转磷酸甘露糖变位酶基因提高水稻维生素C含量
2016-08-09高利芬夏志辉张继王道文翟文学
高利芬 夏志辉 张继 王道文 翟文学
(1江汉大学 系统生物学研究院, 武汉 430056; 2中国科学院 遗传与发育生物学研究所, 北京 100101;3海南大学 生命科学研究院, 海口 570228;*通讯联系人, E-mail: wxzhai@genetics.ac.cn)
转磷酸甘露糖变位酶基因提高水稻维生素C含量
高利芬1,2夏志辉2,3张继1王道文2翟文学2,*
(1江汉大学 系统生物学研究院, 武汉 430056;2中国科学院 遗传与发育生物学研究所, 北京 100101;3海南大学 生命科学研究院, 海口 570228;*通讯联系人,E-mail:wxzhai@genetics.ac.cn)
GAO Lifen, XIA Zhihui, ZHANG Ji, et al. Transgenosis of the phosphomannomutase transgene increases vitamin C content in rice. Chin J Rice Sci, 2016, 30(4): 441-446.
高利芬, 夏志辉, 张继, 等. 转磷酸甘露糖变位酶基因提高水稻维生素C含量. 中国水稻科学, 2016, 30(4): 441-446.
摘要:维生素C (VC) 是人体健康所必需的营养元素。人类由于缺乏VC合成途径中的最后一种酶(L-古洛糖酸内酯氧化酶),自身不能合成VC。水稻是重要的粮食作物,增加水稻种子中VC含量,能够提高其营养价值。磷酸甘露糖变位酶(PMM)是VC合成通路中一种重要的酶,催化甘露糖-6-磷酸到甘露糖-1-磷酸的转变。将水稻PMM基因(OsPMM)构建在双右边界双元载体pMNDRBBin6上,并用种子特异表达的启动子BX14驱动其表达。通过农杆菌介导的转化系统,OsPMM基因被转入粳型三系恢复系C418中。通过分子检测,在T2代筛选到了无选择标记的转基因植株。对OsPMM基因在转基因植株中的表达进行分析,发现OsPMM基因在转基因水稻种子内的表达水平明显提高,相应地,转基因系种子中的VC含量也提高了25%~50%。
关键词:维生素C; 磷酸甘露糖变位酶; 双右边界双元载体系统; 无选择标记; 转基因系
维生素C(VC)又名抗坏血酸(L-ascorbic acid, AsA),是动植物体内一种重要的抗氧化剂,参与了人体的心血管功能、免疫细胞发育、结缔组织和铁离子的利用等生理进程。植物和大多数动物可以合成VC, 人类由于缺乏VC合成途径中的最后一种酶L-古洛糖酸内酯氧化酶(L-gulonolactone oxidase),自身不能合成VC,加上VC不能在体内长期贮存,人类必须每天从膳食中获取足够VC。
水稻是重要的粮食作物,增加水稻种子中VC含量,能够提高其营养价值。到目前为止,已经在植物体内发现了多条VC生物合成途径和一条循环利用途径,其中VC生物合成途径包括L-半乳糖(L-galactose)途径(Wheeler-Smirnoff 途径)[1-3]、L-古洛糖(L-gulose)途径[4]、肌醇(myo-inositol)途径[5]和D-半乳糖醛酸(L-galactonic acid)途径[6]等。
通过对VC合成和代谢通路上的酶进行基因操作,可以提高或降低植物体内的VC水平。在土豆中的研究表明,反义抑制GDP-甘露糖焦磷酸化酶导致了VC含量的大幅下降[7]。在拟南芥中反义抑制L-半乳糖脱氢酶,在强光下生长的转基因植株的VC含量比野生型要降低很多[8]。在烟草中,抑制L-半乳糖醛酸-1,4-内酯脱氢酶的活性,导致了转基因株系VC含量较对照减少30%[9]。而Tokunaga等[10]在烟草悬浮细胞内过量表达L-半乳糖醛酸-1,4-内酯脱氢酶,使得总VC含量提高了2倍左右。不仅如此,一些研究还发现,在植物内表达异源的VC合成或代谢通路上的酶,也能对VC的含量进行调控。在拟南芥中过量表达草莓的D-半乳糖醛酸还原酶导致VC的含量增加2~3倍[6]。L-古洛糖是动物体内VC合成的前体物质,在莴苣和烟草中过量表达老鼠的古洛糖内酯氧化酶使VC含量提高达7倍[11];在拟南芥中表达此酶也使VC含量提高了3倍多[12]。脱氢抗坏血酸还原酶(Dehydroascorbate reductase,DHAR) 参与了VC在植物体内的循环[13],在烟草和玉米中过表达来自小麦的脱氢抗坏血酸还原酶,其表达水平在烟草和玉米中分别达到原来的32倍和100倍,相应地,叶片和谷粒中的VC水平也提高了2~4倍[13]。
磷酸甘露糖变位酶(PMM)催化VC合成通路中甘露糖-6-磷酸到甘露糖-1-磷酸的转化,此反应是可逆的。对植物PMM的转基因操作发现,在烟草叶片中抑制PMM的表达可导致烟草VC含量降低,而增加PMM在叶片中的表达能使VC含量提高20%~50%。在拟南芥中过表达拟南芥PMM也导致了VC含量提高25%~33%[14]。在烟草中过表达来自樱桃的PMM基因,烟草的VC含量提高了两倍,且VC含量的提高和PMM基因的转录水平和酶活性正相关[15]。
以上报道表明,增加植物体内VC合成通路或循环通路中酶的表达,可以提高受体的VC含量。水稻是重要的粮食作物,暂未有通过基因工程提高水稻VC含量的报道。水稻PMM基因(OsPMM,LOC_Os04g58580)位于水稻第4染色体,基因组全长3033 bp,cDNA全长747 bp,编码含249个氨基酸的磷酸甘露糖变位酶,催化VC合成通路中甘露糖-6-磷酸到甘露糖-1-磷酸的转化,所催化的反应是可逆的。
为了提高水稻种子的VC含量,本研究将OsPMM的全长cDNA和种子特异表达的启动子Bx14融合,构建成了表达载体pDRBOsPMM。转基因质粒的骨架选择的是双右边界双元载体pMNDRBBin6,利用此载体,在转基因系后代的分离过程中可以筛选到无选择标记的转基因植株,无标记转基因植株的分离步骤可参见Lu等[16]的报道。通过农杆菌介导的转化,将OsPMM整合到水稻杂交稻骨干亲本C418中,在T2代筛选获得了无选择标记且种子VC含量明显提高的C418转基因纯合系。
1材料与方法
1.1试验材料
转基因受体材料是生产中常用的粳型恢复系C418。之所以选择C418,是因为C418是籼型遗传成分较高、形态偏籼且有特异亲和力的粳型恢复系,是起着“籼粳架桥”作用的品种。 C418与粳稻杂交子一代结实正常,与籼稻杂交子一代结实率 40%~65%。经与多个不育系配组, 其所配组合具有高产、优质、抗病、抗倒、高光效、高结实率、高配合力等诸多特点,在生产中具有重要的应用价值[17]。
1.2质粒构建和水稻转化
转基因质粒pDRBOsPMM是将OsPMM的全长cDNA序列和来自小麦高分子量麦谷蛋白(HMW glutenin)Glu-1B-1Bx基因的种子特异表达启动子Bx14融合构建而成的双元表达载体,由中国科学院遗传与发育生物学研究所王道文研究员提供。通过农杆菌介导的转化,OsPMM基因被转入到C418品种中,水稻组织培养的方法参见Zhai等[18]的报道。
1.3转基因植株的分子检测
在转基因植株生长到高分蘖期时,采集叶片分别提取DNA和RNA进行转基因植株的分子检测。DNA提取采用CTAB法;RNA的提取采用Trizol法,脱氧核糖核酸酶Ⅰ处理后,用Promega公司的M-MLV反转录酶,以Olig(dT)15为引物进行cDNA第1链的合成,具体过程参照厂家说明书。通表1转基因植株分子检测所用引物过对T0、T1和T2代转基因植株的OsPMM和标记基因Hpt(潮霉素抗性基因,Hygromycin),进行连续的PCR检测来筛选无选择标记的转基因纯合系;目标基因在转基因纯合系种子中的表达通过实时定量PCR进行检测,实时PCR检测使用的是Transgene公司的定量PCR混合液(TransStart TipTop Green qPCR SuperMix),反应体系和反应程序按照相应的产品说明书进行,反应于ABI 7900实时PCR仪上扩增检测。以内源的Actin基因为参照基因,转基因受体材料C418为参照样本,对OsPMM在转基因植株种子部位的表达进行相对定量分析。2-ΔΔCT法[19]用于计算基因表达的相对变化,实验设置3次技术重复。转基因植株的分子检测所用的引物及引物序列如表 1所示。
Table 1. Primers used for molecular detection of transgenic plants.

引物名称Primername引物序列Primersequence(5'-3')OsPMM-F/RF:CACGCTTCATTACATTGCTGR:TATGTCCAATTGTCCGGTCHpt-F/RF:TAGGAGGGCGTGGATATGTCR:TACACAGCCATCGGTCCAGAActin-F/RF:AGCAACTGGGATGATATGGAR:CAGGGCGATGTAGGAAAGC
1.4VC含量的测定
在水稻收割晒干后,随机抽取10粒水稻种子,用研钵充分研磨后,进行水稻种子中总VC含量的测定,检测方法参照Gillespie等[20]的报道进行,3次重复。
2结果与分析
2.1无选择标记转OsPMM基因纯合系的获得
为了提高水稻种子的VC含量,将OsPMM基因的全长cDNA和种子特异表达的启动子Bx14融合,以双右边界双元载体pMNDRBBin6为骨架,构建成了转基因质粒pDRBOsPMM,其中标记基因Hpt位于载体的两个右边界RB1和RB2之间,而目标基因OsPMM位于RB2和LB之间,转基因质粒T-DNA区的线形图见图1。
通过农杆菌介导的转化,将OsPMM基因转入C418中,T0代共获得26个独立的转基因株系。在苗期提取T0代植株叶片的DNA,用OsPMM基因特异检测引物OsPMM-F/R和标记基因Hpt基因特异检测引物Hpt-F/R,对以上26个单株进行转基因阳性植株的PCR筛选。在转基因系中,OsPMM-F/R能扩增出814 bp和377 bp两个条带,其中814 bp的条带是以基因组中的OsPMM基因组序列为模板扩增所得,而377 bp则是以转入的cDNA序列为模板扩增而来。在非转基因系中,没有额外整合的OsPMMcDNA序列,故只能扩出一条814 bp的带。在含潮霉素标记基因的转基因单株中,Hpt-F/R能扩增出1035 bp的条带(图2)。通过对T0代检测,共有10个单株既能够扩增出OsPMM基因特异的条带,又能够扩增出Hpt基因特异的条带(图2)。此10个单株即为转基因阳性植株,用于后续研究。
在T0代植株生长成熟后,对以上10个转基因阳性植株单株套袋收种,并在下一个水稻种植季继续种植T1代种子,每个株系种植30~50个单株。待水稻生长至分蘖期,采集叶片,微量提取以上材料的叶片总DNA。以提取的DNA为模板,对T1代单株分别进行OsPMM基因和Hpt基因的PCR检测,OsPMM基因检测阳性而Hpt基因检测阴性的单株即为无选择标记转OsPMM基因植株。经过检测,T0代10个株系中,2号、3号和7号株系分别能分离出2个、3个和3个无标记转基因单株,依据来源将这些单株分别命名2-1,2-2,3-1,3-2,3-3,7-1,7-2和7-3。
对以上8个T1代单株套袋收种,继续种植T2代种子,每个系种植30个单株,并在水稻生长到分蘖期后,提取叶片DNA再次对每个株系的单株进行OsPMM基因和Hpt基因的PCR鉴定,从中筛选无选择标记的转基因纯合系。经过鉴定,上述8个株系中,有3个株系的T2代单株均显示为OsPMM基因阳性且Hpt基因阴性,为无选择标记的OsPMM转基因纯合系。以上3个株系分别来自于T1编号为2-1、3-1和7-1的单株。图 3为2-1 部分T2代单株的检测结果。
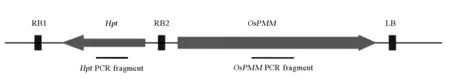
图1转基因质粒pDRBOsPMM T-DNA区的线形结构
Fig.1. Linear structure of transgenic plasmid pDRBOsPMM T-DNA zone.
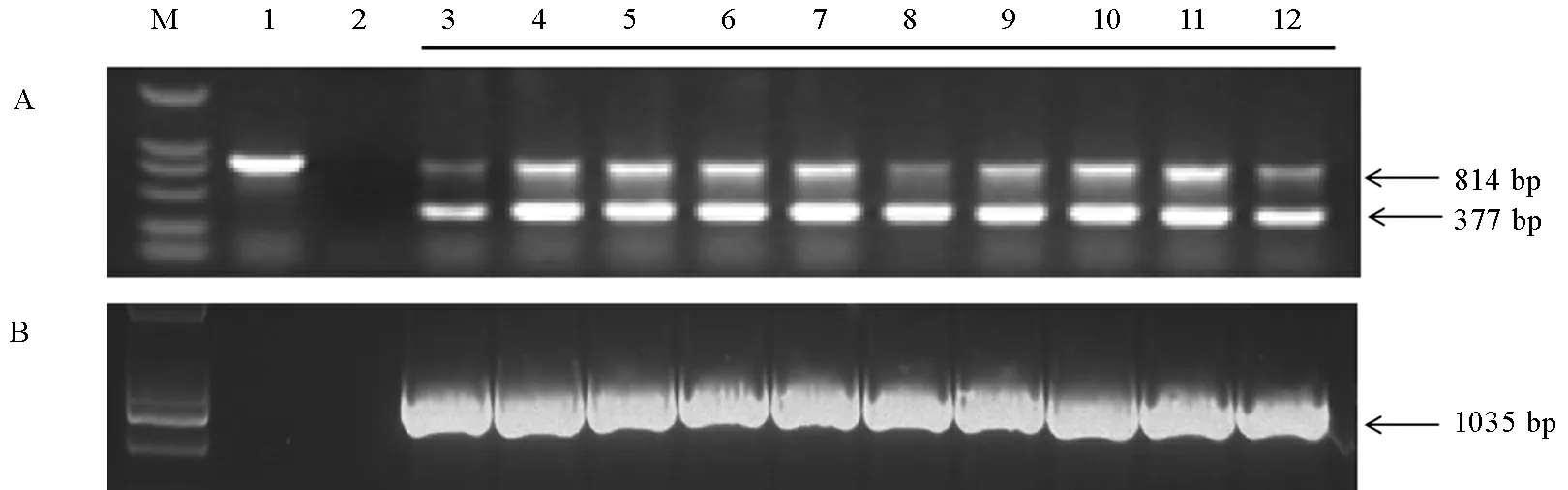
M-DL2000标记; 1-非转基因对照; 2-空白水对照; 3~12-转基因阳性植株。
M, DL2000 marker; 1, Non-transgenic control; 2, PCR control with water as template; 3-12, Transgenic-positive plants.
图2T0代转基因阳性植株的OsPMM基因(A)和Hpt基因(B)的PCR鉴定
Fig. 2. PCR analysis of the OsPMM gene(A) and Hpt gene (B) in T0transgenic-positive plants.
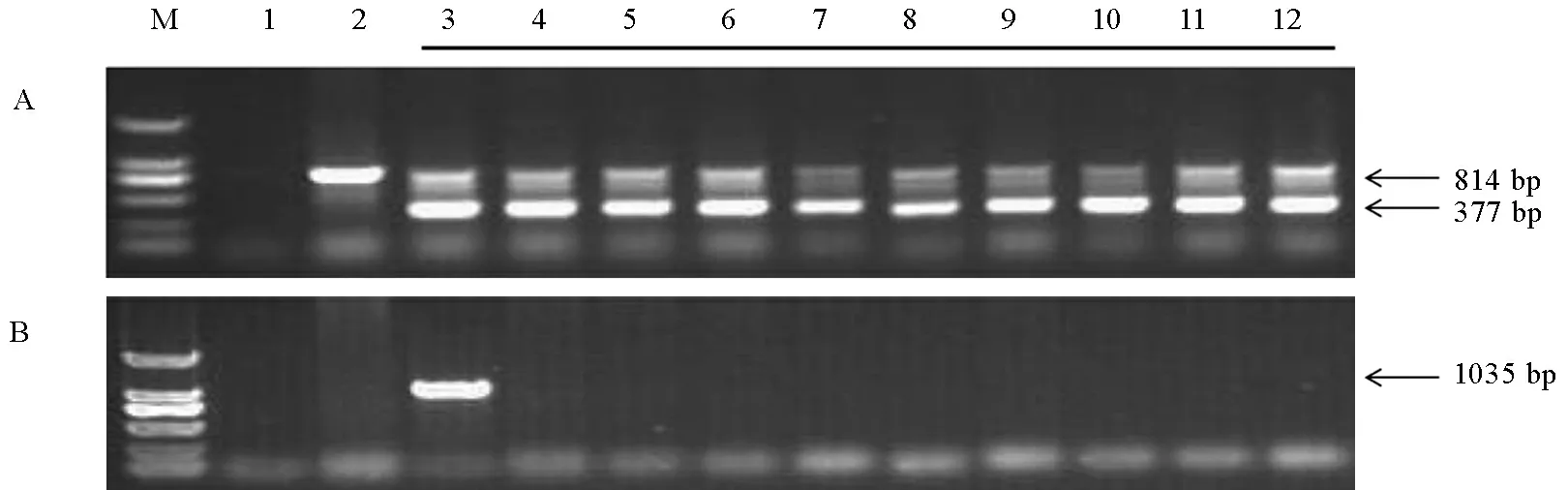
M-DL2000; 1-PCR体系对照; 2-非转基因对照; 3-转基因阳性对照; 4~12-部分T2代转基因纯合单株。
M, DL2000; 1, PCR control with water as template; 2, Transgenic-negative control; 3, Transgenic-positive control; 4-12, Partial transgenic T2plants.
图3无标记转基因系2-1部分T2代单株的OsPMM基因(A)和Hpt基因(B)的PCR鉴定
Fig. 3. PCR analysis of the OsPMM gene (A) and Hpt gene (B) in partial T2progenies of marker-free transgenic line 2-1.
2.2OsPMM基因在转基因纯合系种子部位的表达水平明显提高
待种子成熟时,单株套袋收获以上T2代转基因纯合系的种子。待种子晒干后,随机抽取部分种子,提取种子部位的RNA,以Actin基因为内参,以转基因受体C418品种为对照,对OsPMM基因在3个无标记转基因系2-1,3-1和7-1中的表达进行相对定量分析,发现OsPMM在转基因系中的表达水平有明显的提高(图 4)。
2.3转基因纯合系种子中的VC含量显著提高
OsPMM基因在转基因纯合系种子中的VC转录水平显著提高,为了检验转基因系种子部位的VC含量是否提高,我们将对照系和3个转基因系的种子部位的总VC含量进行了测定。对检测结果进行统计分析发现,对照C418的VC含量约8 μmol/g,而转基因系的VC含量为10~12 μmol/g,与对照C418相比,3个转基因纯合系种子中的VC含量提高了25%~50%(图 5)。
通过对OsPMM在转基因系种子中RNA水平的相对表达分析,以及种子部位的VC含量测定,发现调节OsPMM基因的转录水平可以影响水稻种子中的VC含量。本研究结果表明,在水稻中,通过提高OsPMM基因的表达水平来增加水稻种子的VC含量是行之有效的。
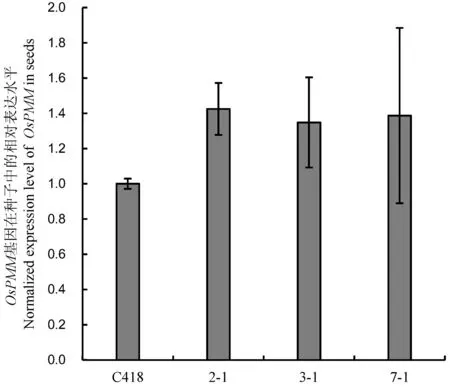
2-1,3-1和7-1为转基因纯合系; C418为转基因阴性对照。
2-1, 3-1 and 7-1, Transgenic homozygous lines; C418, Transgenic-negative control.
图4OsPMM基因在转基因纯合系种子中的相对表达水平
Fig. 4. Relative expression level of OsPMM gene in seeds of transgenic homozygous lines.

2-1,3-1和7-1为转基因系; C418-转基因阴性对照。
2-1, 3-1 and 7-1, Transgenic lines; C418, Transgenic-negative control.
图5转基因纯合系种子中的VC含量分析
Fig. 5. VC content in seeds of transgenic homozygous rice lines.
3讨论
水稻作为重要的粮食作物,是我国南方地区食用的主食。VC是植物体内重要的抗氧化剂[21],也是人体每天需求量最多的维生素。人类不能合成VC,需要从每天的膳食中来摄取VC,因而提高水稻的VC含量,对补充人体的VC有很大帮助。对植物PMM基因的转基因操作发现,通过调节植物中PMM基因的表达,可以改变受体的VC含量。在烟草和拟南芥中过表达PMM都能提高受体的VC含量[14];在烟草中过表达来自樱桃的PMM基因,也能提高受体VC的含量[22]。对于水稻而言,水稻种子是食用的部分, 对水稻的改良重点应是提高水稻种子的VC含量。为了特异地提高水稻种子的VC含量,我们采取了构建融合基因的策略,使用种子特异表达的启动子Bx14驱动OsPMM基因的表达。改造好的基因构建到表达载体上,通过转基因的途径转入到水稻品种中。结果和我们预期一致,OsPMM基因在转基因水稻种子中的表达量相对于对照明显提高,相应地,水稻种子中的VC含量也有一定程度的提高。
在本研究中,我们之所以通过转基因的途径将OsPMM基因转入到C418水稻品种中,一是要使OsPMM基因在种子中特异高表达,使用种子特异表达的启动子驱动OsPMM基因的表达是最有效途径,这需要对OsPMM基因进行基因工程改造,然后通过转基因技术转入到受体水稻中;二是转基因技术是进行作物改良的一个有效途径,相比较于传统的回交转育育种技术,转基因技术的一个明显优势是特异地将含有目标基因的T-DNA区转入到受体中,减少了“连锁累赘”对目标性状的影响,能够在短时间内(一到两年)获得除目标性状外,其他性状和受体保持一致的改良系[23]。三是对转基因和回交转育技术选育的玉米、小麦和水稻等作物的研究发现,在转录组学水平上,通过这两种技术选育的植物具有实质等同性,转基因技术并不会比传统的回交育种技术带来更多的基因表达变化[24-26]。本研究通过对T0、T1和T2代转基因水稻进行连续的分子鉴定,在T2代便筛选得到了主要农艺性状和对照一致且种子VC含量提高的转基因纯合系。由于在构建转基因表达载体上采用了双右边界双元载体作为骨架,所筛选到的转基因纯合系无选择标记基因。另外,目标基因OsPMM来源于水稻,其所用种子特异表达启动子来自于禾本科植物小麦。因此,本研究所得到的高VC含量的转基因水稻在育种实践中具有重要的应用价值。
参考文献:
[1] Wheeler G L, Jones M A, Smirnoff N. The biosynthetic pathway of vitamin C in higher plants.Nature, 1998, 393(6683): 365-369.
[2] Smirnoff N, Wheeler G L. Ascorbic acid in plants: Biosynthesis and function.CritRevBiochemMolBiol, 2000, 35(4): 291-314.
[3] Smirnoff N, Conklin P L, Loewus F A. Biosynthesis of ascorbic acid in plants:A renaissance.AnnuRevPlantPhysiolPlantMolBiol, 2001, 52: 437-467.
[4] Wolucka B A,van Montagu M. GDP-mannose 3′,5′-epimerase forms GDP-L-gulose, a putative intermediate for the de novo biosynthesis of vitamin C in plants.JBiolChem, 2003, 278(48): 47483-47490.
[5] Lorence A, Chevone B I, Mendes P, et al. Myo-inositol oxygenase offers a possible entry point into plant ascorbate biosynthesis.PlantPhysiol, 2004, 134(3): 1200-1205.
[6] Agius F, Gonzalez-Lamothe R, Caballero J L, et al. Engineering increased vitamin C levels in plants by overexpression of a D-galacturonic acid reductase.NatBiotechnol, 2003, 21(2): 177-181.
[7] Keller R, Renz F S, Kossmann J. Antisense inhibition of the GDP-mannose pyrophosphorylase reduces the ascorbate content in transgenic plants leading to developmental changes during senescence.PlantJ, 1999, 19(2): 131-141.
[8] Gatzek S, Wheeler G L, Smirnoff N. Antisense suppression ofL-galactose dehydrogenase inArabidopsisthalianaprovides evidence for its role in ascorbate synthesis and reveals light modulatedL-galactose synthesis.PlantJ, 2002, 30(5): 541-553.
[9] Tabata K, Oba K, Suzuki K, et al. Generation and properties of ascorbic acid-deficient transgenic tobacco cells expressing antisense RNA forL-galactono-1,4-lactone dehydrogenase.PlantJ, 2001, 27(2): 139-148.
[10]Tokunaga T, Miyahara K, Tabata K, et al. Generation and properties of ascorbic acid-overproducing transgenic tobacco cells expressing sense RNA forL-galactono-1,4-lactone dehydrogenase.Planta, 2005, 220(6): 854-863.
[11]Jain A K, Nessler C L. Metabolic engineering of an alternative pathway for ascorbic acid biosynthesis in plants.MolBreed, 2000, 6(1): 73-78.
[12]李坤岚. 转小鼠古洛糖酸内酯氧化酶基因拟南芥的评估. 上海: 复旦大学, 2010.
Li K L. Evaluation of mouse gulonolactone oxidase gene transformedArabidopsisthaliana. Shanghai: Fudan University, 2010.
[13]Chen Z, Young T E, Ling J, et al. Increasing vitamin C content of plants through enhanced ascorbate recycling.ProcNatlAcadSciUSA, 2003, 100(6): 3525-3530.
[14]Qian W, Yu C, Qin H, et al. Molecular and functional analysis of phosphomannomutase (PMM) from higher plants and genetic evidence for the involvement of PMM in ascorbic acid biosynthesis inArabidopsisandNicotianabenthamiana.PlantJ, 2007, 49(3): 399-413.
[15]Badejo A A, Eltelib H A, Fukunaga K, et al. Increase in ascorbate content of transgenic tobacco plants overexpressing the acerola (Malpighiaglabra) phosphomannomutase gene.PlantCellPhysiol, 2009, 50(2): 423-428.
[16]Lu H J, Zhou X R, Gong Z, X, et al. Generation of selectable marker-free transgenic rice using double right-border (DRB) binary vectors.AustJPlantPhysiol, 2001, 28(3): 241-248.
[17]杨振玉, 张宗旭, 魏耀林, 等. 粳型特异亲和恢复系 C418 的选育及其特性. 杂交水稻, 1998, 13(3): 31-32.
Yang Z Y, Zhang Z X, Wei Y L, et al. Breeding and characteristics of japonica type wide compatibility line C418.HybridRice, 1998, 13(3): 31-32.
[18]Zhai W X, Li X B, Tian W, et al. Introduction of a rice blight resistance gene,Xa21, into five Chinese rice varieties through anAgrobacterium-mediated system.ChinaC:LifeSci, 2000, 43(4): 361-368.
[19]Livak K J, Schmittgen T D. Analysis of relative gene expression data using rea L-time quantitative PCR and the 2-ΔΔCTmethod.Methods, 2001, 25(4): 402-408.
[20]Gillespie K M, Ainsworth E A. Measurement of reduced, oxidized and total ascorbate content in plants.NatProtocols, 2007, 2(4): 871-874.
[21]Noctor G, Foyer C H. Ascorbate and glutathione: Keeping active oxygen under control.AnnuRevPlantPhysiolPlantMolBiol, 1998, 49: 249-279.
[22]Badejo A A, Eltelib H A, Fukunaga K, et al. Increase in ascorbate content of transgenic tobacco plants overexpressing the acerola (Malpighiaglabra) phosphomannomutase gene.Plant&CellPhysiol, 2009, 50(2): 423-428.
[23]Gao L, Xia Z, Jiang G, et al. Generation of marker-free, bacterial blight-resistant transgenic sterile line and hybrid rice withXa21.PlantBreed, 2011, 130(4): 438-443.
[24]Gao L, Cao Y, Xia Z, et al. Do transgenesis and marker-assisted backcross breeding produce substantially equivalent plants? A comparative study of transgenic and backcross rice carrying bacterial blight resistant geneXa21.BMCGenom, 2013, 14: 738-750.
[25]Baudo M M, Lyons R, Powers S, et al. Transgenesis has less impact on the transcriptome of wheat grain than conventional breeding.PlantBiotechnolJ, 2006, 4(4): 369-380.
[26]Coll A, Nadal A, Collado R, et al. Gene expression profiles of MON810 and comparable non-GM maize varieties cultured in the field are more similar than are those of conventional lines.TransgenicRes, 2009, 18(5): 801-808.
收稿日期:2015-08-21; 修改稿收到日期: 2015-10-20。
基金项目:国家转基因生物新品种培育重大专项(2014ZX08001-002); 国家自然科学基金资助项目(31300999); 湖北省教育厅项目(B2015229); 武汉市科学技术局项目(2014072704011250); 江汉大学科研启动项目(3003-06000043)。
中图分类号:Q786; S511.0353
文献标识码:A
文章编号:1001-7216(2016)04-0441-06
TransgenosisofthePhosphomannomutaseTransgeneIncreasesVitaminCContentinRice
GAOLi-fen1, 2,XIAZhi-hui2, 3,ZHANGJi1,WANGDao-wen2,ZHAIWen-xue2,*
(1InstituteforSystemsBiology,JianghanUniversity,Wuhan430056,China;2InstituteofGeneticsandDevelopmentalBiology,ChineseAcademyofSciences,Beijing100101,China;3InstituteofLifeSciences,HainanUniversity,Haikou570228,China;*Correspondingauthor,E-mail: wxzhai@genetics.ac.cn)
Abstract:VC (vitamin C) is an essential nutrient to human health. Due to lack of L-gulonolactone oxidase, the last enzyme involved in VC synthesis pathway, human could not synthesize VC by themselves. Rice is an important food crop and its nutritional value could be greatly improved by increasing the VC content in rice seeds. Phosphomannomutase (PMM) is an important enzyme in VC synthesis pathway, catalyzing the interconversion from mannose 6-phosphate from mannose 1-phosphate. In this study, Oryza PMM gene (OsPMM) that was under the control of seed-specific expressed promoter Bx14 was transferred into ‘C418’, a restorer line of three-line japonica hybrid rice, using the double right-border vector pMNDRBBin6 through an Agrobacterium tumefaciens-mediated system. Molecular analysis revealed that OsPMM was integrated into the genome of transgenic ‘C418’, and the homozygous and marker-free transgenic line was obtained in the T2 generation. Gene expression analysis of transgenic lines showed the expression level of OsPMM was significantly increased in seeds, and accordingly, the VC content in the seeds of transgenic plant also increased by 25-50%.
Key words:vitamin C; phosphomannomutase; double right-border vector system; marker-free; transgenic line
