赤水桫椤国家级自然保护区桫椤群落特征及物种多样性研究
2016-08-06宗秀虹张华雨李宗峰吴洪英邓洪平
宗秀虹,张华雨,王 鑫,李宗峰,吴洪英,梁 盛,邓洪平*
(1 西南大学 生命科学学院,三峡库区生态环境教育部重点实验室,重庆 400715;2 贵州赤水桫椤国家级自然保护区管理局,贵州赤水 564700)
赤水桫椤国家级自然保护区桫椤群落特征及物种多样性研究
宗秀虹1,张华雨1,王鑫1,李宗峰1,吴洪英2,梁盛2,邓洪平1*
(1 西南大学 生命科学学院,三峡库区生态环境教育部重点实验室,重庆 400715;2 贵州赤水桫椤国家级自然保护区管理局,贵州赤水 564700)
摘要:桫椤(Alsophila spinulosa),为中生代古老孑遗物种,国家二级保护植物。贵州赤水桫椤国家级自然保护区是以桫椤及其生存环境为保护对象的自然保护区,是拥有世界上数量最多、面积最广的桫椤林区之一。该文采用植物生态学、区系学等方法,对保护区内群落及物种多样性、伴生物种区系成分、生活型谱、群落结构等进行研究。结果表明:(1)群落内维管植物共计60科96属122种,其中蕨类植物12科14属17种,裸子植物2科2属2种,被子植物46科80属103种。(2)群落内植物的科、属区系成分类型多样,热带成分占优势,温带成分次之,表现出从热带区系向温带区系过渡的特征。(3)生活型谱以高位芽植物为主(占57.38%),反映出中亚热带森林以高位芽植物为优势的特点。(4)群落垂直分层显著,乔木层树高在4~12 m的比例最大为76.64%;灌木层树高在2~3 m的比例最大为69.78%;草本层各区间分布较为均匀,但以0~0.2 m的比例较大,群落层次明显。(5)毛竹(Phyllostachys heterocycla cv. pubescens)和桫椤在该地乔木组成中占主导地位;灌木层中,桫椤为最优物种;草本层主要以红盖鳞毛蕨(Dryopteris erythrosora)、翠云草(Selaginella uncinata)为主;物种丰富度分析显示,乔木层<灌木层<草本层;多样性指数、均匀度指数分析表明,乔木层为聚集分布、组成较简单,灌木层和草本层为均匀分布,但数量较少。
关键词:桫椤群落;物种多样性;植物区系;生活型;群落多样性
桫椤为中生代古老孑遗物种[1],为国家二级重点保护野生植物[2],是地球历史变迁的见证,更是科学研究的活化石[3],对研究古地质气候、古环境的演变,探索生物进化的奥秘,保证遗传物种生态系统的持续利用具有重要的保护及学科研究价值。目前,对其生态学方面的研究主要集中在种群结构及特征、分布格局、种内种间竞争及联结、生态位、物种多样性等方面[4-10]。赤水桫椤国家级自然保护区始建立于1984年,1992年经国务院批准为国级自然保护区,次年被纳为中国生物圈保护区,是以桫椤及其生存环境为保护对象的的自然保护区,拥有世界上数量最多、面积最广的桫椤林区之一。保护区内特殊的低海拔沟谷地貌,良好的水热条件、充沛的年降水量,较高的相对湿度、深厚的土壤是桫椤群落生长的绝佳环境。但近年来由于人为因素的影响,和其他物种如毛竹与桫椤在空间、水分、阳光等方面的竞争,导致桫椤的生存受到了较大的威胁[3]。笔者对赤水桫椤国家级自然保护区的桫椤群落进行详实调查,通过对桫椤群落的物种多样性、区系特征、生活型谱、多样性分析,并与前人研究[9]做对比,进一步探讨桫椤群落特征及多样性,以期为赤水桫椤国家级自然保护区桫椤群落的保护及其可持续发展等提供参考资料和依据。
1研究区域自然概况
赤水桫椤国家级自然保护区位于贵州省赤水中部葫市镇金沙沟,E105°57′54″~106°7′7″,N28°20′19″~28°28′40″,总面积为13 300 hn2。保护区属中亚热带湿润季风气候,河谷具有类似南亚热带气候特征[1]。区内气候特点为冬无严寒,夏无酷暑,日照少,温度高,湿度大,降水充沛,云雾雨日多,垂直差异大。保护区的地势东南高,西北低,河流在流经的过程中,对地表进行了强烈的侵蚀和切割,导致保护区内主要河流支流众多。土壤多为中性和微酸性的砂质紫色土,主要为非地带性的紫色土,发育成熟。植被类型多样,以亚热带湿润常绿阔叶林为主,还包括暖性针叶林、落叶阔叶林、常绿落叶阔叶混交林、常绿阔叶林、竹林及常绿阔叶灌丛等多种植被类型。
2研究方法
2.1样地设置及群落调查
2013年至2015年多次对贵州赤水桫椤国家级保护区桫椤群落进行野外调查,采用典型样地调查法,考虑到林分类型、地形等因素,在保护区内金沙沟、甘沟、南厂沟等处的生态旅游景区,选择代表性地段设置样方。样地设置在保护区内人为干扰较少和外貌整齐的桫椤林中,共设9个20 m×20 m样地。在每个样地内再设16个5 m×5 m的样方,样方中所有物种每木必查,高于3 m的立木记为乔木,其余记为灌木,并记录物种名称、株高、胸径、冠幅等;在样地的对角,分别设1个1 m×1 m的草本样方,记录草本样方中的物种数及每个物种的平均高度、盖度及物种名称等,并对样地的郁闭度、海拔、坡度、坡位等进行调查记录。
2.2区系分析
参照臧得奎[11],秦仁昌[12]对中国蕨类植物科属的地理成分划分系统,李锡文[13]关于中国种子植物科的分布区类型划分系统,吴征镒[ 14]种子植物属的分布区类型划分方法,对样地内出现的所有植物进行统计分析。
2.3生活型分析
根据Raunkiaer[15]的生活型分类系统对桫椤群落的生活型谱进行分析。
2.4数据处理
重要值计算按照文献[4-5],计算,公式为:
重要值=(相对多度+相对频度+相对显著度)/3
物种多样性分析按文献[16]进行。
物种丰富度(S),即样方内的物种数。
Simpson优势度指数(D):
D=1-∑Pi2
Shannon-wiener多样性指数(H′):
H′= -∑PilnPi
Pielou均匀度指数(E):
E=H/Hmax
式中,S为群落中的总物种数目;Pi=Ni/N,Pi为属于种i的重要值(经典公式的Pi为属于种i的个体在全部个体中的比例。Ni为种i的个体数,N为群落中全部物种的个体数;H为实际观察的物种多样性指数,Hmax为最大的物种多样性指数,Hmax=LnS(S为群落中的总物种数)。
3结果与分析
3.1赤水桫椤群落中植物种类组成分析
经调查统计,赤水桫椤保护区桫椤群落共有维管植物共计60科96属122种,其中蕨类植物12科14属17种,种子植物48科82属105种;种子植物中裸子植物2科2属2种,被子植物46科80属103种,被子植物中双子叶植物38科63属82种,单子叶植物8科17属21种(表1)。双子叶植物中,山茶科(Theaceae)包含的植物种类最多,共7种,大戟科(Euphorbiaceae)和桑科(Moraceae)各含5种。在单子叶植物中,禾本科(Gramineae)包含了9种,其余各科均含有1~3种不等。双子叶植物作为主要成分,占总种数的67.22%,而蕨类植物、裸子植物单子叶植物则分别占总种数的13.93%、1.64%、17.21%。
3.2桫椤群落植物区系组成
赤水桫椤群落中有维管植物60科96属122种,分别占赤水桫椤保护区维管植物总科数的31.09%、总属数的12.18%、总种数的6.05%。根据臧得奎[11]对中国蕨类植物科属的地理成分的划分系统,秦仁昌分类系统[12],李锡文[13]关于中国种子植物分布区类型划分系统,吴征镒等[14]对中国种子植物属的分布区类型划分方法,对本研究区域桫椤群落中植物进行统计分析结果见表2。
科的类型分析结果显示,赤水桫椤群落中含中等科组成了本区系科的主体,共计50科,占本地区植物总科数的83.33%;从科的水平上看,桫椤群落植物科分为8个分布区类型,热带分布(表2,2~7型) 39科,其中较大的科有茜草科(Rubiaceae),较小的科有芭蕉科(Musaceae)等,温带分布(表2,8~14型)类型10科,分别占桫椤群落中植物非世界分布科的79.59%、20.41%;可见本研究区植物科的分布类型具有明显热带区系性质的同时,也有向温带过渡的趋势。这与保护区地处亚热带山区的地理位置相符[1]并在一定程度上也反映了赤水桫椤群落植物的多样性和植物区系的复杂性。
属的统计结果显示,群落内植物共计96属,少种属[17]所占比例最大,共41属,占总属数的42.71%,少种属包含植物45种,占桫椤群落内植物总数36.89%;其次是中等属,共25属,占总属数的26.04%,包含植物29种,占桫椤群落内植物总数23.77%。从属的水平上看,群落内96属分属于10种类型(表2),体现了该区系地理成分的复杂性。本研究区植物区系中主要以泛热带及其变型分布为主,占桫椤群落植物总属数(不含世界分布)的31.03%。热带分布(2~7型),共67属,占桫椤群落植物总属数(不含世界分布)的77.01%,如:狗尾草属(Setaria)、菝葜属(Smilax)、山矾属(Symplocos)等;温带分布(表2,8~14型)共计19属,占桫椤群落植物总属数(不含世界分布)21.84%,如:松属(Pinus)、鸭儿芹属(Cryptotaenia)、栎属(Quercus)等。从属的水平来看,其温热比值(R/T)为0.28,热带成分占优势,同样表明该研究区域分布类型具有明显热带区系性质的同时,也有向温带过渡的趋势。
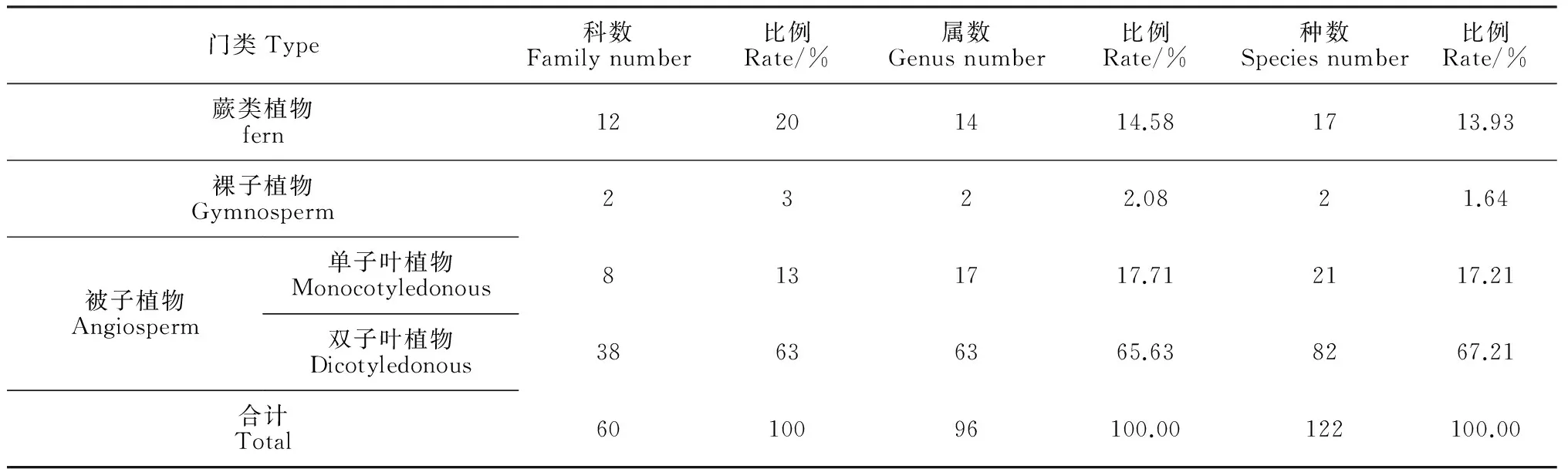
表1 赤水桫椤群落植物物种组成统计表

表2 赤水桫椤群落植物科、属的分布区类型
3.3生活型分析
依照Raunkiaer[15]提出的生活型分类系统,对赤水桫椤保护区桫椤群落122种植物的生活型进行统计分析结果(图1)显示,122种植物中,高位芽植物种类最多,有70种,占总种数的57.38%,地上芽植物有23种,占总种数的18.85%,地下芽植物有22种,占总种数的18.03%,种类较少的是地面芽(3 种) 和一年生(4种) 植物,分别占总种数的2.46%和 3.28%。高位芽植物中,大高位芽植物缺乏,林下灌木层物种丰富,小高位芽(43种) 、矮高位芽(23种) 植物较多,而中高位芽数量较少(4种),该生活型谱基本反映出中亚热带森林群落以高位芽植物为优势的生活型谱的基本特征,同时,高位芽以小、矮高位芽植物为优势也反映出赤水桫椤群落生活型谱以乔木层优势种为主、林下灌草层物种组成丰富的特点。
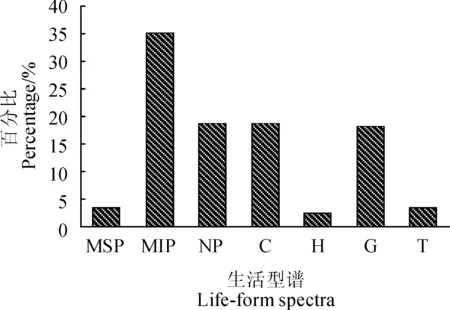
MSP.中高位芽;MIP.小高位芽;NP.矮高位;C.地上芽;H.地面芽;G.地下芽;T.一年生图1 赤水桫椤群落生活型谱MSP is mesophanerophyte; MIP is microphanerophyte; NP is nanophanerophyte; C is chamaephyte; H is hemicryptophyte; G is geophyte; T is therophyte.Fig. 1 Life-form spectra of A. spinulosa community in Chishui
3.4赤水桫椤群落物种的植株高度
群落的垂直结构可用不同高度下植株的株数比例来表示,从表3可见,乔木层树高在4~8 m的比例最大,达39.6%;灌木层树高在2~3 m比例最高,达69.78%;草本层各区间分布较为均匀,以0~0.2 m的比例较大,为30.98%,其余相差不大。
3.5赤水桫椤群落物种的直径
以直径小于2 cm为一个径级,其余以4 cm为一个径阶,对赤水桫椤群落的乔木层径级结构进行分析,结果(图2)显示,较小径级(0~14 cm)株数比例为87.5%,中等径级(14~26 cm)株数比例为11.83%,较大径级(>18 cm),株数比例只有0.67%,表明该类型群落乔木层中,中小径级的植株所占比例较大,具有一定的优势,整个趋势为“J”型。

表3 赤水桫椤群落各物种结构层的植株高度及其分布
以2 cm为一个径阶株数分布状况,对赤水桫椤群落的灌木层直径结构进行分析结果(图3)显示,该类型群落灌木层中较小径级(<2 cm)的株数比例为38.05%,中等径级(2~8 cm)的株数比例为46.46%,较大径级(>8 cm),株数比例只有15.49%。由此可见,该类型群落灌木层中,分布较为均匀。
3.6群落物种多样性分析
重要值和多样性的计算是植物群落特征研究的重要参数,通过计算桫椤群落中乔木层、灌木层和草本层各物种的重要值,分析各物种在群落中的地位及对物种多样性的贡献,结果(表4)显示,赤水桫椤群落的乔木层结构比较简单,重要值大于10%的只有毛竹和桫椤,分别为0.551、0.147,在该地组成中占主导地位,其他伴生物种如:芭蕉、粗叶木、罗伞等在该区域重要值均较低,影响较小。
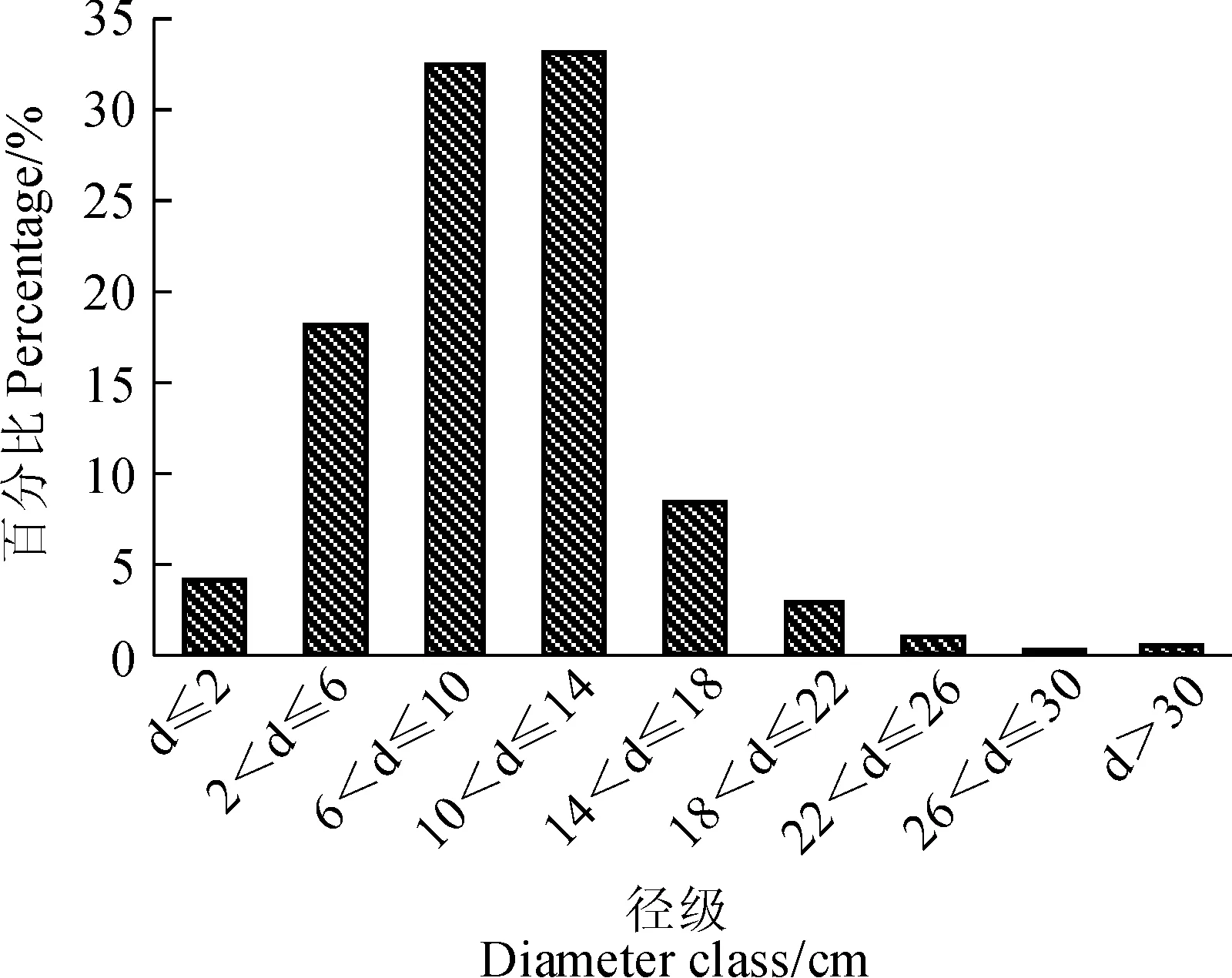
图2 赤水桫椤群落乔木层的径级分布结构Fig. 2 Diameter class structure and distribution of arbor layer of A. spinulosa community in Chishui
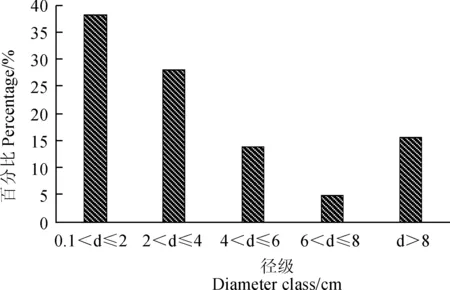
图3 赤水桫椤群落灌木层的径级分布结构Fig. 3 Diameter class structure and distribution of shrub layer of A. spinulosa community in Chishui
赤水桫椤群落灌木层种类较丰富(表4),桫椤的重要值高达0.326,是该群落灌木层的优势种和建群种,占明显的优势地位。芭蕉(Musabasjoo)、毛竹、脚骨脆(Caseariabalansae)、罗伞(Brassaiopsisglomerulata)的重要值也较大,这4种植物是该群落灌木层的主要成分,也是乔木层的更新物种,其他物种如:粗叶木、金珠柳(Maesamontana)、毛桐(Mallotusbarbatus)等均为伴生种。调查发现,该区域水热湿度条件良好、土壤深厚、呈弱酸性,且生境较为隐蔽,公路山坡上及河岸沟谷边较多,是桫椤生长的绝佳环境,这也是其能在灌木层中成为最优物种的优势。
草本层的种类丰富,重要值较大的有红盖鳞毛蕨、翠云草以及竹叶草(Oplismenuscompositus),它们在草本层占有明显优势,其它伴生种有卷柏(Selaginellatamariscina)、楼梯草(Elatostemainvolucratum)等,重要值较低,草本层在群落内的物种较多,但多度较低,可能是由于群落冠层郁闭[18]较大,造成林下草本植物光照较少,影响生长,且群落内较高的空间异质性也增加了群落物种的丰富度。
群落物种多样性既是一个群落或生境中物种的丰富度,变化程度和均匀度的指标,也是群落生态组织水平可测的重要生物学特征。对群落中乔木、灌木、草本3层的物种多样性分析结果(图4)表明,该群落各层次物种丰富度表现为:草本层>灌木层>乔木层,而多样性指数、匀度指数则表现为: 草本层>灌木层>乔木层,表明乔木层为聚集分布,物种组成较少,组成简单,灌木层和草本层的物种数目远超过乔木层,但个体数量不多,呈均匀分布。

表4 赤水桫椤群落乔木层、灌木层和草本层物种重要值
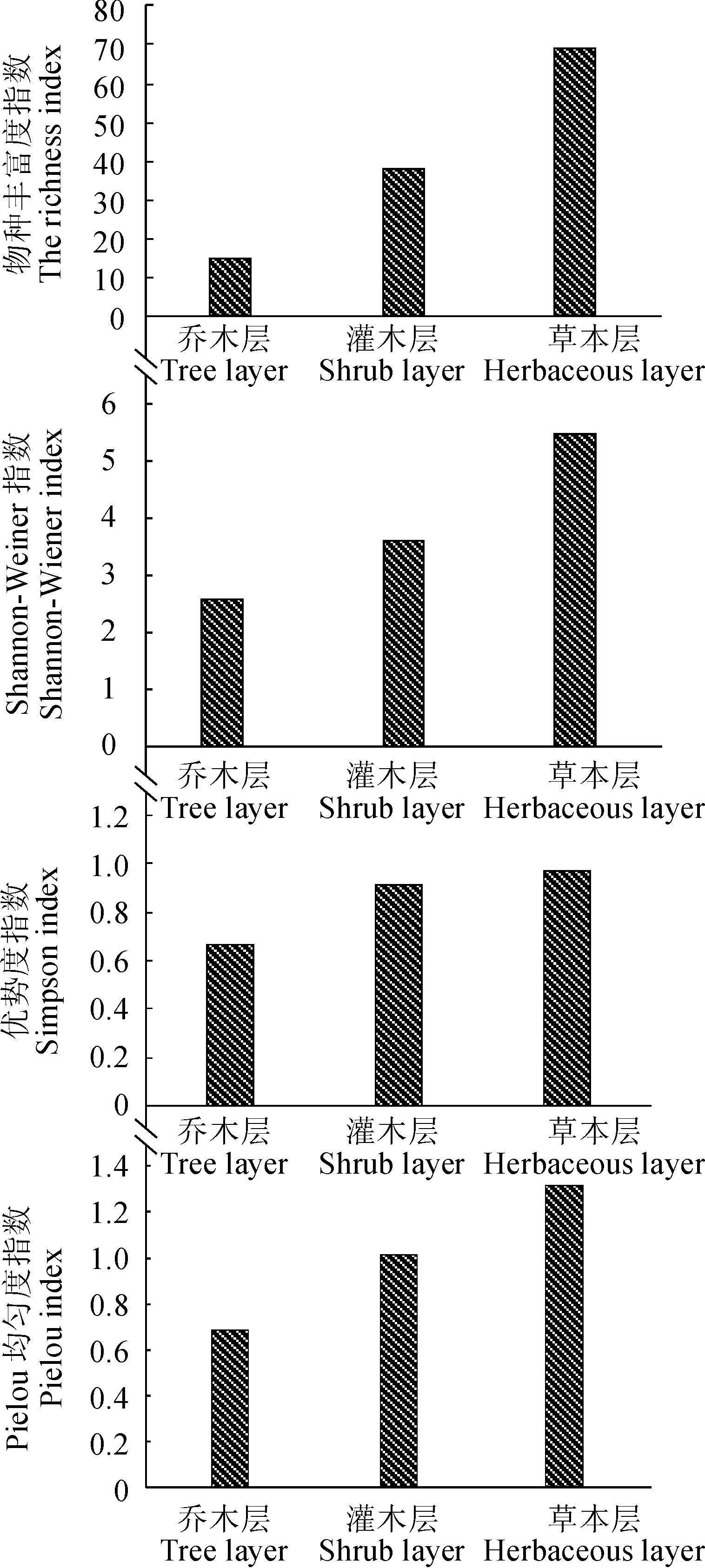
图4 赤水桫椤群落物种多样性指数Fig. 4 Species diversity of A. spinulosa community in Chishui
4讨论
贵州赤水桫椤群落的物种组成丰富,有维管植物共60科90属 122种,其中蕨类植物12科13属17种,种子植物48科82属105种;据统计,在群落中,山茶科、桑科、大戟科等包含的植物种类最多,但却在群落重要值计算中不占优势,分析原因是其群落中山茶科、桑科、大戟科植物几乎全为矮小乔木。
本研究区群落区系特点表明其植物地理成分复杂,在15种中国植物科属的分布区类型中存在有8种科的分布区类型、10种属的分布区类型,其中热带分布类型占明显优势,但温带分布类型在群落物种组成中也占较大的比例,这与其特殊的地理条件即地处中亚热带范围,地形复杂及小生境多样相关,与屠玉林[9]1990年对贵州桫椤群落的初步研究结果一致。说明,该地区区系特征具有由热带向温带过渡的性质[1]。
赤水桫椤群落的生活型主要以高位芽植物为主,所占比例为57.38%,高位芽植物中只有矮高位芽、小高位芽和中高位芽植物,没有大高位芽植物,这主要由于生态旅游和早前人为种植的毛竹、慈竹,导致高位芽植物很难长到25 m以上,造成大高位芽植物的缺乏[19]。研究表明,热带、亚热带的常绿阔叶林植物生活型主要为高位芽植物,地下芽植物较少,而温带地区因气候条件,植物生活型谱中以地面芽和地下芽植物占优势[20],而该群落高位芽植物占优势,地上芽和地下芽分别占18.85%、18.03%,反映了桫椤群落具有明显的亚热带常绿阔叶林的外貌特征,地下芽植物主要是蕨类植物、姜属植物等,也反映出桫椤群落具有南亚热带特色。
从群落的垂直结构来看,可分为乔木层、灌木层和草本层。乔木层树种多在4~16 m之内,灌木层树种多在2~3 m内,草本层各区间分布较为均匀,但以0~0.2 m的比例较大。通过对该群落的径级株数的研究表明,乔木层胸径为小于14 cm的树种占绝大多数,比例高达87.5%,其发展趋势呈倒J形状。可见群落中,中小径阶直径的植物比例较高,但也不乏高大树种,像优势树种桫椤,最大的胸径可达28 cm,这也是其重要值高的原因之一。
物种丰富度与多样性指数是群落稳定性的重要指标,能客观反映群落的物种丰富度和均匀度,群落结构越复杂[21]。从群落物种多样性分析表明,群落多样性不高,主要是草本层包含更多的物种,乔木层和灌木层相对较少。特殊的气候条件、生境以及人为干扰、毛竹入侵等造成该地区物种多样性较低。而多样性指数、均匀度指数都表现为: 草本层>灌木层>乔木层,表明乔木层为聚集分布,物种组成较少,灌木层和草本层的物种数目较多于乔木层,但个体数量不多,呈均匀分布。这可能是由于群落冠层郁闭较大,造成林下草本植物光照较少,影响生长,同时,群落内较高的空间异质性增加了群落物种的丰富度[21]。
综合分析桫椤群落后发现,部分样地桫椤已退居次优势种的地位,其重要值落后于毛竹,且桫椤群落附近还有人为种植经营管理的成片毛竹林,毛竹无论是在数量上或者质量上都占有更重要的建群地位。如果让其以强盛的地下茎分蘖生长方式来保持其数量和质量上的优势,那么桫椤的生长发育将会受到抑制,而退出优势种的地位,最终桫椤群落解体[22]。
依据对赤水桫椤群落特征及物种多样性的分析,在了解桫椤群落特征和物种多样性的基础上,现提出以下建议:人为干扰是影响桫椤群落生长重要因素,部分区域由于毛竹扩张,尤其是当研究区内的湿度、温度等条件受到外界较大的干扰时,桫椤孢子的萌发可能会受到限制,进而影响到桫椤种群的繁衍。所以保护区应适度控制在野生桫椤种群的旅游观光等人为活动,并建立生态监护站,桫椤群落的永久性样地等对群落的种类、外貌及动态演替规律等进行观测,加强对保护区竹林的合理控制与管理,杜绝桫椤林下苔藓、地衣层被破坏,确保桫椤孢子体能正常萌发。
参考文献:
[1]王密, 屠玉麟, 何谋军. 赤水桫椤自然保护区植物和植被多样性现状及特点分析[J]. 贵州师范大学学报:自然科学版, 2005, 23(1):19-22.
WANG M,TU Y L,HE M J. The present statues and characteristics analysis of the plants and vegetation diversity in the Chishui alsophila Spinulosa Nature Reserve[J].JournalofGuizhouNormalUniversity(NaturalSciences), 2005, 23(1):19-22.
[2]中国科学院《中国植物志》编辑委员会. 中国植物志(1-80卷)[M]. 北京: 科学出版社, 1979.
[3]郝云庆, 江洪, 余树全,等. 桫椤植物群落区系进化保守性[J]. 生态学报, 2009, 29(8):4 102-4 111.
HAO Y Q, JIANG H ,YU S Q,etal. Evolutionary donservation of floristic characteristics of relictAlsophilaspinulosa community[J].ActaEcologicaSinica, 2009, 29(08):4 102-4 111.
[4]徐德静, 王鹏鹏, 何跃军,等. 黔北丹霞地貌桫椤群落优势种群生态位研究[J]. 植物研究, 2014,(5):612-618.
XU D J, WANG P P, HE Y J,etal. Niche characteristics of dominantplantpopulations ofAlsophilaspinulosacommunity of Danxia Landform in North Guizhou[J].BulletinofBotanicalReseach, 2014,(5):612-618.
[5]宋萍, 洪伟, 吴承祯,等. 珍稀濒危植物桫椤种群结构与动态研究[J]. 应用生态学报, 2005, 16(3):413-418.
SONG P,HONG W,WU C Z,etal.Population structure and its dynanics of rare and endangered plantAlsophilaspinulosa[J].ChineseJournalofAppliedEcology, 2005, 16(3):413-418.
[6]石胜友, 郭启高, 成明昊,等. 涪陵磨盘沟自然保护区桫椤种群分布格局的分形特征[J]. 生态学杂志, 2005, 24(5):581-584.
SHI S Y, GUO Q G, CHENG M H,etal. Fractal properties of the spatial ofAlsophilaspinulosapopulation in Mopangou[J].ChineseJournalofEcology, 2005, 24(5):581-584.
[7]鞠文彬, 高信芬, 包维楷. 画稿溪国家级自然保护区珍稀植物桫椤种群结构与更新[J]. 植物科学学报, 2014, 32(2):113-121.
JU W B , GAO X F ,BAO W K . Population structure and regeneration of the endangered Tree FernAlsophilaspinulosain Huagaoxi Nature Reserve, Southwestern China[J].PlantScienceJourna, 2014, 32(2):113-121.
[8]石胜友, 成明昊, 郭启高,等. 涪陵磨盘沟桫椤种群格局的分形特征——信息维数[J]. 西北植物学报, 2004,24(7):1 179-1 183.
SHI SH Y ,CHENG M H ,GUO Q G,etal. Information dimension frantal properties of the spatial pattern ofAlsophilaspinulosa population in Mopangou, Fuling.[J].ActaBotanicaBoreali-OccidentaliaSinica, 2004,24(7):1 179-1 183.
[9]屠玉麟. 贵州桫椤群落的初步研究[J]. 植物生态学报, 1990, 14(2):165-171.
TU Y L. Apreliminarystudy on theAlsophilaspinulosacommunity in Guizhou Province[J].ActaPhytoecologicaetGeobotanicaSinica, 1990,(2):165-171.
[10]宋萍,洪伟, 吴承祯,等. 珍稀濒危植物桫椤种群生命过程及谱分析[J]. 应用生态学报, 2008, 19(12):2 577-2 582.
SONG P, HONG W, WU C Z,etal. Population life process and spectral analysis and endangered plantAlsophilaspinulosa[J].ChineseJournalofAppliedEcology, 2008, 19(12):2 577-2 582.
[11]臧得奎. 中国蕨类植物区系的初步研究[J]. 西北植物学报, 1998(3):459-465.
ZANG D K. A Preliminary study on the ferns Flora in China[J],ActaBotanicaBoreali-OccidentaliaSinica, 1998,(3):459-465.
[12]秦仁昌.中国蕨类植物科属系统排列与历史来源[J].植物分类学报, 1978, 16(3):1-19; 16(4):16-37.
QIN R CH. China fern families and genera constellations and historical sources[J].JournalofSystematicsEvolution, 1978, 16(3): 1-19; 16(4):16-37.
[13]李锡文. 中国种子植物区系统计分析[J]. 植物分类与资源学报, 1996, 18(4):363-384.
LI X W. Floristic statistics and analyses of seed plants form China[J].ActaBotanicaYunnanica, 1996,18(4):363-384.
[14]吴征镒. 中国种子植物属的分布区类型[J]. 植物分类与资源学报, 1991, 13(S4):1-139.
WU Z Y . Areal-types of genera of seed plant in China[J].ActaBotanicaYunnanica, 1991, 13(SIV):1-139.
[15]RUANKIAER C. The Life Forms of Plants and Statistical Plant Geography [M]. New York: Oxford University Press,1932: 2~104.
[16]张金屯. 数量生态学[M]. 北京: 科学出版社,2004.
[17]张巧明, 王得祥, 龚明贵,等. 秦岭火地塘种子植物区系及其与毗邻地区的关系分析[J]. 西北植物学报, 2012, 32(3):589-595.
ZHANG Q M , WANG D X ,GONG M G,etal. Flora of seed plant in huoditang and its relationship with floras in other areas of Qinling Mountains[J].BotanicaBoreali-OccidentaliaSinica, 2012,32(3):589-595.
[18]王永杰, 张首军. 不同郁闭度下天然白皮松林更新的研究[J]. 山西师范大学学报:自然科学版, 2008, 22(4):83-85.
WANG Y J , ZHANG S J ,The study of regeneration of naturaPinusbungeanain different forest canopy closure[J].JournalofShanxiNormalUniversity(Nat.Sci.Edi),2008, 22(4):83-85.
[19]高贤明, 陈灵芝. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J]. 植物学报, 1998,(6):553-559.
GAO X M, CHEN L Z. The revision of plant life-form system and an analysis of the life-form spectrum of porest plants in the warm temperate zone of China[J].ActaBotanicaSinica, 1998,(6):553-559.
[20]刘守江, 苏智先, 张璟霞,等. 陆地植物群落生活型研究进展[J]. 西华师范大学学报:自然科学版, 2003, 24(2):155-159.
LIU S J , SU Z X ,ZHANG J X,etal.Perspectives of the research on life form in land plant communities[J].JournalofSichuanTeachersCollege, 2003,24(2): 155-159.
[21]梁文斌, 谢碧霞, 巫涛,等. 南岳栓皮栎群落特征及多样性分析[J]. 中南林业科技大学学报:自然科学版, 2011, 31(9):55-59.
LIANG W B ,XIE B X ,WU T ,etal. Characteristics and species diversity analysis of Quercus variabilis community in Nanyue[J].JournalofCentralSouthUniversityofForestry&Technology,2011,31(9):55-59.
[22]邓洪平,王志坚,陶建平,等. 贵州赤水桫椤国家级自然保护区生物多样性 [M]. 北京:科学出版社,2015:6.
(编辑:潘新社)
文章编号:1000-4025(2016)06-1225-08
doi:10.7606/j.issn.1000-4025.2016.06.1225
收稿日期:2016-03-07;修改稿收到日期:2016-04-26
基金项目:科技部国家科技基础平台国家标本平台——教学标本子平台运行服务资助项目(2005DKA21403-JK)
作者简介:宗秀虹(1991-),女,硕士研究生,主要从事植物系统进化与保护生物学研究。E-mail:zxhong531@126.com *通信作者:邓洪平,博士,教授,主要从事植物系统进化与保护生物学研究。E-mail:denghp@swu.edu.cn
中图分类号:Q948.15+7
文献标志码:A
Community Characteristics and Species Diversity ofAlsophilaspinulosain Chishui Alsophila National Nature Reserve
ZONG Xiuhong1, ZHANG Huayu1, WANG Xin1, LI Zongfeng1, WU Hongying2,LIANG Sheng2, DENG Hongping1*
(1 Key Laboratory of Eco-Environments in Three Gorges Reservoir Region(Ministry of Education),School of Life Sciences, Southwest University, Chongqing 400715, China;2 Chishui Alsophila National Nature Reserve Administration, Chishui, Guizhou 564700,China)
Abstract:Alsophila spinulosa is a mesozoic ancient relic species which under the second-grade state protection. A. spinulosa and its habitat is the protection object of Chishui Alsophila National Nature Reserve of Guizhou. This A. spinulosa population is one of the populations with the most abundant and the largest distribution. Ecological and floristic methods were used to study the species diversity, floristic elements, life form spectrum and community structure. The results show that: (1)a total of 122 species, 96 genera and 60 families were found in the community, among them there were 17 fern species, belonging to 14 genera and 12 families, 2 gymnosperms belonging to 2 genera and 2 families, and 103 angiosperms belonging to 80 genera and 46 families; (2) the families and genera floristic composition was complex with various types, the floristic characteristics were mainly influenced by pantropic, followed by temperate, which reflected its transitional characteristic; (3)the main life form distribution was phanerophytes, about 57.38%, which reflected that the phanerophytes lifeforms were the dominant in mid-subtropical forest; (4)vertical stratification was obvious, trees with the height between 4 m and 12 m had the highest proportion about 76.64%, shrubs with the height between 2 m and 3 m had the highest proportion of 69.78%, and herbs with the height between 0-0.2 m had the highest proportion, the level is obvious; (5)Phyllostachys heterocycla cv. pubescens and A. spinulosa were the dominant tree species; A. spinulosa was the dominant species of shrub layer; Dryopteris erythrosora and Selaginella uncinata were the main herbs. The result of species richness analysis was tree layer < shrub layer < herbaceous layer. Diversity index and uniformity analysis shown aggregated distribution for tree layer and its relatively simple composition. The distribution forms of shrub and herb layers were uniform with low abundance.
Key words:Alsophila spinulosa community; species diversity; flora; life form; community diversity.
