滇桂两省中越边境地区藓类物种多样性及区系研究
2016-07-05章博远韦玉梅魏倩倩朱瑞良王幼芳
章博远,韦玉梅,李 敏,魏倩倩,朱瑞良,王幼芳*
(1 华东师范大学 生命科学学院,上海 200241;2 中国科学院 广西植物研究所,广西桂林 541006;3 河北师范大学 生命科学学院,石家庄 050024)
滇桂两省中越边境地区藓类物种多样性及区系研究
章博远1,韦玉梅2,李敏3,魏倩倩1,朱瑞良1,王幼芳1*
(1 华东师范大学 生命科学学院,上海 200241;2 中国科学院 广西植物研究所,广西桂林 541006;3 河北师范大学 生命科学学院,石家庄 050024)
摘要:对2012~2015年间采集自滇、桂两省中越边境地区的2 600余份藓类植物标本进行物种鉴定和文献研究,分析了中越边境地区藓类植物的物种组成和区系特点,并将中越边境地区与邻近地区和国家的藓类植物进行物种丰富度、属种相似性和区系谱的比较及聚类分析。结果表明:(1)滇、桂两省中越边境地区共有藓类植物51科、155属、452种(含种下单位),其中卷边麻锦藓[Taxithelium kerianum (Broth.) Broth.]为中国新记录种。(2)中越边境地区的藓类植物共有优势科9科,优势属12属。(3)中越边境地区的藓类植物区系以热带亚洲成分所占比例最高,达31.19%,其次为东亚成分(25.46%)和北温带广布成分(13.76%)。(4)与中越边境地区的藓类植物属及种的相似性最高的分别是越南和中国贵州。(5)中越边境地区的藓类植物区系与贵州、广东、福建及台湾的亲缘关系较近。
关键词:广西;云南;越南;物种多样性;藓类区系;热带
滇、桂两省中越边境地区(简写为中越边境地区)是指中国的云南省(滇)和广西壮族自治区(桂)与越南交界的地区。中越两国陆地边界线西起中国、越南、老挝三国的交界点,东至北仑河口止,全长约1 065km;中国和越南分别位于边界线的北面和南面。中越边境地区的地形地貌特征错综复杂:云南省与越南的交界地区位于该省的东南部,此地区的海拔较该省西北部要低,形成海拔2 000m左右的高原,河流呈帚状分布,河口瑶族自治县附近的元江河谷海拔仅76m左右,堪称云南海拔最低处,河流发育较好的区域则形成顶部平坦宽阔的山地;广西自治区与越南的交界地区位于该区的西南部,广西南部多山地,包括云开大山、六万大山、十万大山等山脉,虽名为大山,但实际海拔不过1 000m左右,山间多峡谷、急流,除此之外,还分布有一些岩溶地貌,地理状况亦是相当复杂。
中越边境地区大致位于北回归线(N23° 26′)以南,西南临近北部湾,属于热带范围,但是又处于热带的北缘,紧邻北温带,距离热带的中心区域(赤道附近)较远。故该地区以热带-亚热带季风气候为主,兼受印度洋和太平洋季风的影响,热量充足、雨量充沛,一年可分为明显的旱季和雨季。中越边境地区的气候地理特征使得该地区的生物多样性指数极高,大围山、黄连山、分水岭、十万大山、弄岗等多个国家级自然保护区都位于该区域内。
对于中越边境地区的苔藓植物,研究最多的地区是云南大围山[1-4],此外云南木洪大山[5]、广西十万大山[6-7]以及广西那坡县[8]和龙州县[9]也有过调查;韦玉梅[10]调查了中国广西与越南接壤的岩溶地区的苔藓植物;黄乐乐[11]则调查了中国境内中越边境地区的苔类植物区系。可见前人开展的关于该地区藓类植物的调查研究都是针对其中的一些片区进行的。到目前为止,关于中越边境地区完整的苔藓植物区系调查还从未开展过。
基于该地区苔藓植物的研究现状,本研究首次针对中越边境地区的藓类植物,对其进行较为完整的区系研究,以期完善中国在中越边境地区藓类植物调查的不足之处。另外,在中国苔藓植物十大分区中,该地区位于岭南区内,是中国苔藓植物种类和生态类型最为丰富的地区[12],具有很高的研究价值。此外,中国大部分国土位于北温带,该地区是中国境内为数不多的属于热带的地区,因此可通过对滇、桂两省中越边境地区藓类植物区系的研究,探讨中国苔藓植物区系与热带苔藓植物区系之间的亲缘关系,同时也为苔藓植物地理分布区的划定提供资料和佐证。
1材料和方法
1.1野外考察和标本鉴定
2012年至2015年间,作者对云南省和广西壮族自治区的中越边境地区进行了9次野外考察和标本采集,采集地点(图1)包括:云南的绿春县、元阳县、金平县、屏边县、河口县、马关县、麻栗坡县、西畴县、富宁县,以及广西的那坡县、靖西县、大新县、龙州县、凭祥市(县级)、宁明县、上思县、防城港市防城区,共计17个采集地。共获得藓类植物标本2 612号,标本均存放于华东师范大学标本馆(HSNU)。
将每份标本在解剖镜和光学显微镜下进行镜检,参考相关文献和专著对标本进行物种鉴定[13-25],具体种名及系统位置以《中国生物物种名录》第一卷的苔藓植物部分[12]为标准,并参照中国种子植物分布区类型的划分[26-27]和每个种的分布范围确定其区系成分。
1.2数据分析
采用物种数和Gleason物种丰富度指数DGl作为生物多样性的指标[28],利用Kroeber系数[29]和区系谱[30]将中越边境地区与邻近地区和国家的藓类植物进行属种相似性及区系的比较,采用SPSS19.0软件[31-33]对中越边境地区与邻近地区、国家的藓类植物区系地理成分进行聚类分析。
(1)Gleason物种丰富度指数
DGl= (S-1)/lnA,
式中S为藓类植物物种数,A为面积。
(2)Kroeber相似性系数
S = c(a + b) / 2ab×100%,式中a、b为两地各自物种数,c为共有种数;a、b、c均不含世界广布种。
(3)区系谱
区系谱是指某一特定植物区系中各区系成分百分率的合集。
FER = (FEi/T) × 100%,式中FEi指某区系中n种区系成分的第i个区系成分的分类群数量(属或种),T指某区系中分类群总数;FEi、T均不包括世界分布类型的种类。
(4)SPSS软件聚类分析
以类间平均锁链法(Between-groupslinkage)进行聚类分析,对间距的测量方法选择皮尔逊相关系数(Pearson-correlation)。
2结果与分析
2.1中越边境地区藓类植物物种多样性分析
2.1.1科、属、种的组成通过对2 600余份标本的显微鉴定,发现中越边境地区共有藓类植物51科、155属、452种(含种下单位)。其中,卷边麻锦藓[Taxithelium kerianum (Broth.)Broth.]为中国新记录种[34]。
2.1.2优势科将科内种数≥20种的科定义为优势科(表1)。中越边境地区共有优势科9科,共包含67属,251种,分别占该地区总属数和总种数的43.23% 和55.53%。优势科中,除广布的灰藓科(Hypnaceae)和真藓科(Bryaceae)以及温带分布为主的青藓科(Brachytheciaceae)和丛藓科(Pottiaceae)外,主要是以热带、亚热带分布为主的蔓藓科(Meteoriaceae)、平藓科(Neckeraceae)、白发藓科(Leucobryaceae)、凤尾藓科(Fissidentaceae)和毛锦藓科(Pylaisiadelphaceae)。优势科的组成特点体现了中越边境地区藓类植物区系热带和温带成分交汇的特色,并已趋近于热带植物区系。这一结果与该地区所处的纬度位置相一致。此外,中越边境地区仅有单种分布的科有13科,分别为短颈藓科(Diphysciaceae)、紫萼藓科(Grimmiaceae)、曲背藓科(Oncophoraceae)、蔓枝藓科(Bryowijkiaceae)、壶藓科(Splachnaceae)、稜蒴藓科(Ptychomniaceae)、油藓科(Hookeriaceae)、毛枝藓科(Pilotrichaceae)、碎米藓科(Fabroniaceae)、万年藓科(Climaciaceae)、异齿藓科(Regmatodontaceae)、白齿藓科(Leucodontaceae)和金毛藓科(Myuriaceae)。它们也以暖温带和热带、亚热带成分为主。
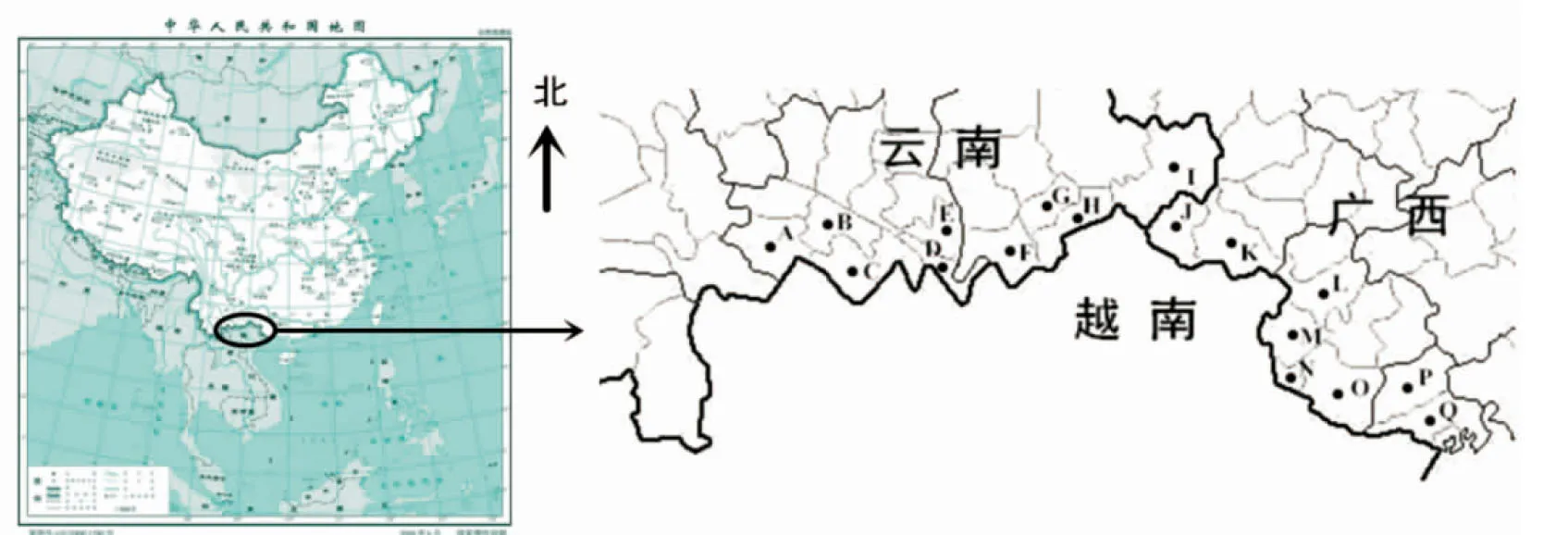
A.绿春县;B.元阳县;C.金平县;D.河口县;E.屏边县;F.马关县;G.西畴县;H.麻栗坡县;I.富宁县;J.那坡县;K.靖西县;L.大新县;M.龙州县;N.凭祥市;O.宁明县;P.上思县;Q.防城港市防城区图1 中越边境地区藓类植物采集地分布图A.Lvchun County; B.Yuanyang County; C.Jinping County; D.Hekou County; E.Pingbian County; F. Maguan County;G. Xichou County; H.Malipo County; I.Funing County; J.Napo County; K.Jingxi County; L.Daxin County; M.Longzhou County; N.Pingxiang City; O.Ningming County; P.Shangsi County; Q.Fangcheng District of Fangchenggang CityFig. 1 Map of Sino-Vietnam Border Area (Yunnan & Guangxi, China) with moss collection localities
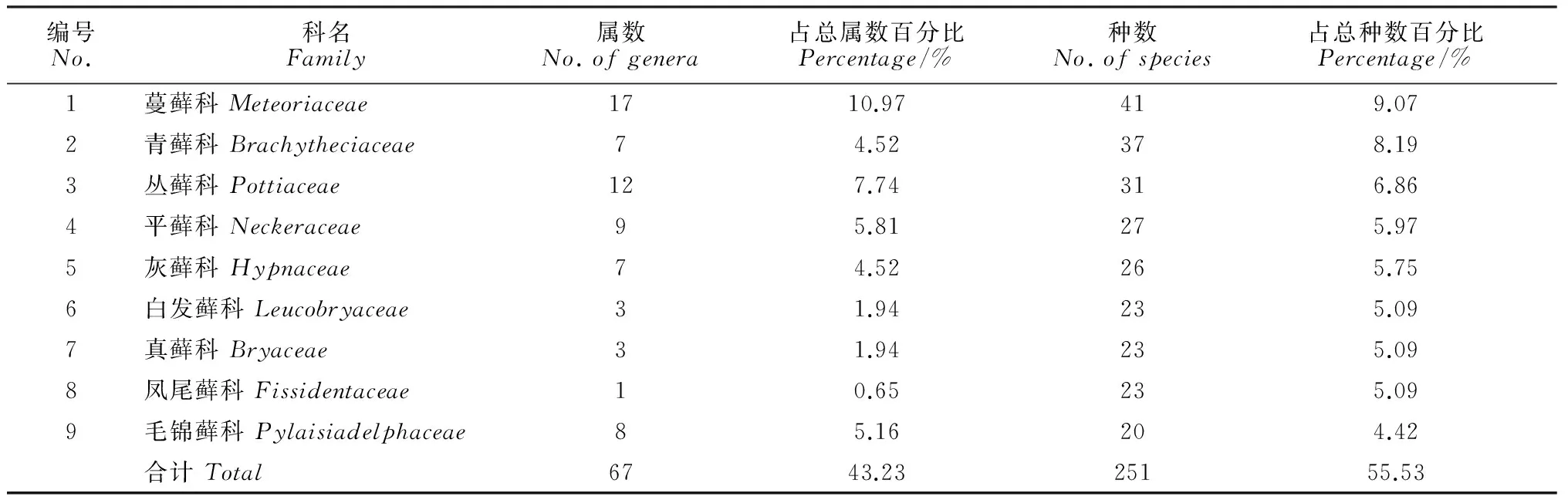
编号No.科名Family属数No.ofgenera占总属数百分比Percentage/%种数No.ofspecies占总种数百分比Percentage/%1蔓藓科Meteoriaceae1710.97419.072青藓科Brachytheciaceae74.52378.193丛藓科Pottiaceae127.74316.864平藓科Neckeraceae95.81275.975灰藓科Hypnaceae74.52265.756白发藓科Leucobryaceae31.94235.097真藓科Bryaceae31.94235.098凤尾藓科Fissidentaceae10.65235.099毛锦藓科Pylaisiadelphaceae85.16204.42合计Total6743.2325155.53
2.1.3优势属对中越边境地区155属藓类进行属内种数多寡的统计,属的组成见表2。属内种数≥10种的属有7属,仅占该地区总属数的4.52%;而少种属(属内种数少于5种)和单种属所占比例较高,尤其以单种属为最,达66属,占该地区总属数42.58%,而种的结构总体上较为均匀。上述结果体现出中越边境地区藓类植物属的多样性及区系的复杂性。
将属内种数≥7种的属定义为优势属(表3)。中越边境地区藓类共有优势属12属,包含130种,占该地区藓类植物总种数的28.76%。优势属的组成和优势科有一定的相似性,既包括广布性质的真藓属(Bryum),温带性质的匐灯藓属(Plagiomnium)、青藓属(Brachythecium)、美喙藓属(Eurhynchium)和绢藓属(Entodon),也有兼具温、热性质的曲柄藓属(Campylopus)、泽藓属(Philonotis)和长喙藓属(Rhynchostegium)以及热带性质的白发藓属(Leucobryum)、凤尾藓属(Fissidens)、树平藓属(Homaliodendron)和拟平藓属(Neckeropsis)。值得注意的是仅凤尾藓属一属就包含23种,占该地区藓类植物总种数的5.09%,是该地区优势性最为明显的属。中越边境地区藓类植物区系的热带性质由此可见一斑。
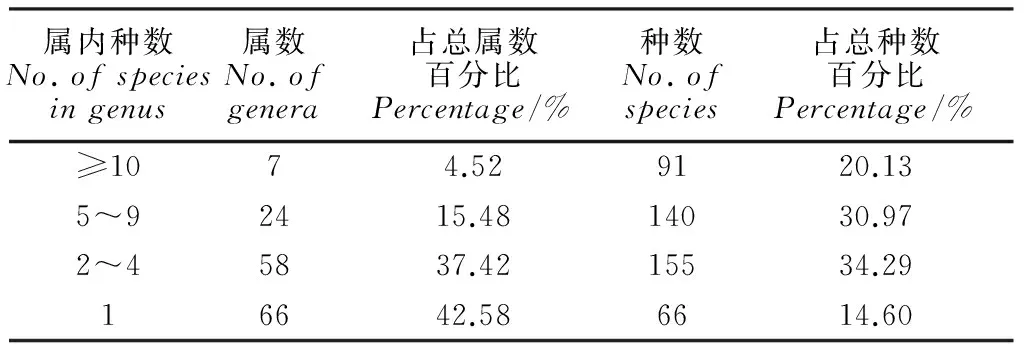
表2 中越边境地区藓类植物属的组成
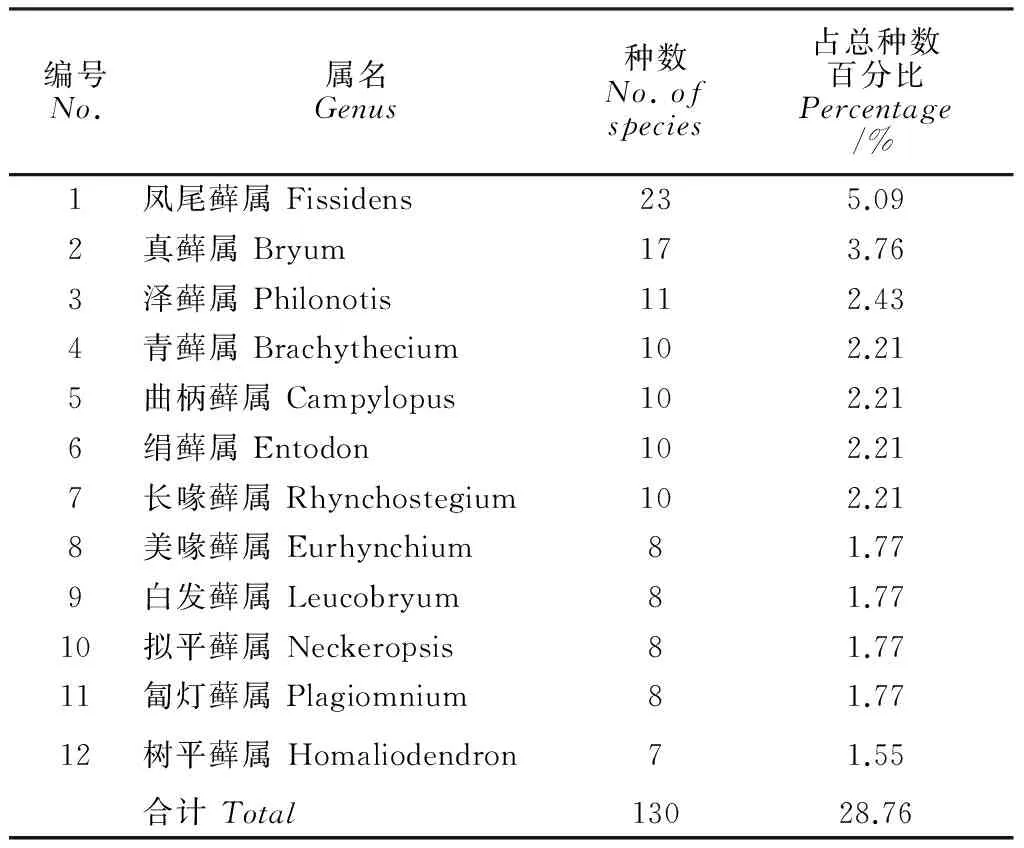
表3 中越边境地区藓类植物优势属
2.2中越边境地区藓类植物区系地理成分分析
2.2.1藓类植物区系成分的构成参照中国种子植物分布区类型的划分[26-27],中越边境地区藓类植物的区系成分共包括以下12种类型(表4):
(1)世界分布(Cosmopolitan):指分布范围几乎遍布世界各个大洲而没有特殊分布中心,或虽有一个或数个分布中心同时又广布于世界各地的种。属于这一分布类型的种有:丛生真藓(Bryum caespiticiumHedw.)、曲柄藓[Campylopus flexuosus (Hedw.)Brid.]、鳞叶凤尾藓(Fissidens taxifoliusHedw.)、大羽藓[Thuidium cymbifolium (Dozy&Molk.)Dozy&Molk.]、长蒴藓(Trematodon longicollisMichx.)等。
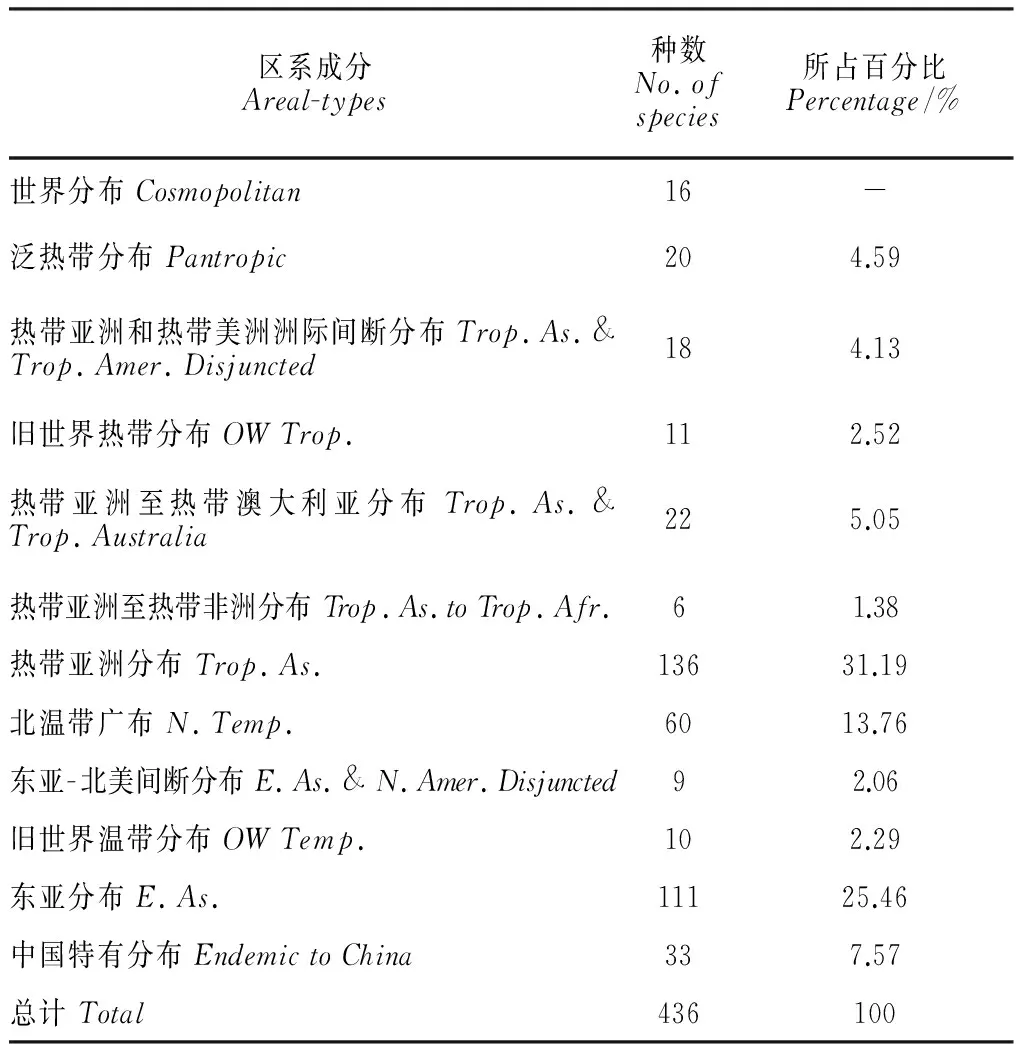
表4 中越边境地区藓类植物区系成分
注:世界分布成分未计入总数。
Note:CosmopolitanSpeciesareexcluded.
(2)泛热带分布(Pantropic):指广泛分布于环球的热带地区的种,通常有亚、澳,非、印和中南美三大分布中心。属于这一分布类型的种有:蕊形真藓(Bryum coronatumSchwägr.)、短肋凤尾藓(Fissidens gardneriMitt.)、长柄蓑藓[Macromitrium microstomum (Hook. &Grev.)Schwägr.]、黄色斜齿藓[Mesonodon flavescens (Hook.)W.R.Buck]、匍网藓[Mitthyridium fasciculatum (Hook. &Grev.)H.Rob.]等。
(3)热带亚洲和热带美洲洲际间断分布(TropicalAsia&TropicalAmericaDisjuncted):指间断分布于美洲和亚洲温暖地区的热带种,在东半球可延伸到澳大利亚东北部或西南太平洋岛屿。属于这一分布类型的种有:近高山真藓(Bryum paradoxumSchwägr.)、梯网花叶藓(Calymperes afzeliiSw.)、锦叶藓[Dicranoloma dicarpum (Nees)Paris]、网孔凤尾藓(Fissidens polypodioidesHedw.)、狭叶白发藓(Leucobryum bowringiiMitt.)等。
(4)旧世界热带分布(OldWorldTropics):指分布范围在亚洲、大洋洲、非洲三大洲的热带地区的种。属于这一分布类型的种有:宽叶短月藓[Brachymenium capitulatum (Mitt.)Paris]、尾枝藓[Caduciella mariei (Besch.)Enroth]、穗枝赤齿藓[Erythrodontium julaceum (Hook.exSchwägr.)Paris]、丝带藓[Floribundaria floribunda (Dozy&Molk.)M.Fleisch.]、小树平藓[Homaliodendron exiguum (Bosch&SandeLac.)M.Fleisch.]等。
(5)热带亚洲至热带澳大利亚分布(TropicalAsiatoTropicalAustralia):指分布范围在热带亚洲和大洋洲的种,西端可达马达加斯加,但不见于非洲大陆。属于这一分布类型的种有:灰气藓[Aerobryopsis wallichii (Brid.)M.Fleisch.]、拟悬藓[Barbellopsis trichophora (Mont.)W.R.Buck]、疏网曲柄藓(Campylopus laxitextusSandeLac.)、大麻羽藓[Claopodium assurgens (Sull. &Lesq.)Cardot]、淡色同叶藓[Isopterygium albescens (Hook.)A.Jaeger]等。
(6)热带亚洲至热带非洲分布(TropicalAsiatoTropicalAfrica):指分布范围在热带非洲至印度-马来西亚的种,部分种也可分布到斐济等南太平洋岛屿,但不见于澳大利亚大陆。属于这一分布类型的种有:短月藓(Brachymenium nepalenseHook.)、刀叶树平藓[Homaliodendron scalpellifolium (Mitt.)M.Fleisch.]、阔叶桧藓[Pyrrhobryum latifolium (Bosch&SandeLac.)Mitt.]、南亚火藓(Schlotheimia grevilleanaMitt.)、矮刺果藓[Symphyodon pygmaeus (Broth.)S.He&Snider]等。
(7)热带亚洲分布(TropicalAsia):该分布型在中国境内的分布范围从南岭以南至南海诸岛,境外的分布范围则包括热带喜马拉雅、东南亚诸国,东达新几内亚、西太平洋,南不到澳大利亚北部的热带部分。属于这一分布类型的种有:针叶顶胞藓[Acroporium diminutum (Brid.)M.Fleisch.]、毛扭藓[Aerobryidium filamentosum (Hook.)M.Fleisch.]、气藓(Aerobryum speciosumDozy&Molk.)、爪哇扭口藓(Barbula javanicaDozy&Molk.)、赤茎小锦藓[Brotherella erythrocaulis (Mitt.)M.Fleisch.]等。
该天然气的低位发热量为Qdr=36.59MJ/Nm3,高位发热量为Qgr=40.52 MJ/Nm3,两者相差为3.93 MJ/Nm3,该差值即为烟气中水蒸气的汽化潜热。在一般燃气锅炉排烟温度下,水蒸气仍处于过热状态,因而由锅炉排烟所带走的汽化潜热占低位发热量的比例可用式1求得。
(8)北温带广布(NorthTemperate):指分布在欧洲、亚洲和北美洲温带地区的种,有些种甚至跨入南温带或两极地区。属于这一分布类型的种有:柳叶藓[Amblystegium serpens (Hedw.)Schimp.]、灰白青藓[Brachythecium albicans (Hedw.)Schimp.]、高山红叶藓[Bryoerythrophyllum alpigenum (Venturi)P.C.Chen]、卵蒴真藓(Bryum blindiiBruch&Schimp.)、辛氏曲柄藓(Campylopus schimperiMilde)等。
(9)东亚-北美间断分布(EastAsia&NorthAmericaDisjuncted):指分布中心在东亚和北美洲温带及亚热带的种。属于这一分布类型的种有:中华曲柄藓[Campylopus sinensis (Müll.Hal.)J.-P.Frahm]、羊角藓[Herpetineuron toccoae (Sull. &Lesq.)Cardot]、黄边孔雀藓(Hypopterygium flavolimbatumMüll.Hal.)、扭叶小金发藓[Pogonatum contortum (MenziesexBrid.)Lesq.]、凸尖鳞叶藓(Taxiphyllum cuspidifolium (Cardot)Z.Iwats.)等。
(10)旧世界温带分布(OldWorldTemperate):亦称欧亚温带分布,指广泛分布在欧洲、亚洲中高纬度的温带和寒温带的种。属于这一分布类型的种有:小疣毛藓[Leratia exigua (Sull.)B.Goffinet]、深绿褶叶藓[Palamocladium euchloron (BruchexMüll.Hal.)Wijk&Margad.]、偏叶泽藓[Philonotis falcata (Hook.)Mitt.]、日本立碗藓[Physcomitrium japonicum (Hedw.)Mitt.]、阔叶棉藓(Plagiothecium platyphyllumMönk.)等。
(11)东亚分布(EastAsia):指分布范围在亚洲东北部的种,北以中国内蒙古的阴山和狼山为界,南到泰国东北部、老挝和越南的北部。属于这一分布类型的种有:小牛舌藓[Anomodon minor (Hedw.)Lindb.]、东亚仙鹤藓[Atrichum yakushimense (Horik.)Mizush.]、柔叶青藓(Brachythecium morienseBesch.)、弯叶真藓[Bryum recurvulumMitt.]、毛叶梳藓[Ctenidium capillifolium (Mitt.)Broth.]等。
(12)中国特有分布(EndemictoChina):指分布范围在中国境内的种,有些种则越出国界达到邻国边界。属于这一分布类型的种有:云南灰气藓(Aerobryopsis yunnanensisX.J.Li&D.C.Zhang)、带叶耳平藓(Calyptothecium phyllogonioidesNog. &X.J.Li)、宝岛绢藓(Entodon taiwanensisC.K.Wang&S.H.Lin)、小叶美喙藓[Eurhynchium filiforme (Müll.Hal.)Y.F.Wang&R.L.Hu]、中华细枝藓[Lindbergia sinensis (Müll.Hal.)Broth.]等。
2.2.2藓类植物区系成分的特点由表4可知,中越边境地区藓类植物区系所包含的12种地理成分中,热带亚洲成分所占比例最高,达31.19%,其次为东亚成分和北温带广布成分,分别占25.46%和13.76%,由此可知中越边境地区的藓类植物区系已具明显的热带亚洲特色。亚热带至温带成分(包括北温带广布、东亚-北美间断分布、旧世界温带分布、东亚分布和中国特有分布)的总和所占比例达51.15%,而所有的热带成分(包括泛热带分布、热带亚洲和热带美洲洲际间断分布、旧世界热带分布、热带亚洲至热带澳大利亚分布、热带亚洲至热带非洲分布和热带亚洲分布)总和所占的比例亦达到48.85%,即亚热带至温带成分和热带成分这两大类成分在中越边境地区藓类植物区系中近于持平,温带性质的青藓科、丛藓科等科植物以及热带、亚热带性质的蔓藓科、平藓科、凤尾藓科、毛锦藓科等科植物在该地区有较多的分布,表明该地区的藓类植物区系兼具温带和热带特色。在中国植物区系分区系统中,中越边境地区属于古热带植物区中马来西亚亚区内的北部湾地区[27],是北温带植物区系和热带植物区系相互交融和影响的区域,并且是亚热带植物区系向热带植物区系过渡的一个关键地带。
2.3中越边境地区与邻近地区藓类植物区系比较
为进一步探明中越边境地区藓类植物区系的特点,本研究选取了中国境内的贵州[35]、广东[36]、海南[36]、福建[37]和台湾[38]5省以及地处南亚热带地区的缅甸[39]、泰国[40]、越南[41]和菲律宾[42]4国的藓类植物与之进行比较分析。
2.3.1藓类植物组成的比较表5为本研究从物种数、物种丰富度、共有科属种数和属种相似性系数等方面将中越边境地区的藓类植物与上述9个地区进行比较结果。
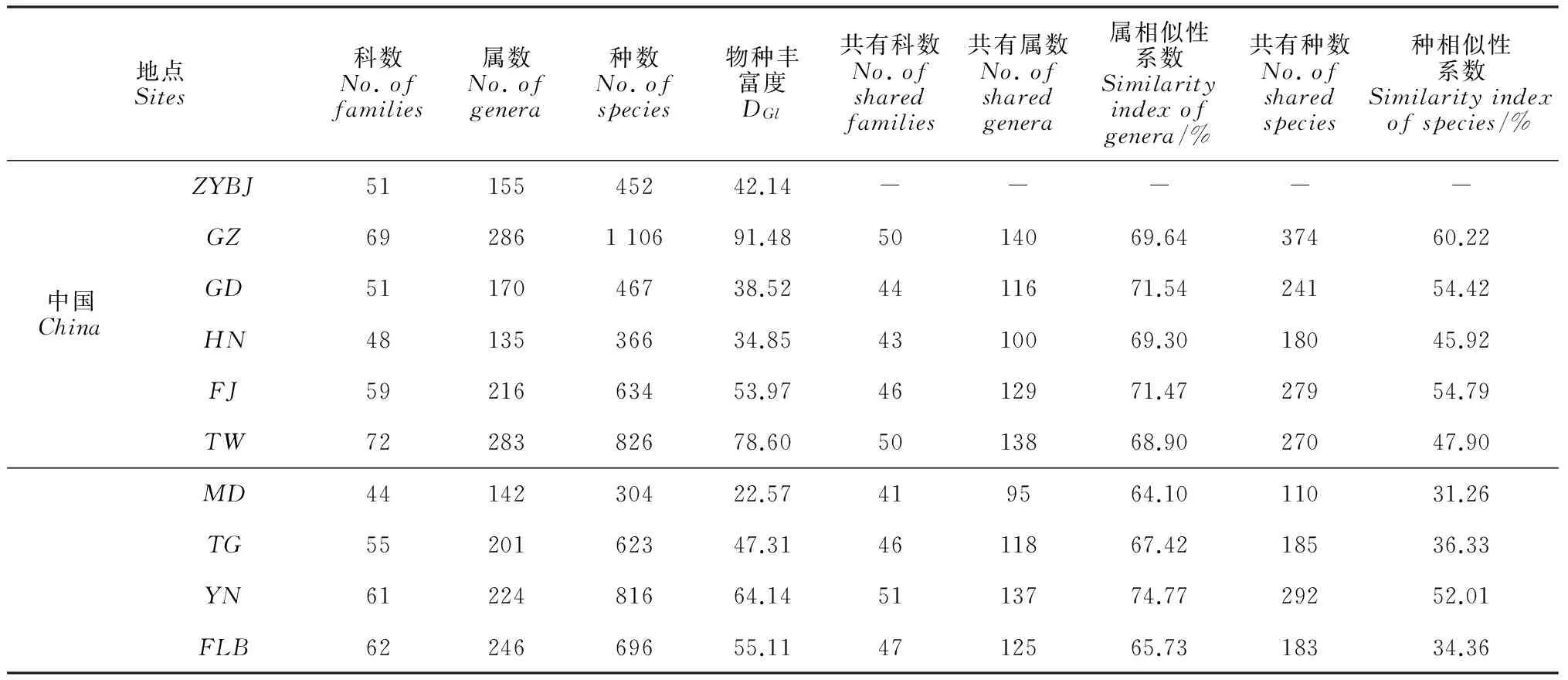
表5 中越边境地区与邻近地区藓类植物物种丰富度及属种相似性的比较
注:世界分布成分未计入。ZYBJ.中越边境;GZ.贵州;GD.广东;HN.海南;FJ.福建;TW.台湾;MD.缅甸;TG.泰国;YN.越南;FLB.菲律宾;下同。
Note:CosmopolitanSpeciesareexcluded.ZYBJ.Sino-Vietnamborder;GZ.Guizhou;GD.Guangdong;HN.Hainan;FJ.Fujian;TW.Taiwan;MD.Burma;TG.Thailand;YN.Vietnam;FLB.Philippines;Thesameasbelow.
(1) 物种丰富度比较:包括中越边境地区在内的10个地区中,物种丰富度较高的3个地区分别是中国贵州(91.48)、中国台湾(78.60)以及越南(64.14),而中越边境地区(42.14)位于第7位,仅高于中国广东、中国海南和缅甸。
(2) 属种组成的比较:从属种组成来看,所选取的国内外9个对比地与中越边境地区藓类植物属的相似性系数均达到60%以上。其中,越南(74.77%)、中国广东(71.54%)、中国福建(71.47%)3地达到70%以上,是与中越边境地区藓类植物属的相似性系数最高的对比地;而中国贵州则以69.64%的相似性系数位居第四。种的相似性系数较高的对比地分别为中国贵州(60.22%)、福建(54.79%)和广东(54.42%),而属的相似性系数最高的越南种的相似性系数仅为52.01%,位居第四。
单就属种组成而言,中越边境地区的藓类植物与中国的贵州、广东、福建以及邻国越南关系较近。从属的水平来看,越南与之最为接近;从种的水平来看,中国贵州与之表现出的相似性最高。越南位于北回归线以南,是南亚的热带国家;中国贵州则位于北回归线稍北,为典型的亚热带地区。由此可知,中越边境地区的藓类植物具有显著的热带、亚热带相互交汇融合的特色。
2.3.2藓类植物地理成分区系谱的比较与邻近地区进行地理成分区系谱的比较,能更为准确地揭示出中越边境地区藓类植物区系的本质。为便于比较,本研究将包括中越边境地区在内的6处中国境内的对比地所包含的中国特有成分并入东亚成分内,对比结果见表6。
包括中越边境地区在内的6处中国境内的对比地,其藓类植物区系均以东亚成分、热带亚洲成分和北温带成分为主,而国外的4处对比地的藓类植物区系则主要以热带亚洲成分、旧世界热带成分、热带亚洲至热带澳大利亚成分等一些热带成分为主。东亚成分、热带亚洲成分和北温带成分在10个藓类植物区系中所占的比例如图2所示。
东亚成分在中国境内对比地(除海南外)的藓类植物区系中均占有最高的比例,其中贵州(38.00%)、福建(34.64%)和台湾(33.50%)3地藓类区系的东亚成分所占比例较高,东亚成分在中越边境地区藓类区系中所占的比例为33.03%,高于广东(31.78%),表明中越边境地区藓类区系的东亚色彩不如贵州、福建、台湾等地浓厚。热带亚洲成分在中越边境地区藓类区系中所占的比例为31.19%,在中国境内的对比地中,这一比例仅低于海南(40.45%),高于贵州(18.81%)、福建(21.73%)、台湾(27.48%)和广东(29.33%),表明中越边境地区藓类区系的热带特色相较于贵州、福建、台湾等地更为显著。东亚成分和热带亚洲成分的差值与对比地所处的纬度位置形成了较好的对应关系,即随着纬度逐渐降低,贵州、福建、台湾、广东以及中越边境地区的东亚成分和热带亚洲成分的差值随之逐渐变小,表明该地区藓类植物区系的热带性质更趋显著;而海南,其热带亚洲成分所占比例则已超过东亚成分,这是由于海南位于N20°10′ ~ 18°10′ 之间,地处较为典型的热带地区,其藓类植物区系的热带色彩较为浓厚。
中国境外的缅甸、泰国、越南和菲律宾,因其地处南亚热带地区,因此热带亚洲成分在其藓类植物区系中占有绝对优势。这4国中,仅有越南的东亚成分所占比例稍高,这是因为越南与中国接壤,其植物区系受到以中国为代表的东亚植物区系的影响。而同与中国接壤的缅甸,其东亚成分所占比例仅为6.44%,究其原因,可能是因为缅甸的藓类植物名录资料较为陈旧,调查不够完善所导致,此点可从缅甸国土面积为越南的2倍以上,而藓类植物的物种丰富度却不足越南的一半看出。
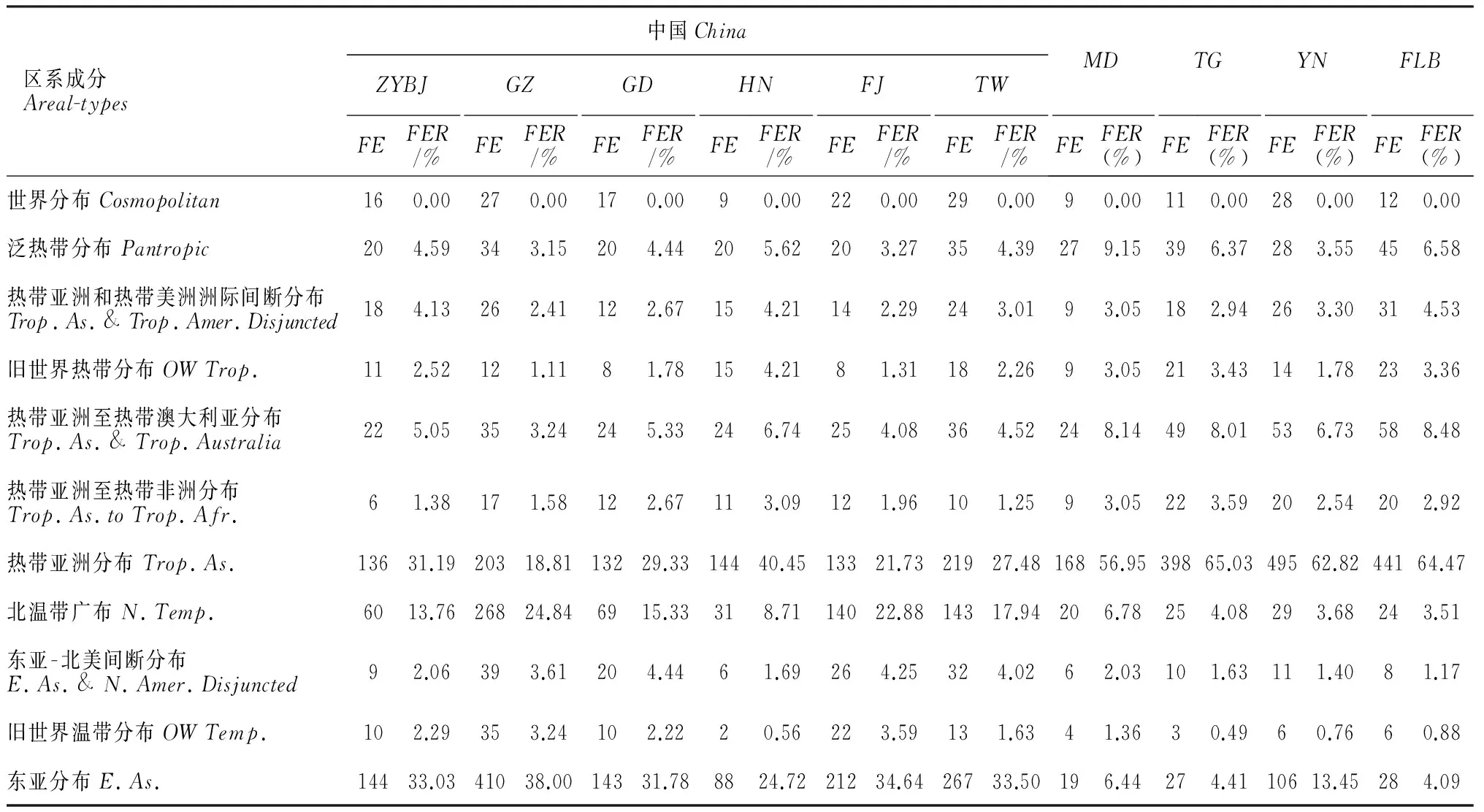
表6 中越边境地区与邻近地区藓类植物地理成分区系谱
注:FE表示某一特定区系成分所包含的种数,FER为该区系成分所占的百分比。
Note:FEmeansthenumberofspeciesofaspecificareal-types,FERmeansthepercentageoftheareal-types.
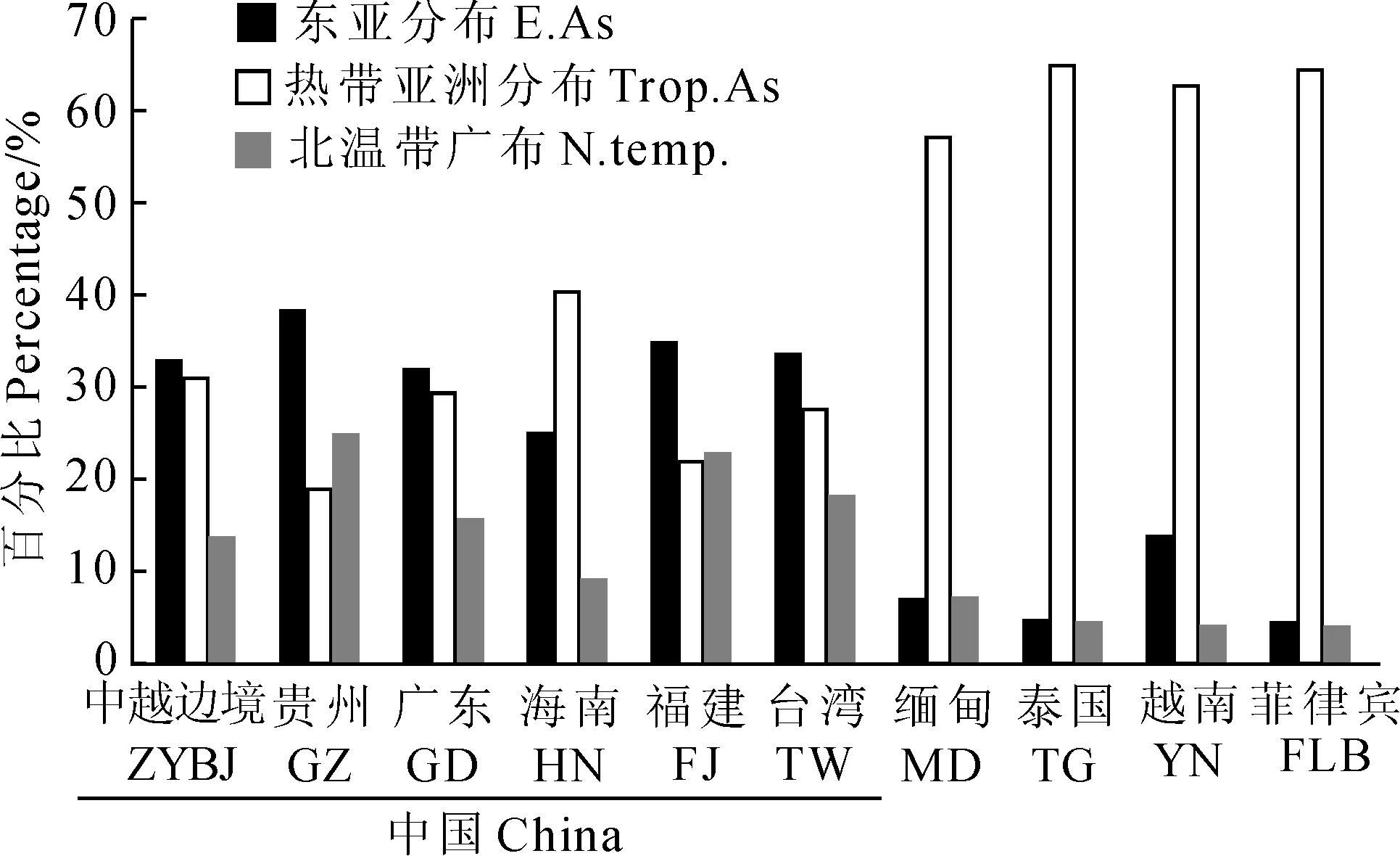
图2 10个地区3种主要区系成分比较Fig. 2 Comparison of three main areal-types of ten regions
总体来看,各区系成分所占的比例与该地所处的地理位置有较为明显的对应关系:纬度越高,其温带成分的比例越高;随着纬度的降低,温带成分逐渐减少,热带成分比例相应升高。
2.4中越边境地区与邻近地区藓类植物区系关系
运用SPSS19.0软件对包括中越边境地区在内的10个地区的藓类植物地理成分区系谱进行聚类分析。
由图3可知,10个地区被聚为两大分支:中国的贵州、福建、广东、台湾以及中越边境地区聚为一支,为亚热带至热带区系类群,缅甸、泰国、菲律宾、越南以及中国海南聚为一支,为典型热带区系类群。其中中国贵州、福建、广东、台湾4地的藓类植物区系关系较近,中越边境地区虽与它们聚为一支,但是距离相去较远。由此可见,中越边境地区的藓类植物区系与中国贵州、福建、广东、台湾4地关系不甚亲密。而中国海南则与缅甸、泰国、菲律宾和越南这些南亚热带国家聚为一支,表明中国海南是中国境内的典型热带地区。从整体来看,中越边境地区虽主要位于北回归线以南,在地理上属于热带地区的范围,但是由于其地处热带的北缘,紧邻北温带,植物区系受到温带植物区系的影响较大。因此,该地区的藓类植物区系与贵州、福建、台湾等地相比虽已表现出其热带特色,但却不如中国海南、泰国、越南等地所呈现的热带色彩浓厚。
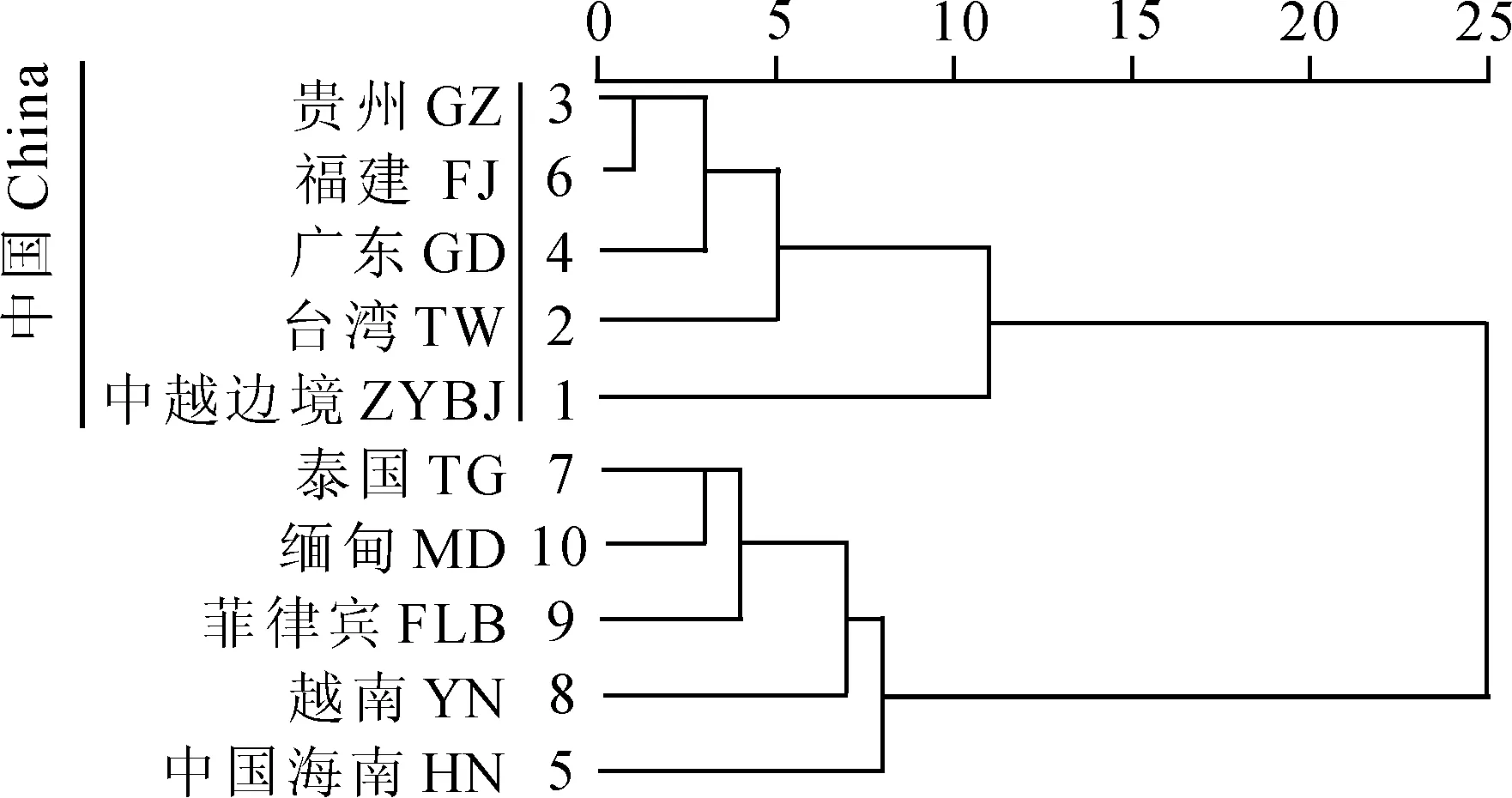
图3 10个地区藓类植物地理成分区系谱聚类分析Fig. 3 Cluster analysis of the floristic spectrum of moss flora of ten regions
综上所述,中越边境地区的藓类植物区系是热带和温带藓类植物区系相互交汇和影响的结果,兼具温热特色;中越边境地区是亚热带植物区系向热带植物区系过渡的地区。
参考文献:
[1]杨比伦, 肖银松. 大围山自然保护区苔藓植物资源[J]. 西南林学院学报, 2003, 23(2): 5-12.
YANGBL,XIAOYS.TheresourcesofbryophyteinDaweishanNatureReserve[J]. Journal of Southwest Forestry College, 2003, 23(2): 5-12.
[2]杨丽琼. 云南屏边大围山自然保护区藓类植物区系研究[D]. 上海: 华东师范大学, 2004.
[3]翟德逞, 王幼芳. 云南大围山自然保护区的叶附生苔类植物[J]. 西北植物学报, 2005, 25(9): 1 854-1 858.
ZHAIDC,WANGYF.EpiphyllousliverwortsintheDaweishanNatureReserveofYunnanprovince[J]. Acta Botanica Boreall-Occidentalia Sinica, 2005, 25(9): 1 854-1 858.
[4]翟德逞, 杨丽琼, 朱瑞良. 云南大围山常绿阔叶林内苔藓植物的研究[J]. 华东师范大学学报·自然科学版, 2005, (5/6): 188-197.
ZHAIDC,YANGLQ,ZHURL.StudyonbryophytesinevergreenbroadleavedforestsofDaweishanNatureReserve,YunnanProvince,China[J]. Journal of East China Normal University, (Nat.Sci.Edi.), 2005, (5/6): 188-197.
[5]徐力, 熊源新, 王美会, 等. 云南富宁县木洪大山苔藓植物区系研究[J]. 山地农业生物学报, 2010, 29(6): 475-481.
XUL,XIONGYX,WANGMH, et al.BryofloraatMuhongmountainofFuningYunnan,China[J]. Journal of Mountain Agriculture and Biology, 2010, 29(6): 475-481.
[6]ZHURL,SOML.LiverwortsandhornwortsofShangsicountyofGuangxi(Kwangsi),withanupdatedchecklistofthehepaticfloraofGuangxiProvinceofChina[J]. Cryptogamie, Bryologie, 2003: 24(4): 319-334.
[7]唐艳雪. 广西十万大山自然保护区苔藓植物区系及地理分布研究[D]. 上海: 上海师范大学, 2014.
[8]贾鹏, 熊源新, 王美会, 等. 广西那坡县苔藓植物初步研究[J]. 广西植物, 2011, 31(5): 627-635.
JIAP,XIONGYX,WANGMH, et al.PreliminarystudyonthebryoflorainNapoCountyofGuangxi[J]. Guihaia, 2011, 31(5): 627-635.
[9]只佳增. 广西龙州县苔藓植物初步研究[D]. 武汉: 华中师范大学, 2013.
[10]韦玉梅. 桂西南中越边境石灰岩地区苔藓植物的初步研究[D]. 桂林: 广西师范大学, 2009.
[11]黄乐乐. 中国境内中越边境地区苔类植物的区系研究[D]. 上海: 华东师范大学, 2015.
[12]贾渝, 何思. 中国生物物种名录第1卷(苔藓植物)[M]. 北京: 科学出版社, 2013.
[13]高谦. 中国苔藓志(第1卷)[M]. 北京: 科学出版社, 1994.
[14]高谦. 中国苔藓志(第2卷)[M]. 北京: 科学出版社, 1996.
[15]黎兴江. 中国苔藓志(第3卷)[M]. 北京: 科学出版社, 2000.
[16]黎兴江. 中国苔藓志(第4卷)[M]. 北京: 科学出版社, 2006.
[17]吴鹏程, 贾 渝. 中国苔藓志(第5卷)[M]. 北京: 科学出版社, 2011.
[18]吴鹏程. 中国苔藓志(第6卷)[M]. 北京: 科学出版社, 2002.
[19]胡人亮, 王幼芳. 中国苔藓志(第7卷)[M]. 北京: 科学出版社, 2005.
[20]吴鹏程, 贾 渝. 中国苔藓志(第8卷)[M]. 北京: 科学出版社, 2004.
[21]NOGUCHIA,IWATSUKIZ.IllustratedMossFloraofJapan(Part1)[M].Nichinan-shi:theHattoriBotanicalLaboratory, 1987.
[22]NOGUCHIA,IWATSUKIZ.IllustratedMossFloraofJapan(Part2)[M].Nichinan-shi:theHattoriBotanicalLaboratory, 1988.
[23]NOGUCHIA,IWATSUKIZ.IllustratedMossFloraofJapan(Part3)[M].Nichinan-shi:theHattoriBotanicalLaboratory, 1989.
[24]NOGUCHIA,IWATSUKIZ,YAMAGUCHIT.IllustratedMossFloraofJapan(Part4)[M].Nichinan-shi:theHattoriBotanicalLaboratory, 1991.
[25]NOGUCHIA,IWATSUKIZ,YAMAGUCHIT.IllustratedMossFloraofJapan(Part5)[M].Nichinan-shi:theHattoriBotanicalLaboratory, 1994.
[26]王荷生. 植物区系地理[M]. 北京: 科学出版社, 1992.
[27]吴征镒, 孙 航, 周浙昆, 等. 中国种子植物区系地理[M]. 北京: 科学出版, 2011.
[28]GLEASONHA.Ontherelationsbetweenspeciesandarea[J]. Ecology, 1992, 3: 158-162.
[29]胡晓云, 吴鹏程. 四川金佛山藓类植物区系的研究[J]. 植物分类学报, 1991, 29(4): 315-334.
HUXY,WUPC.StudyonthemossfloraofMt.Jinfu,SichuanProvince[J]. Acta Phytotaxonomica Sinica, 1991, 29(4): 315-334.
[30]马克平, 高贤明, 于顺利. 东灵山地区植物区系的基本特征与若干山区植物区系的关系[J]. 植物研究, 1995, 15(4): 501-515.
MAKP,GAOXM,YUSL.OnthecharacteristicsofthefloraofDonglingmountainareaanditsrelationshipwithanumberofothermountainousflorasinChina[J]. Bulletin of Botanical Research, 1995, 15(4): 501-515.
[31]陆志波, 陆雍森. 环境监测数据分析和监测网设计中SPSS10.0的应用[J]. 环境监测与技术, 2002, 14(3): 12-16.
LUZB,LUYS.ApplicationofSPSS10.0toenvironmentalmonitoringdataanalysisandnetworkdesign[J]. The Administration and Technique of Environmental Monitoring, 2002, 14(3): 12-16.
[32]吴文英, 左勤, 王幼芳, 等. 福建戴云山国家级自然保护区藓类植物区系研究[J]. 植物科学学报, 2012, 30(2): 122-132.
WUWY,ZUOQ,WANGYF, et al.StudyonthemossfloraoftheDaiyunshannationalnaturereserve,FujianProvince[J]. Plant Science Journal, 2012, 30(2): 122-132.
[33]魏倩倩, 买买提明·苏莱曼, 李 敏, 等. 中国新疆柳叶藓科、青藓科和灰藓科植物区系研究[J]. 西北植物学报, 2014, 34(2): 379-386.
WEIQQ,SULAYMANM,LIM, et al.StudyonthemossfloraofAmblystegiaceae,BrachytheciaceaeandHypnaceaeofXinjiangUygurAutonomousRegion,China[J]. Acta Botanica Boreall-Occidentalia Sinica, 2014, 34(2): 379-386.
[34]章博远, 魏倩倩, 韦玉梅, 等. 毛锦藓科中国新记录种——卷边麻锦藓[J]. 西北植物学报, 2015, 35(8): 1 690-1 693.
ZHANGBY,WEIQQ,WEIYM, et al. Taxithelium kerianum (Broth.)Broth. (Pylaisiadelphaceae,Bryophyta)reportednewtoChina[J]. Acta Botanica Boreall-Occidentalia Sinica, 2015, 35(8): 1690-1693.
[35]熊源新. 贵州苔藓植物志(第1卷、第2卷)[M]. 贵阳: 贵州科技出版社, 2014.
[36]吴德邻, 张力. 广东苔藓志[M]. 广州: 广东科技出版社, 2013.
[37]WUWY,WANGYF,ZUOQ, et al.AnupdatedchecklistofmossesofFujianProvince,China[J]. CHENIA, 2013, 11: 144-182.
[38]蒋镇宇, 许再文, 牟善杰, 等. 台湾藓类植物新名录[M]. 南投: 中国生物学会, 2001.
[39]TANBC,IWATSUKIZ.AchecklistofIndochinesemosses[J]. Journal of the Hattori Botanical Laboratory, 1993, 74: 325-405.
[40]HES.AnannotatedchecklistandatlasofthemossesofThailand[DB/OL].http://www.mobot.org/MOBOT/moss/Thailand/welcome.shtml
[41]HES,NGUYENSK.NewrecordsandanupdatedchecklistofthemossesofVietnam[J]. Tropical Bryology, 2012, 34: 32-88.
[42]TANBC,IWATSUKIZ.AnewannotatedPhilippinemosschecklist[J]. Harvard Papers in Botany 1991, 3: 1-64.
[43]陈邦杰. 中国苔藓植物生态群落和地理分布的初步报告[J]. 植物分类学学报, 1958, 7(4): 271-293.
CHENPC.PreliminaryreportofthecenologicalandgeographicalstudyofChinesemossflora[J]. Acta Phytotaxonomica Sinica, 1958, 7(4): 271-293.
[44]吴鹏程, 贾渝. 中国苔藓植物的地理及分布类型[J]. 植物资源及环境学报, 2006, 15(1): 1-8.
WUPC,JIAY.TheregionalizationanddistributiontypesofthebryophytesinChina[J]. Journal of Plant Resources and Environment, 2006, 15(1): 1-8.
(编辑:潘新社)
SpeciesDiversityandMossFloraofSino-VietnamBorderArea(Yunnan&Guangxi,China)
ZHANGBoyuan1,WEIYumei2,LIMin3,WEIQianqian1,ZHURuiliang1,WANGYoufang1*
(1Schooloflifesciences,EastChinaNormalUniversity,Shanghai200241,China; 2GuangxiInstituteofBotany,ChineseAcademyofSciences,Guilin,Guangxi541006,China; 3CollegeofLifeSciences,HebeiNormalUniversity,Shijiazhuang050024,China)
Abstract:More than 2 600 moss specimens were collected from Sino-Vietnam border area (Yunnan & Guangxi, China) during 2012 to 2015. Through the specimen identification and literature research, we analyzed the species composition and floristic characteristics of Sino-Vietnam Border Area (Yunnan & Guangxi, China), comparisons of moss DGland similarity index of genera & species along with floristic spectrum and the cluster analysis of Sino-Vietnam Border Area (Yunnan & Guangxi, China) with adjacent provinces and countries were made. Results are as follows: (1) the moss flora of Sino-Vietnam Border Area (Yunnan & Guangxi, China) consist of 452 species (including infraspecific taxa), belonging to 155 genera of 51 families. Taxithelium kerianum (Broth.) Broth. was reported new to China. (2) There are nine dominant families and 12 dominant genera in Sino-Vietnam Border Area (Yunnan & Guangxi, China). (3) The moss flora of Sino-Vietnam Border Area (Yunnan & Guangxi, China) is dominated by tropical Asia element, which accounts for 31.19% of the entire species. East Asia element (25.46%) and North Temperate element (13.76%) ranked second and third, respectively. (4)Vietnam has the highest similarity index of genera with Sino-Vietnam Border Area (Yunnan & Guangxi, China), whereas Guizhou(China) gets the highest similarity index of species within the studied area. (5) The moss flora of Sino-Vietnam Border Area (Yunnan & Guangxi, China) is more closely related to Guizhou, Guangdong, Fujian and Taiwan.
Key words:Guangxi; Yunnan; Vietnam; species diversity; moss flora; tropical
文章编号:1000-4025(2016)05-1021-010
doi:10.7606/j.issn.1000-4025.2016.05.1021
收稿日期:2015-10-12;修改稿收到日期:2016-01-17
基金项目:国家自然科学基金(31270255);中国科学院西部之光项目(2013-165);河北省自然科学基金(C2015205074)
作者简介:章博远(1992-),男,在读硕士研究生,主要从事苔藓植物分类学研究。E-mail: zhangboyuan9953@163.com *通信作者:王幼芳,教授,博士生导师,主要从事苔藓区系及分子系统学研究。E-mail: yfwang@bio.ecnu.edu.cn
中图分类号:Q949.35; Q948.5
文献标志码:A
