湖南桃源洞国家级自然保护区南方铁杉种群结构与生存分析
2016-08-06丁巧玲刘忠成蔡松辰石祥刚曾茂生廖文波
丁巧玲,刘忠成,王 蕾,蔡松辰,石祥刚*,曾茂生,廖文波
(1 中山大学 生命科学学院,广州 510275; 2 首都师范大学 资源环境与旅游学院,北京100048; 3 湖南桃源洞国家级自然保护区 管理局,湖南炎陵 412500)
湖南桃源洞国家级自然保护区南方铁杉种群结构与生存分析
丁巧玲1,刘忠成2,王蕾2,蔡松辰1,石祥刚1*,曾茂生3,廖文波1
(1 中山大学 生命科学学院,广州 510275; 2 首都师范大学 资源环境与旅游学院,北京100048; 3 湖南桃源洞国家级自然保护区 管理局,湖南炎陵 412500)
摘要:采用样方法对桃源洞国家级自然保护区的牛石坪南方铁杉(Tsuga chinensis)群落和梨树洲南方铁杉群落进行调查,分析群落种类组成和结构特征;采用Shannon-Weiner指数、Simpson指数和Pielou均匀度指数研究群落各层的物种多样性;依据径级频率分布分析2个南方铁杉群落中南方铁杉种群的年龄结构和更新状况。结果表明:(1)桃源洞的2个南方铁杉群落中常绿植物占优势,群落有一定的季相变化,夏季呈暗绿色,秋冬淡黄绿色。(2)牛石坪南方铁杉群落共有维管植物13科21属26种,梨树洲南方铁杉群落共有维管植物22科30属36种;地理成分分析表明温带成分明显高于热带成分,表现为亚热带山地性质。(3)群落垂直结构可分为乔木层(进一步分为乔木上层、乔木中层、乔木下层3个亚层)、灌木层和草本层。(4)2个群落的Shannon-Wiener指数和Simpson指数均为乔木下层>乔木中层>灌木层>乔木上层,Pielou均匀度指数为乔木中层和乔木下层>乔木上层和灌木层。(5)牛石坪南方铁杉群落的优势种群为马银花(Rhododendron ovatum)、南方铁杉、尖连蕊茶(Camellia cuspidata)和鹿角杜鹃(Rhododendron latoucheae),梨树洲南方铁杉群落的优势种群为银木荷(Schima argentea)、南方铁杉、鹿角杜鹃和背绒杜鹃(Rhododendron hypoblematosum)。(6)根据年龄结构和生存分析,牛石坪的南方铁杉种群为衰退型,低龄级个体和种群数量都严重不足,难以实现自然更新;梨树洲的南方铁杉种群为增长型,低龄级个体数较丰富,年龄结构分布基本连续,惟其群落郁闭度较高,自我更新还存在一定的挑战。根据以上结果建议对牛石坪的南方铁杉群落进行就地保护和适当人为干扰,对梨树洲群落加强后续监测工作。
关键词:南方铁杉;群落结构;年龄结构;生存分析;种群更新;湖南桃源洞
南方铁杉(Tsugachinensis)是第三纪孑遗植物,中国特有种。南方铁杉的分布虽广但数量少而分散,是珍稀濒危植物,被列为国家Ⅱ级重点保护植物,具有很高的科研价值和潜在的经济价值[1]。南方铁杉分布于浙江、安徽南部、福建北部、武夷山、江西武功山、湖南莽山、广东北部、广西北部及云南麻栗坡等地,分布区地跨中亚热带至北亚热带[1]。其分布的垂直高度变化较大,在海拔600~2 100 m之间,但以海拔800~1 400 m的生长较好[1]。目前,关于南方铁杉的研究已有许多报道,包括群落结构和物种多样性[1-6]、空间分布格局和种群动态[4,7-17]、种间和种内关系[18-21]、生态位[5,22]、生长规律[23-25]、越冬策略[26]、土壤金属含量与土壤养分关系[27]、菌根生态学[28-29]以及扦插繁殖、林隙干扰规律等[30-31]方面。以上对南方铁杉群落的研究包括浙江九龙山、福建光泽、福建武夷山、江西武夷山、湖南莽山、湖南黄桑坪、广西猫儿山、贵州茂兰和贵州高坡等区域。但至目前为止,还未有对湖南桃源洞南方铁杉群落的研究报道。本研究从南方铁杉群落的外貌、组成、结构和种群年龄结构等方面着手,旨在探讨桃源洞南方铁杉群落的结构和种群的更新,为加强对南方铁杉的保护和管理提供理论依据。
1研究地概况
桃源洞国家级自然保护区位于湖南省东南部炎陵县的东北部(114°03′45″~114°07′30″ E, 26°30′00″~26°32′30″ N),总面积113 153 hm2[32]。该区山峦重叠、地势险峻、沟谷深邃、溪流纵横,属中亚热带季风湿润气候,年平均气温12.3~14.4 ℃,1月份气温最低( 0.2~1.5 ℃),7月份气温最高(25.8~29.7 ℃),极端最高气温34.5 ℃(1990年8月16日),极端最低气温 9 ℃(1991年12月29日);年降水量1 967.9~2 165.2 mm,最大降水强度为129.6 mm·d-1(1984年9月1日),4~6月降水量占全年的42 %;土壤分属4个亚类:山地草甸土(分布海拔1 700~1 841 m)、山地黄棕壤(分布海拔1 200~1 700 m)、山地暗黄壤(分布海拔650~1 200 m)和山地黄红壤(分布海拔0~650 m)[33]。
2研究方法
2.1样地设置
针对湖南桃源洞国家级自然保护区的植被状态进行实地考察,发现在牛石坪(114°2′49.2″ E, 26°25′53.6″ N)和梨树洲(113°59′05.54″ E, 26°20′56.38″ N)分别保存有较丰富的南方铁杉种群,为探讨其种群生存状态,特设置两片样地进行调查,样地面积分别为1 000 m2和1 600 m2。其中,牛石坪样地海拔1 370 m,坡向东南,坡度40°,土壤为黄棕壤,郁闭度为0.65~0.70;梨树洲样地海拔1 495 m,坡向向东,坡度45°,土壤为黄棕壤,郁闭度为0.95。南方铁杉种群在这两片样地中生长状况差异较大,牛石坪样地南方铁杉数量较少但多为大树,鲜有幼树;梨树洲样地南方铁杉数量较多但多为小树,少大树。此外,梨树洲南方铁杉群落的郁闭度明显高于牛石坪南方铁杉群落的郁闭度。因此,对这2个南方铁杉群落进行比较研究,将有利于了解南方铁杉种群的生存状态及其更新演替的趋势,可为南方铁杉的保护和管理提供理论依据。本研究采用相邻格子法将2个样地进一步划分成面积为10 m×10 m的样方,每个样方内再设置1个2 m×2 m的小样方。样方调查记录植物的种名、胸径、高度、冠幅,起测径阶>1.5 cm;小样方记录植物的种名、株数、高度和盖度。
2.2数据分析
2.2.1重要值的计算根据王伯荪等[34]的测度方法,调查计算乔、灌木的重要值:
乔木重要值IV=(相对多度+相对频度+相对优势度)/3×100
灌木重要值IV=(相对多度+相对频度+相对盖度)/3×100
2.2.2多样性测度方法采用马克平等[35]的测度方法确定南方铁杉群落的物种多样性:
Shannon-Weiner指数(H′):H′=∑PilnPi
Simpson指数(D):D=1-∑Pi2
Pielou均匀度指数(JSW):JSW=(-∑PilnPi)/lnS式中,Pi=Ni/N,N为调查样地中乔木层、灌木层或草本层的总个体数;Ni为第i个物种的个体数;S为调查样地中乔木层、灌木层或草本层的物种总数。
2.2.3高度结构和径级结构的划分根据南方铁杉种群的高度组成,按每隔2 m划分成15个等级,第1高度级为0~2 m,第15高度级为28~30 m,也是本群落的最大高度级。种群年龄结构采用立木径级代替,共划分13个径级,每5 cm为一个径级,第1径级为0~5 cm,第2径级为6~10 cm,以此类推,最后把大于60 cm的归为第13径级[2]。在绘制种群年龄结构的条形图时,第1径级对应第1龄级,第2径级对应第2龄级,以此类推。
2.2.4静态生命表编制静态生命表的编制按照江洪[36]的方法:x是单位时间年龄等级的中值;ax是在x龄级内现有的个体数;lx是在x龄级开始时标准化存活个体数(转化为1 000);dx是从x到x+1龄级间隔期间标准化死亡数;qx是从x到x+1龄级间隔期间死亡率;Tx是从x龄级到超过x龄级的个体总数;ex是进入x龄级的生命期望寿命;Kx是消失率(损失度);Sx是存活率。相关参数的关系和计算公式如下:
lx=ax/a0× 1 000
dx=lx-lx+1
qx=dx/lx
Lx=(lx+lx+1)/2
ex=Tx/lx
Kx=lnlx-lnlx+1
Sx=lx+1/lx
由于牛石坪南方铁杉种群属于“幼龄株数少的衰退种群”,因此无法编制生命表。本研究仅对梨树洲南方铁杉种群编制了生命表和进行了生存分析。此外,在生命表的编制中,为了避免出现负值,往往对各龄级的个体数进行标准化后作匀滑处理[36]。但是Wretten等[37]认为,生命表分析中产生一些负值虽然与数据假设技术不符,但仍提供有用的生态记录,即表明种群并非静止不动,而是在发展或衰落之中。因此,本研究未对相关数据作匀滑处理,而是将各龄级个体数标准化后直接用于各参数的计算。
2.2.5生存分析方法种群的生存分析按照杨凤祥[38]的方法进行,计算种群生存率函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti),并绘制生存曲线、累计死亡率曲线、死亡密度曲线和危险率曲线。这4个函数的计算公式如下:
S(i)=S1·S2·S3···Si(Si为存活率)
F(i)=1-S(i)
f(ti)=(Si-1-Si)/hi(hi为龄级宽度)
λ(ti)=2(1-Si)/[hi(1+Si)]
3结果与分析
3.1群落种类组成及地理成分分析
牛石坪南方铁杉群落共有维管植物26种,隶属于13科21属。其中,裸子植物仅南方铁杉1种,被子植物12科20属25种。种子植物中,物种数最多的科为山茶科,共5属5种;山矾科次之,为1属4种;樟科和杜鹃花科分别有3属3种和2属3种;五加科和漆树科分别为2属2种和1属2种;其余的科均为1属1种(表 1)。梨树洲南方铁杉群落共有维管植物36种,隶属于22科30属。其中,蕨类植物3科3属4种,裸子植物3科4属4种,被子植物16科23属28种。种子植物中,物种数最多的科为山茶科和杜鹃花科,分别为4属5种和2属5种;蔷薇科次之,为2属3种;樟科、壳斗科和松科皆为2属2种;其余的科均为1属1种(表 1)。
牛石坪南方铁杉群落中种子植物共21属,其中热带分布区类型10属,占47.62%,温带分布区类型11属,占52.38%;梨树洲南方铁杉群落中种子植物共27属,其中热带分布区类型11属,占44.00%,温带分布区类型14属,占56.00%。2个南方铁杉群落的温带成分均高于热带成分,与其位于罗霄山脉西坡,受到西部和北部温带成分的影响相符合。将桃源洞2个南方铁杉群落的地理成分组成与福建光泽、浙江九龙山、贵阳高坡的南方铁杉群落进行比较,发现四者的温带成分分布在52.38%~65.79%,并且均明显高于热带成分。湖南桃源洞(海拔1 370~1 495 m)、贵阳高坡(海拔1 350~1 450 m)、福建光泽(海拔1 445~1 625 m)、浙江九龙山(海拔1 300~1 620 m)4个区域均属于中国亚热带常绿阔叶林区,其中湖南桃源洞、贵阳高坡位于常绿林区的南部亚地带,福建光泽、浙江九龙山位于常绿区的北部亚地带,按照地带性规律,南部亚地带的温带成分低于北部亚地带[39]。但从数据看,贵阳高坡南方铁杉群落的温带成分却与福建光泽、浙江九龙山的基本相当,甚至略高(表2),这可能与其特殊的喀斯特地貌相关[4]。4个南方铁杉群落的温带成分中,北温带分布、东亚北美间断分布所占的比例较大(表 2)。
3.2群落外貌与垂直结构
在群落外貌上,牛石坪南方铁杉群落和梨树洲南方铁杉群落都为常绿落叶针阔叶混交林,但以常绿树种占优势,亦有部分落叶树种如漆树(Toxicodendronvernicifluum)、檫木(Sassafrastzumu)、青榨槭(Acerdavidii)、柿树(Diospyroskaki)、木蜡树(Toxicodendronsylvestre)、吴茱萸五加(Gambleaciliatavar.evodiifolia)等,因此群落有一定的季相变化,夏季呈暗绿色,秋冬淡黄绿色。
根据群落的高度级频率分布(图 1,a、b),除了草本层外,桃源洞两处的南方铁杉群落的林木层可分为4层,由下至上依次为灌木层、乔木下层、乔木中层和乔木上层。但由于2个群落的组成及所处的演替阶段不同,两者的分层高度稍为差异:牛石坪群落的灌木层为0~4 m,乔木下层为4~10 m,乔木中层为10~16 m,乔木上层为16~30 m(图1,a);梨树洲群落的0~4 m为灌木层,4~6 m为乔木下层,6~14 m为乔木中层,14~22 m为乔木上层(图1,b)。
从乔木层和灌木层的生活型来看,桃源洞南方铁杉群落共有常绿针叶树3种,常绿阔叶树29种,落叶阔叶树10种。牛石坪南方铁杉群落各层的常绿树种相对多度由上自下依次为89.74%、59.46%、84.16%和95.03%;杜鹃花科植物在灌木层、乔木下层和乔木中层中都为多度最高的类群,主要为马银花(Rhododendronovatum);山茶科在灌木层和乔木下层也有较高的多度,主要为尖叶连蕊茶(Camelliacuspidata);而南方铁杉在乔木上层占据绝对的多度优势(图1,c)。梨树洲群落各层常绿树的相对多度由上自下依次为97.30%、82.29%、69.63%和89.10%;杜鹃花科植物在灌木层和乔木下层为多度最高的类群,主要是鹿角杜鹃(Rhododendronlatoucheae)和背绒杜鹃(Rhododendronhypoblematosum);乔木中层和乔木上层中多度最高的类群为山茶科植物,主要是银木荷(Schimaargentea);南方铁杉在各层都占有一定的比例,多度仅次于杜鹃花科或山茶科植物(图1,d)。
注:NP表示物种数,NG表示属数。
Note: NP. number of species, NG. number of genera.

表2 5处南方铁杉群落种类组成的地理成分比较

a、c. 牛石坪南方铁杉群落;b、d. 梨树洲南方铁杉群落。图1 桃源洞南方铁杉群落高度频率分布(a、b)及其垂直结构图(c、d)a, c. Tsuga chinensis community from Niushiping; b, d. Tsuga chinensis community from LishuzhouFig. 1 The height class frequency distribution (a, b) and vertical structure (c, d) of T. chinensis community from Taoyuandong
3.3群落物种多样性
由表3可知,牛石坪南方铁杉群落乔木层和草本层的Shannon-Wiener指数、Simpson指数和Pielou均匀度指数均大于灌木层,草本层的Simpson指数和Pielou均匀度指数略大于乔木层,而Shannon-Wiener指数则略低于乔木层;梨树洲南方铁杉群落的Shannon-Wiener指数、Simpson指数和Pielou均匀度指数变化趋势相同,均为草本层>乔木层>灌木层;分别比较2个南方铁杉群落各层的3个指数,其差别不大。具体比较每个群落灌木层、乔木下层、乔木中层和乔木上层的三个指数可得:2个南方铁杉群落的Shannon-Wiener指数和Simpson指数均为乔木下层>乔木中层>灌木层>乔木上层;牛石坪南方铁杉群落的Pielou均匀度指数为乔木中层>乔木下层>灌木层>乔木上层;梨树洲南方铁杉群落的Pielou均匀度指数为乔木中层>乔木下层>乔木上层>灌木层,并且乔木层的3个亚层均匀度指数变化不大,这与银木荷和南方铁杉在该群落乔木上层分布均匀且数量较多相关(表 3)。
3.4群落乔、灌木种群重要值
对2个南方铁杉群落的乔、灌木种群重要值分别进行分析,结果表明:(1)牛石坪南方铁杉群落乔木层中的优势种群是马银花种群和南方铁杉种群,重要值分别为24.84和16.31;次优势种群是尖叶连蕊茶种群、鹿角杜鹃种群和漆树(Toxicodendronvernicifluum)种群,重要值都在5以上(表 4)。(2)牛石坪南方铁杉群落灌木层中的优势种群是马银花种群,重要值为52.14,高于其余所有种群的重要值之和;次优势种群是尖连蕊茶种群和鹿角杜鹃种群,重要值分别为18.08和6.98;南方铁杉在灌木层没有分布(表 5)。(3)梨树洲南方铁杉群落乔木层中的优势种群是银木荷种群和南方铁杉种群,重要值分别为23.98和20.85;次优势种群为鹿角杜鹃种群、吴茱萸五加种群和华东山柳(Clethrabarbinervis)种群(表 6)。(4)梨树洲南方铁杉群落灌木层中的优势种群是鹿角杜鹃种群和背绒杜鹃种群;次优势种群是华东山柳种群、吴茱萸五加种群、马银花种群和南方铁杉种群,其中南方铁杉种群的重要值为3.57(表 7)。
3.5南方铁杉种群的年龄结构和数量动态
从图2可以看出,牛石坪南方铁杉群落中南方铁杉种群的年龄结构为衰退型种群,幼年阶段的个体数量较少,成年个体相对丰富;种群内个体集中分布在第6至第9龄级,并在第7龄级出现个体数量高峰;此外,种群在第2龄级、第12龄级和13龄级出现断层,表明受到过严重的干扰,例如人为砍伐、自然灾害等。而梨树洲南方铁杉群落中南方铁杉种群的年龄结构为增长型种群,个体数随龄级的增加而递减;第10到第13龄级个体数为0;第6龄级和第7龄级出现断层,同样表明存在干扰。
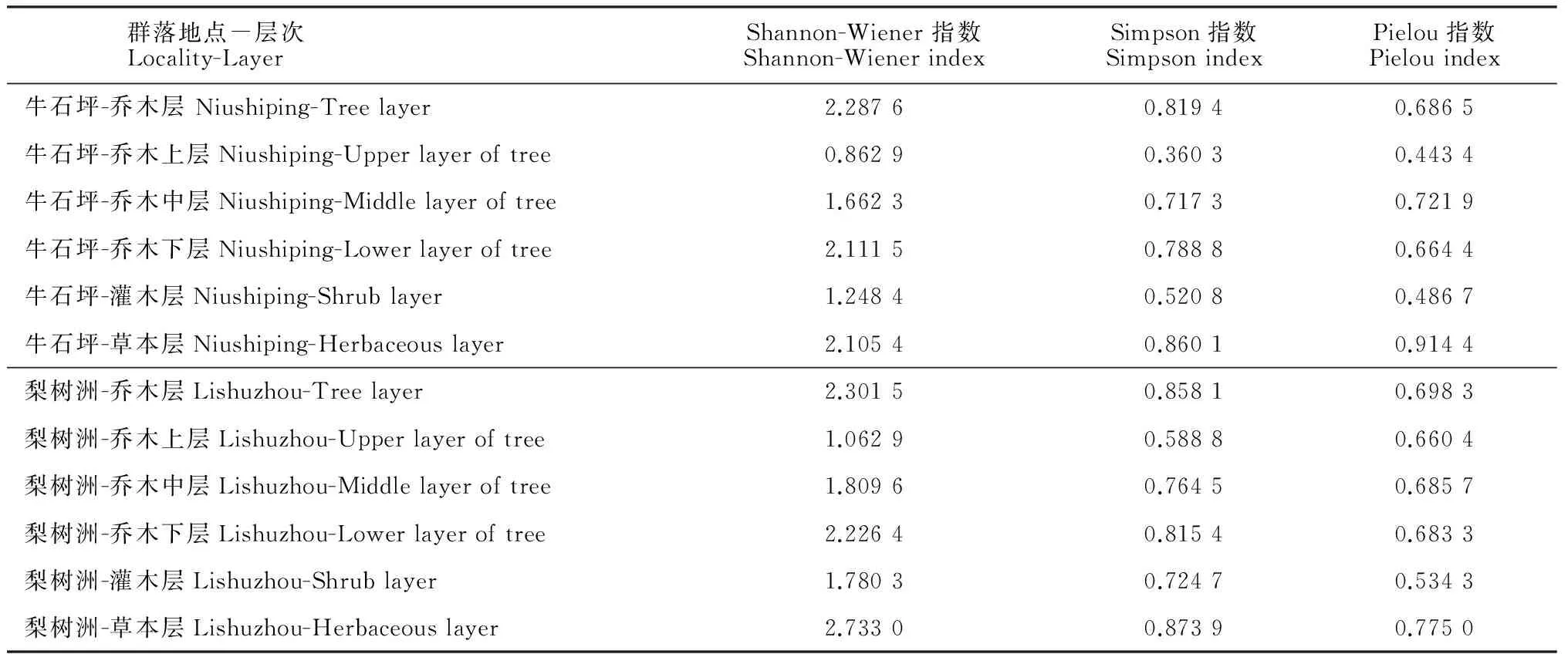
表3 桃源洞南方铁杉群落物种多样性
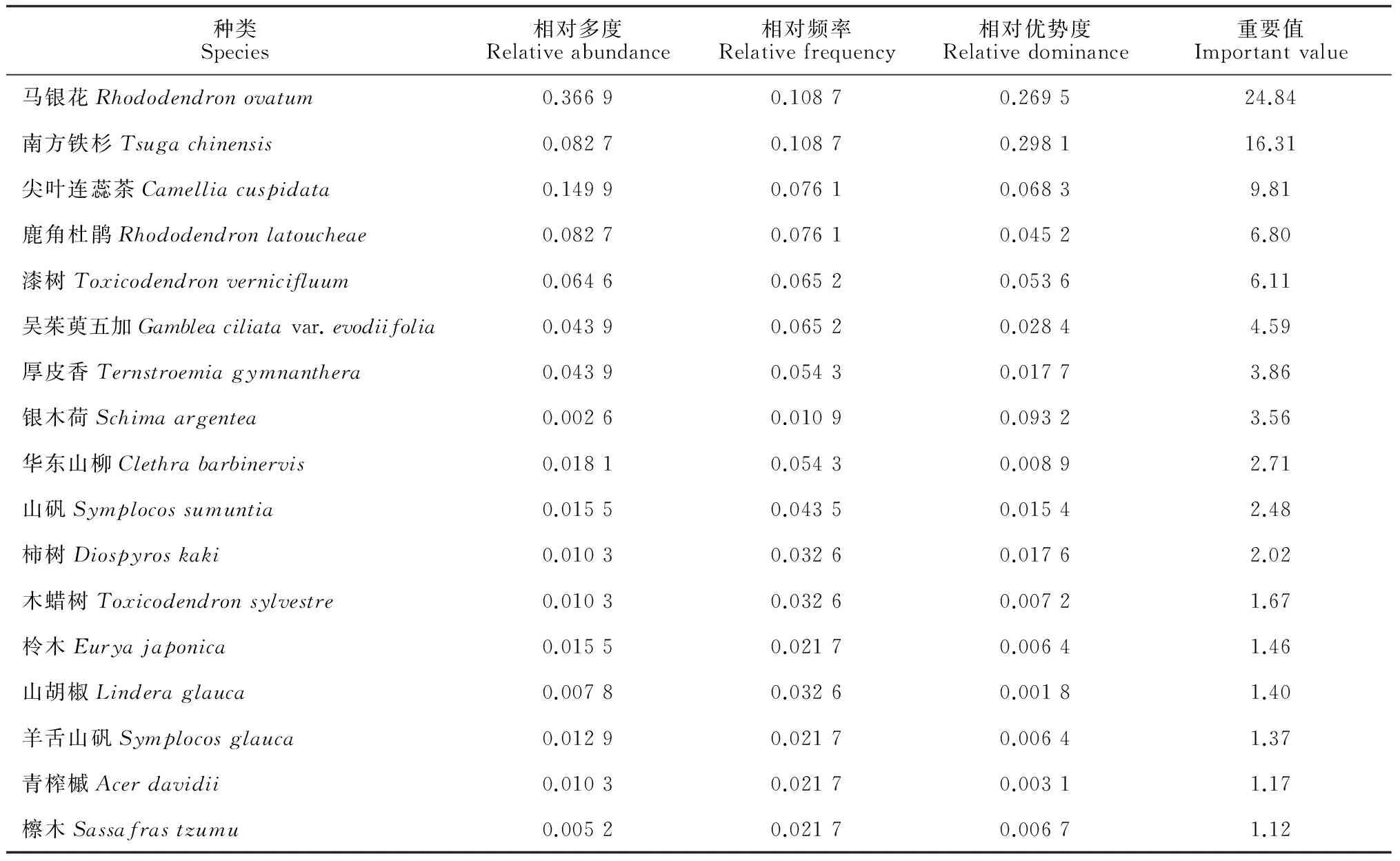
表4 牛石坪南方铁杉群落乔木种群重要值
注:仅列出了重要值大于1的种群。表6、7同。
Note: Tree populations which importance values less than 1.00 are not listed. The same as Table 6 and 7.

表5 牛石坪南方铁杉群落灌木种群重要值
注:包括所有的灌木种群。
Note: Including all the shrub populations.
梨树洲南方铁杉种群的静态生命表(表 8)和存活曲线(图3)表明:(1)梨树洲南方铁杉种群结构存在一定的波动性;(2)第3龄级是其存活的一个关键时期,表现为其存活数量迅速下降(图3,a)以及死亡率和消失率达到第一个峰值(图3,b);(3)在第3龄级以前,该种群的生存率、累积死亡率和危险率变化显著,生存率锐减而累积死亡率和危险率骤增(图3,c、d);(4)到第5龄级以后,生存率和死亡率变化趋于平缓,但由于干扰的存在,使得第5到第6龄级的存活率为0(图3,c)。
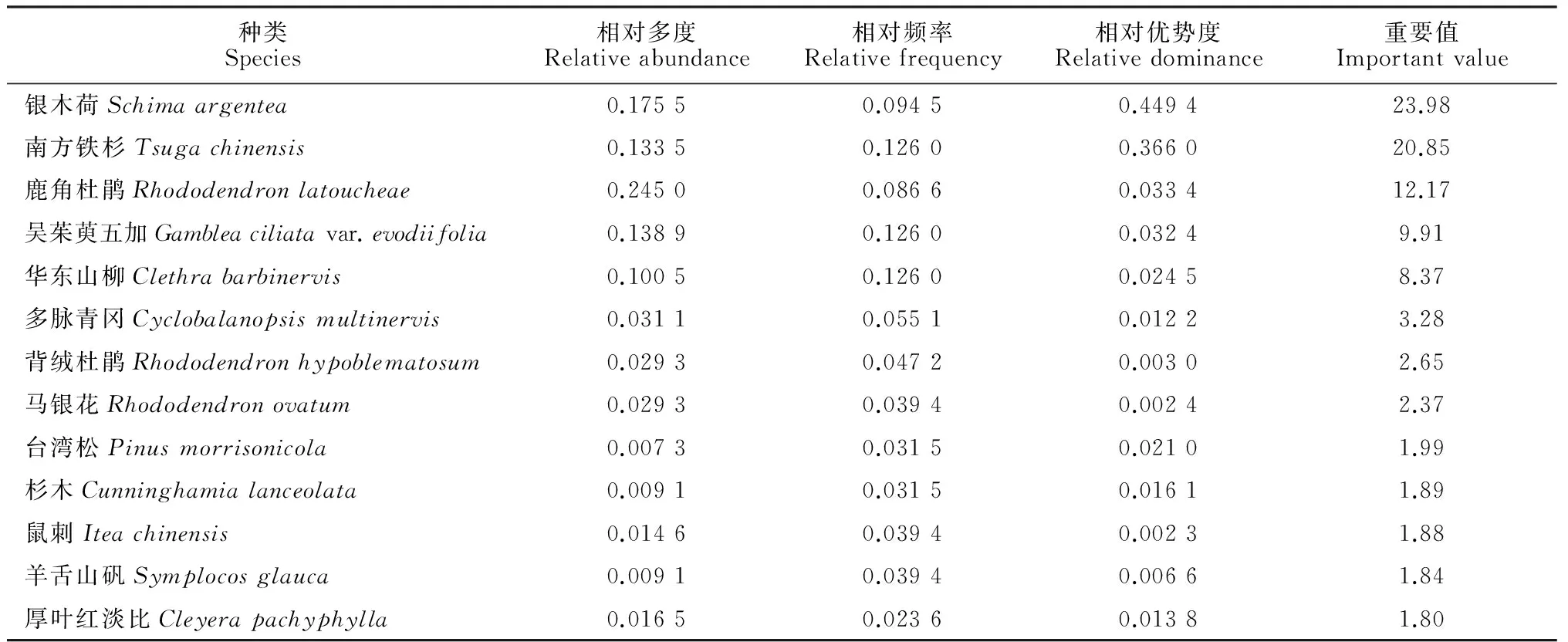
表6 梨树洲南方铁杉群落乔木种群重要值
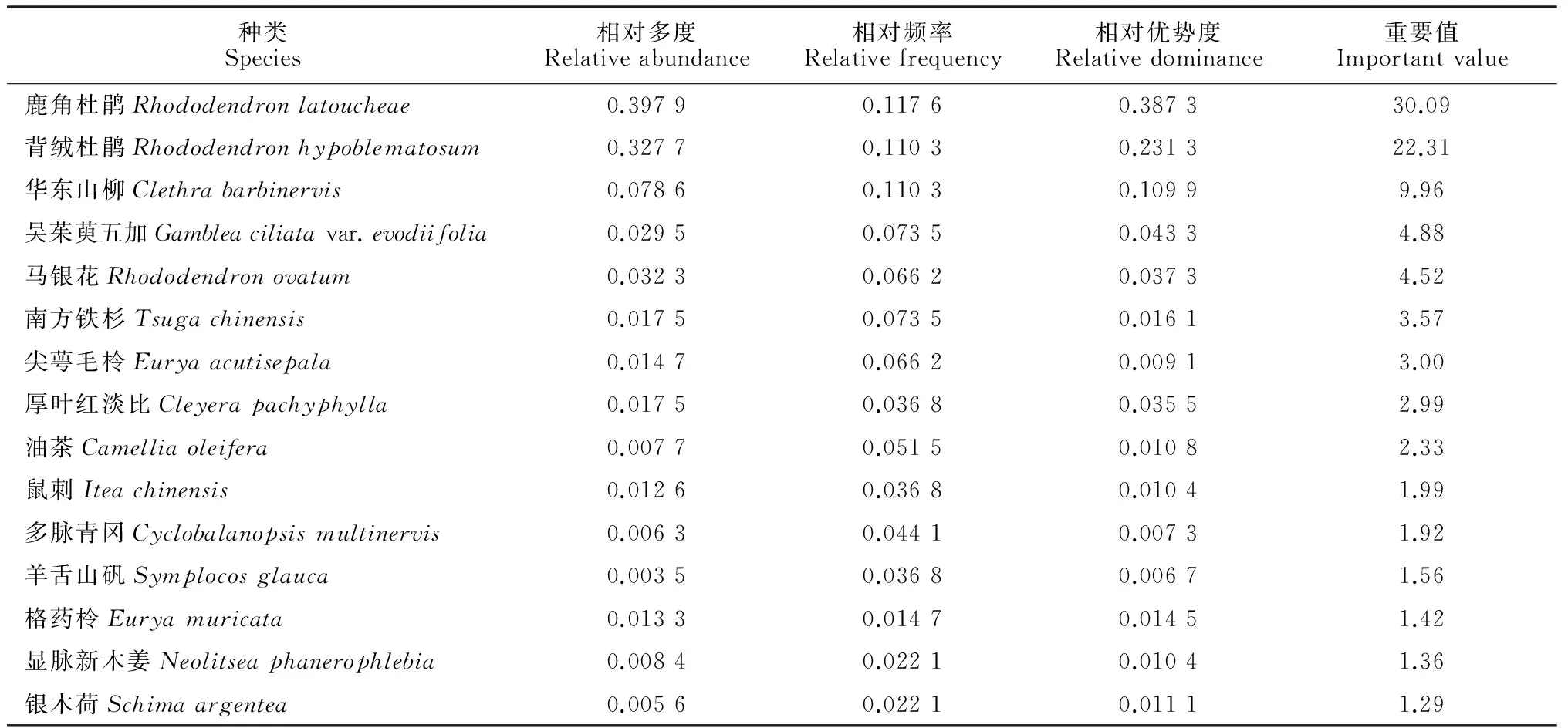
表7 梨树洲南方铁杉群落灌木种群重要值
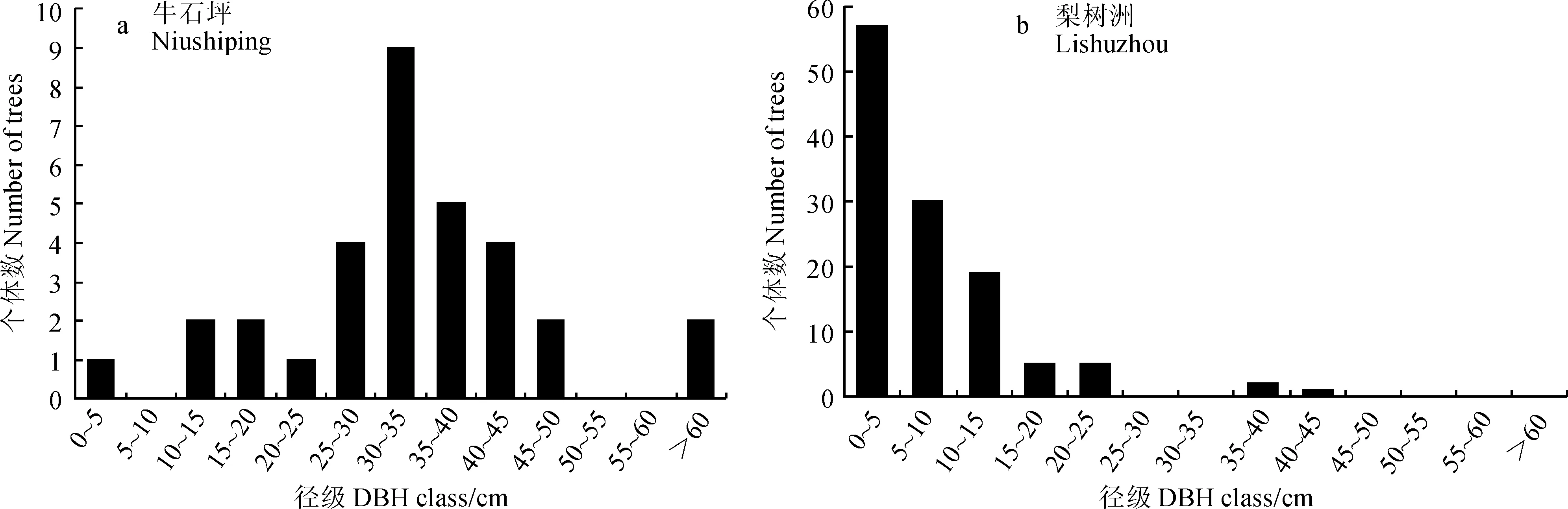
图2 桃源洞2个南方铁杉种群年龄结构Fig. 2 The age class frequency distribution of T. chinensis population from Taoyuandong

龄级Ageclass径级DBHclass组中值MeanvalueaxlxlnlxdxqxLxTxexKxSx10~52.55710006.9084740.47476315881.5880.6420.52625~107.5305266.2661930.3674308251.5670.4570.633310~1512.5193335.8092460.7372113951.1841.3350.263415~2017.55884.47400.000881842.1000.0001.000520~2522.55884.474881.00044961.100/0.000625~3027.500/0/053///730~3532.500/-35/1853///835~4037.52353.558180.50026351.0000.6930.500940~4542.51182.865181.000990.500/0.0001045~5047.500/0/00///1150~5552.500/0/00///1255~6057.500/0/00///13>6070.000/0/00///
注:DBH.胸径;ax.存活数;lx.存活量;dx.死亡率;qx.死亡率;Lx.区间寿命;Tx.总寿命;ex.期望寿命;Kx.消失率;Sx.存活率。
Note:DBH. Diameter at breast height;ax. Survival;lx. Survival quantity;dx. Death number;qx. Mortality rate;Lx. Span life;Tx. Total life;ex. Life expectancy;Kx. Vanish rate;Sx. Survival rate.

a. 存活曲线;b. 死亡率(qx)和消失率(Kx)曲线;c. 生存率[S(i)]和累积死亡率[F(i)]曲线;d. 死亡密度[f(ti)]和危险率[λ(ti)]曲线图3 梨树洲南方铁杉种群生存分析曲线a. Survival curve; b. Mortality rate (qx) and vanish rate (Kx) value curves; c. Survival rate [S(i)] and cumulative mortality rate [F(i)] functional curves; d. Mortality density [f(ti)] and hazard rate [λ(ti)]functional curvesFig. 3 Survival analysis curves of T. chinensis population from Lishuzhou
总的来说,牛石坪南方铁杉种群的个体数明显少于梨树洲南方铁杉种群的个体数,前者仅有南方铁杉32株,后者共有南方铁杉98株。根据南方铁杉木解析资料[25],第7龄级植株树龄在100年左右,表明100年前牛石坪南方铁杉种群存在自我更新。然而,由于低龄级个体的缺乏和种群总体数量的不足,牛石坪南方铁杉种群可因为高龄级个体的生理衰老而不断死亡和低龄级个体的缺失而呈现更新困难和衰亡的趋势。梨树洲南方铁杉种群中低龄级个体数较丰富,年龄结构分布基本连续,理论上可实现自我更新。但是,根据生存分析,第3龄级是其存活的一个关键时期,群落的郁闭度为其限制因子,并且梨树洲南方铁杉种群还较年轻,其是否能自然更新还存在一定的挑战。此外,两个南方铁杉种群均有较严重的干扰现象,成为影响高龄级个体数量的一个重要原因。
4讨论
桃源洞自然保护区中牛石坪南方铁杉群落共有维管植物13科21属26种,梨树洲南方铁杉群落共有维管植物22科30属36种。地理成分分析表明温带成分明显高于热带成分,表现为温带向热带过渡的特性,也体现了该群落的亚热带山地性质。2个群落中常绿树种占优势,整体外貌有一定的季相变,春夏暗绿色,秋冬淡黄绿色。群落垂直结构可分为乔木层、灌木层和草本层,其中乔木层又可分为3亚层。2个南方铁杉群落的Shannon-Wiener指数和Simpson指数均为乔木下层>乔木中层>灌木层>乔木上层,Pielou均匀度指数为乔木中层和乔木下层>乔木上层和灌木层。
牛石坪南方铁杉群落乔木层的优势种群为马银花种群和南方铁杉种群,次优势种群是尖连蕊茶种群、鹿角杜鹃种群和漆树种群;灌木层优势种群是马银花种群,次优势种群是尖连蕊茶种群和鹿角杜鹃种群。其中,南方铁杉种群几乎全部的个体都分布于乔木上层,是乔木上层的压倒性优势植物。根据牛石坪南方铁杉种群的年龄结构分布特征,表明其属于衰退型种群。该南方铁杉种群不论是低龄级个体还是种群的总体数量都严重不足,难以实现自然更新。
梨树洲南方铁杉群落乔木层中的优势种群是银木荷种群和南方铁杉种群,次优势种群为鹿角杜鹃种群、吴茱萸五加种群和华东山柳种群;灌木层优势种群是鹿角杜鹃种群和背绒杜鹃种群,次优势种群是华东山柳种群、吴茱萸五加种群、马银花种群和南方铁杉种群。南方铁杉在各层均有分布,乔木中层有42株、灌木层有25株、乔木下层有17株、乔木上层有14株。梨树洲南方铁杉种群的年龄结构分布属于增长型,低龄级个体数较丰富,年龄结构分布基本连续,理论上可实现自我更新。
南方铁杉作为国家Ⅱ级重点保护植物,加强对其的保护具有很高的学术价值和实际意义。南方铁杉濒危的原因主要有三点:第一是南方铁杉的种子休眠期较长且幼苗呈聚集生长,导致种群更新较慢和幼苗死亡率较高;二是南方铁杉为喜光树种,在其生长的各个阶段都需要充足的阳光,因此群落的郁闭度是其主要的限制因子之一;三是南方铁杉多散生于针阔混交林中,为小种群,自然灾害和人为破坏带来的伤害可能是毁灭性的[1]。在本研究的2个南方铁杉群落中,牛石坪南方铁杉种群虽然是乔木层优势种群,但已经处于衰退阶段,随着南方铁杉成年个体的不断死亡以及幼苗的缺乏,呈现更新困难和衰亡的趋势。而梨树洲南方铁杉种群目前生长旺盛,具有一定的更新率,但该群落郁闭度高,且有鹿角杜鹃等耐阴种群的竞争,自我更新存在一定的挑战。总体来说,南方铁杉种群早期的自我更新限制因子主要是郁闭度,后期个体数量减少的主要原因是人为砍伐等干扰。因此,建议对桃源洞南方铁杉群落加强后续监测,如有必要应进行人为干扰预以降低林地郁闭度和加强群落通风条件。此外,还应强保护性标识牌的使用和警示。
致谢:中山大学2012级生物学类本科生黄玉梅、杜静静、刘子豪,研究生许可旺、赵万义,首都师范大学刘羽霞、阿尔孜古力,桃源洞自然保护区孟长庆等参加了野外调查,特此鸣谢!
参考文献:
[1]张志祥. 珍稀濒危植物南方铁杉研究进展[J]. 生物学教学, 2011, 36(6): 3-5.
ZHANG Z X. Research advance ofTsugatchekiangensis[J].BiologicalTeaching, 2011, 36(6): 3-5.
[2]张志祥, 刘鹏, 刘春生, 等. 浙江九龙山南方铁杉(Tsugatchekiangensis)群落结构及优势种群更新类型[J]. 生态学报, 2008, 28(9): 4 547-4 558.
ZHANG Z X, LIU P, LIU C S,etal. The structure characteristics and dominant population regeneration types ofTsugatchekiangensiscommunities in the Jiulongshan National Natural Reserve of Zhejiang Province[J].ActaEcologySinica, 2008, 28(9): 4 547-4 558.
[3]陈璟. 莽山自然保护区南方铁杉种群物种多样性和稳定性研究[J]. 中国农学通报, 2010, 26(12): 81-85.
CHEN J. The Species Diversity and Stability ofTsugatchekiangensisCommunity in Mang Mountain Nature Reserve[J].ChineseAgriculturalScienceBulletin, 2010, 26(12): 81-85.
[4]冯祥麟, 胡刚, 刘正华. 贵阳高坡南方铁杉群落特征及种群动态调查研究[J]. 贵州林业科技, 2011, 39(2): 26-29.
FENG X L, HU G, LIU Z H. Research on the Community Characteristics and Population Dynamics ofTsugachinesisvar.tchekiangensison Gaopo in Guiyang City[J].GuizhouForestryScienceandTechnology, 2011, 39(2): 26-29.
[5]谢琼中. 南方铁杉群落物种多样性及乔木优势种生态位初步研究[J]. 天津农业科学, 2011, 17(2): 133-136.
XIE Q Z. Primary Study on Species Diversity and Niche Characteristics of DominantTreeSpeciesin Tsuga tchekiangensis Community[J].TianjinAgriculturalSciences, 2011, 17(2): 133-136.
[6]谢旺生. 福建光泽南方铁杉群落植物组成与多样性分析[J]. 福建林业科技, 2012, 39(3): 8-14.
XIE W S. Plant Species Composition and Diversity ofTsugachinensisvar.tchekiangensis Community in Guangze, Fujian, China[J].JourofFujianForestryScienceandTechnology, 2012, 39(3): 8-14.
[7]杜道林, 刘玉成, 刘川华. 茂兰喀斯特山地南方铁杉种群结构和动态初探[J]. 西南师范大学学报 (自然科学版) , 1994, 19(2): 169-174.
DU D L, LIU Y C, LIU C H. A priliminary study on the structures and dynamics ofTsugatechekiangensispopulations in Karst Mountain of Maolan[J].JournalofSouthwestChinaNormalUniversity(Natural Science) Natual Science Edition, 1994, 19(2): 169-174.
[8]黄宪刚, 谢强. 猫儿山南方铁杉种群结构和动态的初步研究[J]. 广西师范大学学报 (自然科学版), 2000, 18(2): 86-90.
HUANG X G, XIE Q. A priliminary study on the structures and dynamics ofTsugatechekiangensis population on Maorshan mountain[J].JournalofGuangxiNormalUniversityNatual Science Edition, 2000, 18(2): 86-90.
[9]封磊, 洪伟, 吴承祯, 等. 珍稀濒危植物南方铁杉种群动态研究[J]. 武汉植物学研究, 2003, 21(5): 401-405.
FENG L, HONG W, WU C Z,etal. Study on the dynamics of the endangered plant population ofTsugatchekiangensis[J].JournalofWuhanBotanicalResearch, 2003, 21(5): 401-405.
[10]封磊, 洪伟, 吴承祯, 等. 南方铁杉种群结构动态与空间分布格局[J]. 福建林学院学报, 2008, 28(2): 110-114.
FENG L, HONG W, WU C Z,etal. Structure dynamics and spatial distribution pattern of population ofTsugatchekiangensis[J].JournalofFujianCollegeofForestry, 2008, 28(2): 110-114.
[11]郭连金, 洪森荣, 夏华炎. 武夷山自然保护区濒危植物南方铁杉种群数量动态分析[J]. 上饶师范学院学报, 2006, 26(6): 74-78.
GUO L J, HONG S R, XIA H Y. Analysis on the Number Dynamic ofTsugaTchekiangensisPopulation in Wuyishan Nature Reserve[J].JournalofShangraoNormalCollege, 2006, 26(6): 74-78.
[12]张志祥, 刘鹏, 蔡妙珍, 等. 九龙山珍稀濒危植物南方铁杉种群数量动态[J]. 植物生态学报, 2008, 32(5): 1 146-1 156.
ZHANG Z X, LIU P, CAI M Z,etal. Population quantitative characteristics and dynamics of rare and endangeredtsugatchekiangensisin Jiulongshan Nature Reserve of China[J].JournalofPlantEcology, 2008, 32(5): 1146-1156.
[13]张志祥, 刘鹏, 等. 珍稀濒危植物南方铁杉种群结构与空间分布格局研究[J]. 浙江林业科技, 2009, 29(1): 7-14.
ZHANG Z X, LIU P,etal. Studies on the population structure and spatial distribution pattern ofTsugachinensisvar.tchekiangensis[J].Jour.ofZhejiangForestryScienceandTechnology, 2009, 29(1): 7-14.
[14]何建源, 卞羽, 吴焰玉, 等. 不同坡向濒危植物南方铁杉的分布格局[J]. 中国农学通报, 2010, 26(13): 122-125.
HE J Y, BIAN Y, WU Y Y,etal. Spatial distribution pattern of the endangered plantTsugachekiangensisin Different Aspect[J].ChineseAgriculturalScienceBulletin, 2010, 26(13): 122-125.
[15]王大来. 莽山南方铁杉种群格局分布格局研究[J]. 中国农学通报, 2010, 26(1): 74-77.
WANG D L. Distribution pattern ofTsugatchekiangensispopulations in Mang Mountain[J].ChinsesAgriculturalScienceBulletin, 2010, 26(1): 74-77.
[16]李林, 魏识广, 黄忠良, 等. 猫儿山两种孑遗植物的更新状况和空间分布格局分析[J]. 植物生态学报, 2012, 36(2): 144-150.
LI L, WEI S G, HUANG Z L,etal. Regenerative condition and analysis of spatial distribution pattern of two relic plants in Mao’ershan Mountain, China[J].ChineseJournalofPlantEcology, 2012, 36(2): 144-150.
[17]杨清培, 金志农, 裘利洪, 等. 江西武夷山南方铁杉更新格局及代际关联性分析[J]. 生态学杂志, 2014, 33(4): 939-945.
YANG Q P, JIN Z N, QIU L H,etal. Regeneration pattern and intergeneration association ofTsugachinensisvar.tchekiangensisin Jiangxi Wuyishan National Nature Reserve[J].ChineseJournalofEcology, 2014, 33(4): 939-945.
[18]刘春生, 刘鹏, 张志祥, 等. 九龙山南方铁杉群落物种多样性及乔木种种间联结性[J]. 生态环境, 2008, 17(4): 1 533-1 540.
LIU C S, LIU P, ZHANG Z X,etal. Species diversity and interspecific association of trees ofTsugatchekiangensiscommunities in Jiulong mountain[J].EcologyandEnvironment, 2008, 17(4): 1 533-1 540.
[19]何建源, 荣海, 吴焰玉, 等. 武夷山南方铁杉群落乔木层种间联结研究[J]. 福建林学院学报, 2010, 30(2): 169-173.
HE J Y, RONG H, WU Y Y,etal. Study on interspecific association of main populations in arborous layer of aTsugatchekiangensiscommunity in Wuyishan National Nature Reserve of Fujian Province[J].JournalofFujianCollegeofForestry, 2010, 30(2): 169-173.
[20]罗金旺. 福建光泽天然林中南方铁杉的种内与种间竞争[J]. 林业科技开发, 2011, 25(4): 71-74.
LUO J W. Intra and interspecific competition inTsugachinensisvar.tchekiangensisin a natural forest in Guangze, Fujian, China[J].ChinaForestryScience&Technology, 2011, 25(4): 71-74.
[21]赵峰. 莽山南方铁杉群落种间关系研究[J]. 中国农学通报, 2011, 27(31): 68-72.
ZHAO F. The relationship of species inTsugatchekiangensiscommunity in Mangshan Mountain Natural Reserve[J].ChineseAgriculturalScienceBulletin, 2011, 27(31): 68-72.
[22]刘春生, 刘鹏, 张志祥, 等. 九龙山濒危植物南方铁杉的生态位研究[J]. 武汉植物学研究, 2009, 27(1): 55-61.
LIU C S, LIU P, ZHANG Z X,etal. The niche characteristics of endangered plantTsugatchekiangensisin Jiulongshan National Nature Reserve of Zhejiang Province[J].JournalofWuhanBotanicalResearch, 2009, 27(1): 55-61.
[23]李晓铁. 猫儿山林区南方铁杉生长调查初报[J]. 广西林业科技, 1992, 21(1): 24-26.
LI X T. Primary research ofTsugatchekiangensisin Mao’ershan Mountain[J].GuangxiForestryScienceandTechnology, 1992, 21(1): 24-26.
[24]罗金旺. 福建光泽南方铁杉天然林的生长规律与生物量[J]. 福建林学院学报, 2011, 31(2): 156-160.
LUO J W. Growth and biomass ofTsugachinensisvar.tchekiangensisin a natural forest in Guangze, Fujian, China[J].JournalofFujianCollegeofForestry, 2011, 31(2): 156-160.
[25]祁红艳, 金志农, 等. 江西武夷山南方铁杉生长规律及更新困难的原因解释[J]. 江西农业大学学报, 2014, 36(1): 137-143.
QI H Y, JIN Z N,etal. Growing law and cause of poor regeneration ofTsugachinensisvar.tchekiangensisin Jiangxi Wuyishan National Nature Reserve[J].ActaAgriculturaeUniversitatisJiangxiensis, 2014, 36(1): 137-143.
[26]张强, 郭传友, 张兴旺, 等. 基于光合作用和抗氧化机制的南方铁杉和褐叶青冈越冬策略研究[J].BulletinofBotanicalResearch, 2015, 35(2): 200-207.
ZHANG Q, GUO C Y, ZHANG X W,etal. Photosynthesis and antioxidant defense strategies in overwintering plants ofTsugachinensisandCyclobalanopsisstewardiana[J].BulletinofBotanicalResearch, 2015, 35(2): 200-207.
[27]张志祥, 刘鹏, 等. 不同群落类型下南方铁杉金属元素含量差异及其与土壤养分因子的关系[J]. 植物生态学报, 2010, 34(5): 505-516.
ZHANG Z X, LIU P,etal. Metal element contents ofTsugachinensisvar.tchekiangensisin different community types and its relationship with soil nutrient factors in eastern China[J].ChineseJournalofplantEcology, 2010, 34(5): 505-516.
[28]吴九玲, 钱晓鸣, 刘燕. 南方铁杉外生菌根的扫描电镜观察[J]. 厦门大学学报 (自然科学版) , 2001, 40(6): 1 337-1 341.
WU J L, QIAN X M, LIU Y. SEM-observation of ectomycorrhizal outer surface ofTsugachinensisvar.tchekiangensis(Flous) Cheng[J].JournalofXiamenUniversity(Natural Science Edition), 2001, 40(6): 1 337-1 341.
[29]钱晓鸣, 黄耀坚, 等. 武夷山自然保护区南方铁杉外生菌根生物多样性[J]. 福建农林大学学报 (自然科学版), 2007, 36(2): 180-185.
QIAN X M, HUANG Y J,etal. Biodiversity of the ectomycorrhiza on the rare and endangered tree speciesTugachinensistchekiangensis(Flous) Cheng in Wuyishan Nature Reserve[J].JournalofFujianAgricultureandForestryUniversity(Natural Science Edition), 2007, 36(2): 180-185.
[30]李晓铁, 玉伟朝, 罗远周, 等. 南方铁杉扦插繁殖技术[J]. 林业实用技术, 2008, 6: 21-22.
LI X T, YU W C, LUO Y Z,etal. Cutting propagation techniques ofTugatchekiangensis[J].PracticalForestryTechnology, 2008, 6: 21-22.
[31]何建源, 卞羽, 吴焰玉, 等. 南方铁杉林林隙自然干扰规律[J]. 西南林学院学报, 2009, 29(6): 7-10.
HE J Y, BIAN Y, WU Y Y,etal. Study on the law of natural disturbance in the gaps ofTsugachekiangensisfores[J].JournalofSouthwestForestryUniversity, 2009, 29(6): 7-10.
[32]侯碧清. 湖南酃县桃源洞自然保护区综合考察报告[M]. 长沙: 国防科技大学出版社, 1993. 3-20.
[33]谭益民, 吴章文. 桃源洞国家级自然保护区的生态状况[J]. 林业科学, 2009, 45(7): 52-58.
TAN Y M, WU Z W. Ecological status of taoyuandong national Nature Reserve[J].ScientiaSilvaeSinicae, 2009, 45(7): 52-58.
[34]王伯荪, 余世孝, 彭少麟, 等. 植物群落学实验手册[M]. 广州: 广东高等教育出版社, 1996. 1-56.
[35]马克平, 黄建辉, 于顺利, 等. 北京东灵山地区植物群落多样性的研究:II. 丰富度、均匀度和物种多样性指数[J]. 生态学报, 1995, 15(3): 268-277.
MA K P, HUANG J H, YU S L,etal. Plant community diversity in Dongling Mountain, Beijing, China: II. species richness, evenness and species diversities[J].ActaEcologicaSinica, 1995, 15(3): 268-277.
[36]江洪. 云杉种群生态学研究[M]. 北京: 中国林业出版社, 1992.
[37]WRETTEN S. Field and Laboratory Exercises in Ecology [M]. London: Edward Arnad Publishers Limited, 1980.
[38]杨凤翔, 王顺庆, 徐海根, 等. 生存分析理论及其在研究生命表中的应用[J]. 生态学报, 1991, 11(2): 153-158.
YANG F X, WANG S Q, XU H G,etal. The theory of survival analysis and its application to life table[J].ActaEcologicaSinica, 11(2): 153-158.
[39]陈灵芝, 孙航, 郭柯. 中国植物区系及植被地理[M]. 北京: 科学出版社, 2015. 413-438.
[40]张志祥. 九龙山自然保护区珍惜濒危植物南方铁杉种群生态学研究[D]. 浙江金华: 浙江师范大学, 2009.
(编辑:潘新社)
文章编号:1000-4025(2016)06-1233-12
doi:10.7606/j.issn.1000-4025.2016.06.1233
收稿日期:2016-03-10;修改稿收到日期:2016-04-29
基金项目:国家科技基础性工作专项(2013FY111500);2015年度中山大学与广东省本科教学质量和教学改革项目
作者简介:丁巧玲(1986-),女,博士,助理研究员,主要从事生态学和古生物学研究。E-mail:qlding@126.com *通信作者:石祥刚,博士,讲师,主要从事植物系统分类与植物资源学研究。E-mail:lsssxg@mail.sysu.edu.cn
中图分类号:Q948.15+7
文献标志码:A
Structure and Survival Analysis ofTsugachinensisPopulations in Taoyuandong National Nature Reserve, Hunan Province
DING Qiaoling1, LIU Zhongcheng2, WANG Lei2, CAI Songchen1,SHI Xianggang1*, ZENG Maosheng3, LIAO Wenbo1
(1 School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China; 2 College of Resource Environment and Tourism, Capital Normal University, Beijing 100048, China; 3 Administration Bureau of Taoyuandong National Nature Reserve, Yanling, Hunan 412500, China)
Abstract:Two T. chinensis communities, from Niushiping and Lishuzhou in Taoyuandong National Nature Reserve, are studied by using the methods of plots for the composition and structure of the communities, using Shannon-Weiner index, Simpson index and Pielou index for species diversity of every layers and using diameter class frequency distribution of T. chinensis to analyze the age class frequency distribution and the regeneration of the target population. The results are as follows: (1) since the evergreen plants are dominant, the physiognomy of the two communities show some seasonal changes of dark green in summer and light yellow-green in autumn and winter. (2) Twenty-six species in 21 genera of 13 families compose the T. chinensis community of Niushiping and the T. chinensis community of Lishuzhou contains 22 families, 30 genera with 36 species. Their proportions of temperate genera of seed plants are higher than that of tropical genera, which shows the subtropical montane property of geographical elements. (3) The vertical structure of above two communities can be divided into three layers, i.e. tree layer, shrub layer and herb layer and the tree layer includes three sub-layers, i.e. upper layer, middle layer and lower layer. (4) Shannon-Wiener index and Simpson index arrange as lower layer of tree > middle layer of tree > shrub layer > upper layer of tree in both communities, and Pielou index of middle layer of tree and lower layer of tree is higher than that of upper layer of tree and shrub layer in both communities. (5) The dominant populations of the T. chinensis community in Niushiping are Rhododendron ovatum, T. chinensis, Camellia cuspidata and Rhododendron latoucheae while the dominant populations of the T. chinensis community in Lishuzhou are Schima argentea, T. chinensis, Rhododendron latoucheae and Rhododendron hypoblematosum. (6) According to the age frequency distribution and the survival analysis, the T. chinensis population from Niushiping is a declining population and unable to regenerate itself because of lacking of enough younger age individuals; the T. chinensis population from Lishuzhou is a increasing population with a certain number of younger age individuals and relatively reasonable age structure while the high canopy density could be a limiting factor that challenge its natural regeneration. In conclusion, we suggest that in situ conservation and proper human intervention are necessary for the T. chinensis community of Niushiping and further monitoring is essential for the T. chinensis community of Lishuzhou.
Key words:Tsuga chinensis; community structure; population structure; survival analysis; population regeneration; Taoyuandong of Hunan Province
