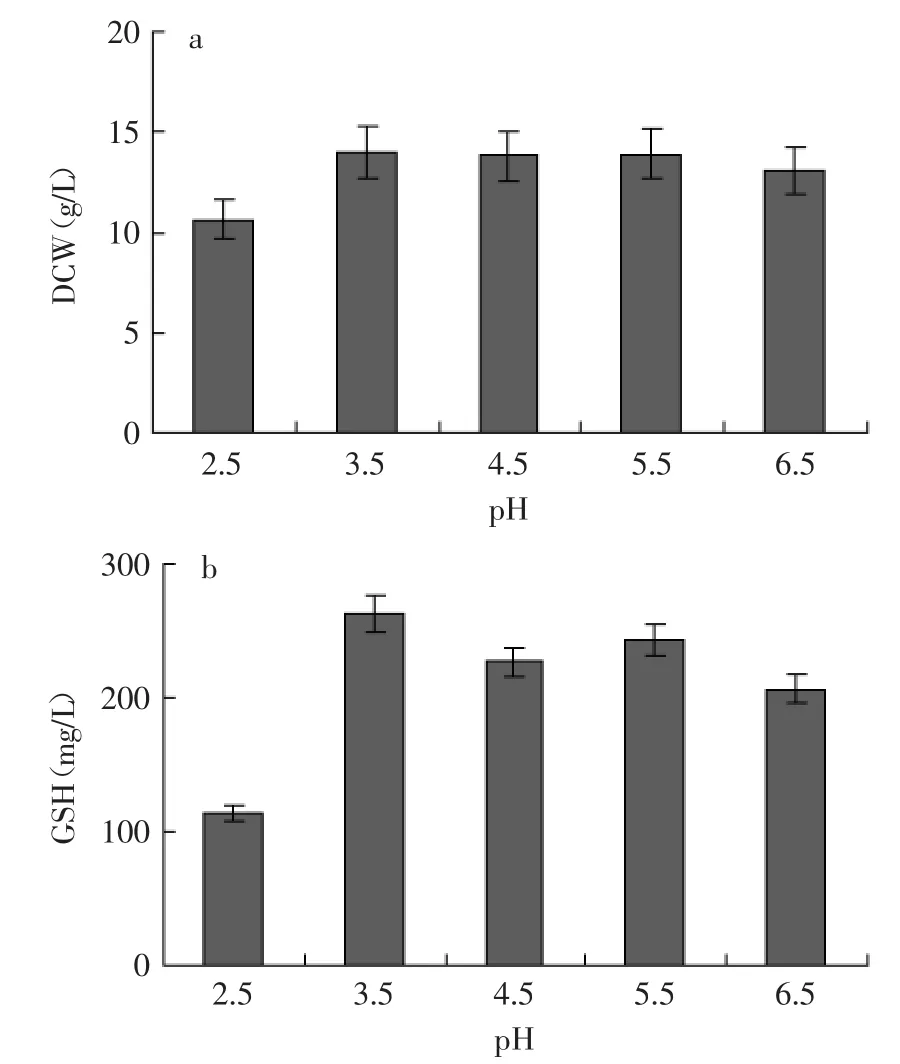
图2 不同pH条件下的富硒产朊假丝酵母细胞生长和GSH合成量变化Fig.2 Cell growth and GSH biosynthesis during selenium enrichment under different pHs
值得关注的是,pH3.5时的酵母细胞干重和GSH合成量均高于其他pH条件下的结果。由此可以发现,弱酸胁迫环境(如pH3.5)有利于产朊假丝酵母在富硒过程中过量合成GSH,从而有效提高富硒酵母的性能。该实验结果与前期研究中的C.utilis SZU 07-01在细胞生长结束时富硒所得出的结论[14]是一致的。
2.2.2 富硒产朊假丝酵母性能 比较了发酵结束时不同pH(2.5、3.5、4.5、5.5、6.5)条件下富硒产朊假丝酵母胞内有机硒(ISeC)和GSH含量(IGC),结果见图3。除了pH2.5以外,在其他pH条件下的IGC都处于较高的水平,其中pH3.5和pH5.5时IGC最大值在21 h分别为18.48 mg/g和18.25 mg/g,与对照的胞内GSH含量基本相当,说明产朊假丝酵母的富硒过程没有对GSH合成能力产生不良影响。然而,不同pH条件对酵母的富硒能力有不同的影响,其中弱酸胁迫(pH3.5)下的酵母细胞具有最强的富硒能力,胞内有机硒含量达到为1.08 mg/g。
综合图3中的富硒酵母的两项关键性能指标IGC 和ISeC可以发现,与其他pH条件相比,pH3.5最有利于富硒产朊假丝酵母性能的提高。以下将从酵母胞内相关酶活和物质水平变化规律等角度对弱酸胁迫提升富硒酵母性能的作用机理进行探讨。

图3 pH对富硒产朊假丝酵母胞内GSH和有机硒含量的影响Fig.3 Effect of pH on intracellular contents of both GSH and organic selenium
2.3 弱酸胁迫提升富硒产朊假丝酵母性能的机理

图4 酸胁迫条件下富硒产朊假丝酵母胞内γ-GCS酶活Fig.4 γ-GCS activities of selenium-enriched C.utilis under acid stress
2.3.1 γ-谷氨酰半胱氨酸合成酶活性 γ-谷氨酰半胱氨酸合成酶(γ-GCS)是酵母胞内GSH合成的关键限速酶,其活性的大小与GSH合成速率密切关联[18]。为此,测定了不同酸胁迫条件和不同培养时间下的富硒产朊假丝酵母胞内γ-GCS酶活,结果见图4。由
图4可见,pH2.5时的γ-GCS酶活显著(p<0.05)低于其他pH条件下的酶活水平,最终导致细胞合成的GSH量也较低(图2)。在pH3.5~6.5范围内,细胞快速生长期(9 h)和稳定期(21 h)时的γ-GCS酶活没有明显差异,但在GSH快速合成期(15 h),γ-GCS酶活只有在pH3.5时最高,表明在该pH条件下酵母细胞具有更强的GSH合成能力,因而更有利于促进富硒产朊假丝酵母性能的提升。
2.3.2 过氧化氢酶和超氧化物歧化酶活性 GSH是还原性物质,对于维持酵母胞内适宜的氧化还原环境起着重要的作用[18]。然而,酸胁迫环境以及富硒过程都容易引起胞内氧化性物质水平的提高,进而降低GSH的含量[12]。如果胞内参与自由基消除反应的过氧化氢酶(CAT)和超氧化物歧化酶(SOD)的活性较高,则更加有利于GSH水平的维持和提高。为此,分别测定了不同pH条件和不同培养时间下的CAT和SOD酶活,结果见图5。可以看出,在GSH快速合成期(15 h),CAT酶活在pH3.5和4.5下均处于较高水平;在稳定期(21 h)时,不同pH条件下CAT酶活没有明显差异(pH2.5除外)。结合图2的结果可以看出,CAT酶活的高低与GSH合成速率和产量都有一定的关联,当CAT活力下降时,GSH可能参与氧化还原反应而被消耗。对于SOD来说,其活性大小在不同pH和不同培养时间条件下没有显著差异,体现出富硒产朊假丝酵母胞内的SOD可能对弱酸胁迫不敏感,或者SOD对胞内GSH含量的变化没有实质性影响。

图5 酸胁迫条件下富硒产朊假丝酵母胞内CAT和SOD酶活Fig.5 The activities of CAT and SOD in selenium-enriched C.utilis under acid stress
2.3.3 丙二醛含量 丙二醛(MDA)是生物体内脂质与自由基发生过氧化反应的产物,其含量多少通常体现细胞内氧化性环境的强弱,也可以反映出细胞膜损伤程度和通透性的大小[19]。由于细胞膜通透性的增加将导致GSH的泄漏,不利于胞内GSH含量的维持和提高[20]。为此,测定了不同pH和不同培养时间下的富硒产朊假丝酵母胞内MDA含量,结果见图6。可以发现,pH3.5时的MDA含量较低,说明该条件下酵母胞内氧化性物质的水平要低于其他pH环境,同时细胞膜在富硒过程中受到的损伤也较小。此外,不同pH下的MDA含量在15 h和21 h时没有明显差异,说明不同培养时间下细胞膜的被氧化程度变化不大。
在酵母细胞转化亚硒酸钠为有机硒的过程中,亚硒酸钠的存在容易导致MDA的形成,增加了细胞膜的通透性,而从胞内渗漏出来的GSH又极易与培养基中的亚硒酸钠反应形成红色的单质硒[21],降低了有机硒转化率和胞内GSH含量,这对提高富硒酵母的性能是十分不利的。因此,通过弱酸胁迫提高胞内GSH含量并改善胞内氧化性环境是解决以上问题的有效方法之一。
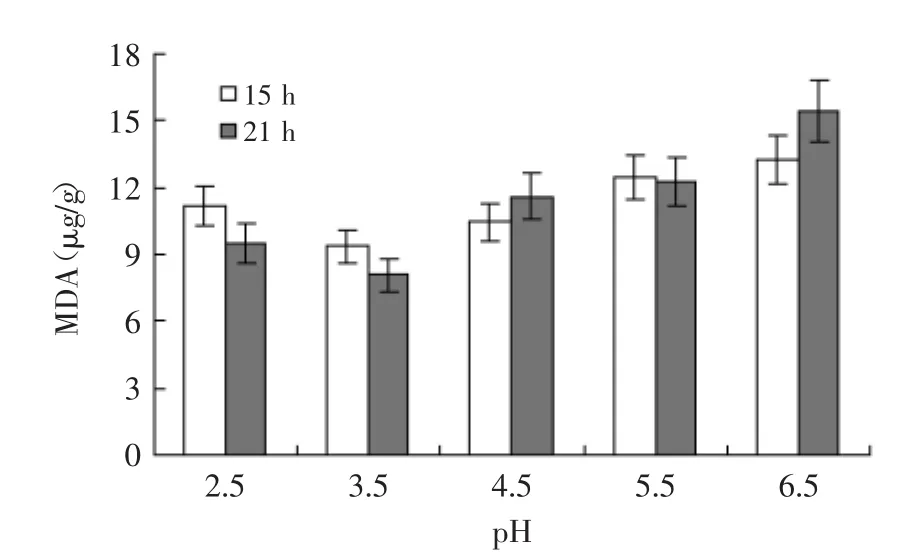
图6 酸胁迫条件下富硒产朊假丝酵母胞内丙二醛含量Fig.6 The intracellular MDA content of selenium-enriched C.utilis under acid stress
3 结论
通过对不同酸胁迫条件下的富硒产朊假丝酵母细胞生长以及GSH合成过程进行分析,发现弱酸胁迫(pH3.5)条件有利于富硒酵母胞内有机硒和GSH含量等性能的提高。富硒酵母分批培养过程中胞内相关酶活和物质水平表明,在弱酸胁迫条件下酵母细胞具有更强的GSH合成能力,更高的过氧化氢酶活性,以及更低的胞内丙二醛含量。弱酸胁迫条件下制备得到的富硒产朊假丝酵母不仅具有较高的性能,而且还具有较强的抗氧化能力,因此将具有更为广阔的应用前景。
[1]Kieliszek M,Blazejak S.Selenium:significance,and outlook for supplementation[J].Nutrition,2013,29(5):713-718.
[2]Schrauzer G N.Selenium yeast:composition,quality,analysis,and safety[J].Pure Appl Chem,2006,78(1):105-109.
[3]付俊鹤,余明华,许武桥,等.富硒酵母在面包中的应用[J].食品工业科技,2011,32(10):373-374,377.
[4]肖方正,刘曲滨.富硒酵母的研究开发与应用[J].广东微量元素科学,2001,8(8):7-10.
[5]范秀英,郭雪娜,傅秀辉,等.高生物量富硒酵母的选育及培养条件初步优化[J].生物工程学报,2003,19(6):720-724.
[6]牛海涛,汤燕花,谢必峰.制备富硒产朊假丝酵母(Candida utilis)的条件优化[J].药物生物技术,2007,14(2):123-126.
[7]Wang D,Yang B,Wei G,et al.Efficient preparation of selenium/glutathione-enriched Candida utilis and its biological effects on rats[J].Biol Trace Elem Res,2012,150(1-3):249-257.
[8]Yang B,Wang D,Wei G,et al.Selenium-enriched Candida utilis:Efficient preparation with L-methionine and antioxidant capacity in rats[J].J Trace Elem Med Bio,2013,27(1):7-11.
[9]葛晓光,卫功元,聂敏,等.富硒产朊假丝酵母的制备条件研究[J].粮食与饲料工业,2009(10):31-33.
[10]王大慧,杨波,卫功元,等.提高产朊假丝酵母富硒能力的工艺条件研究[J].食品与机械,2011,27(6):245-248.
[11]杨波,刘志奎,卫功元,等.培养方式对富硒产朊假丝酵母性能的影响[J].生物加工过程,2012,10(4):7-11.
[12]Ge X,Wang D,Wei G,et al.Improvement of physiological characteristic of selenium-enriched Candida utilis with amino acids addition[J].Biotechnol Res Int,2010,2011:1-7.
[13]王大慧,葛晓光,卫功元,等.高性能富硒产朊假丝酵母的制备[J].食品与发酵工业,2011,37(6):106-109.
[14]王大慧,许宏庆,汪成富,等.酸胁迫在提升富硒/GSH产朊假丝酵母性能中的作用[J].中国生物工程杂志,2013,33(11):81-85.
[15]董颖颖,卫功元,张君丽,等.谷胱甘肽生物合成过程中酸胁迫的作用及其机制[J].化工学报,2011,62(11):3228-3235.
[16]Ter Beek A,Wijman J G,Zakrzewska A,et al.Comparative physiological and transcriptional analysis of weak organic acid stress in Bacillus subtilis[J].Food Microbiol,2015,45:71-82.
[17]王世平,周玉岩,滕冰.酵母中有机硒转化的研究[J].东北农业大学学报,1997,28(4):378-383.
[18]Li Y,Wei G,Chen J.Glutathione:a review on biotechnological production[J].Appl Microb Biotech,2004,66(3):233-242.
[19]Izawa S,Inoue Y,Kimura A.Oxidative stress response in yeast:effect of glutathione on adaptation to hydrogen peroxide stress in Saccharomyces cerevisiae[J].FEBS Lett,1995,368(1):73-76.
[20]Nie W,Wei G,Du G,et al.Enhanced intracellular glutathione synthesis and excretion capability of Candida utilis by using a low pH-stress strategy[J].Lett Appl Microbiol,2005,40(5):378-384.
[21]Iizuka M,Murata K,Kimura A.Induction of glutathione leakage from Saccharomyces cerevisiae cells by selenite[J].Agric Biol Chem,1988,52(2):613-614.
Moderate acid stress improves the performance of selenium-enriched Candida utilis and its related mechanism
WANG Dong-hua,WANG Da-hui,WEI Gong-yuan*
(School of Biology and Basic Medical Sciences,Soochow University,Suzhou 215123,China)
The capacities of selenium enrichment and glutathione biosynthesis as well as antioxidant properties of a sodium selenite-resisting Candida utilis Se-1301 were investigated in this study.The results illustrated that the highest intracellular organic selenium and glutathione contents of 1.08 mg/g and 18.48 mg/g,respectively,were obtained under a moderate acid stress environment(pH3.5).After assaying the activities of intracellular γ-glutamylcysteine synthetase,catalase and superoxide dismutase,together with determining the intracellular malondialdehyde content,the moderate acid stress was found to be not only favored to improve the ability of glutathione biosynthesis,but also helped to increase the antioxidant capacity of the yeast.The results provided a feasible route for efficient preparation of selenium-enriched Candida utilis with high performance.
selenium-enriched yeast;acid stress;organic selenium content;glutathione;Candida utilis
TS201.1
A
1002-0306(2016)08-0191-05
10.13386/j.issn1002-0306.2016.08.031
2015-09-10
王冬华(1988-),女,硕士研究生,研究方向:食品添加剂的微生物制造,E-mail:1013813289@qq.com。
*通讯作者:卫功元(1975-),男,博士,教授,研究方向:微生物发酵工程,E-mail:weigy@suda.edu.cn。
国家自然科学基金(21506136);苏州市应用基础研究计划项目(SYN201314)。
