基于微卫星位点的中国4个野生东方田鼠群体的遗传多样性分析
2016-07-02倪丽菊孙凤萍肖君华周宇荀
高 骏,倪丽菊,孙凤萍,肖君华,周宇荀,李 凯*
(1东华大学化学化工与生物工程学院,上海201620;2上海市农业科学院畜牧兽医研究所,上海201106;3上海实验动物研究中心,上海201203)
基于微卫星位点的中国4个野生东方田鼠群体的遗传多样性分析
高 骏1,2,倪丽菊3,孙凤萍2,肖君华1,周宇荀1,李 凯1*
(1东华大学化学化工与生物工程学院,上海201620;2上海市农业科学院畜牧兽医研究所,上海201106;3上海实验动物研究中心,上海201203)
摘 要:利用20个微卫星位点,采用荧光-多重PCR分型技术,对4个野生东方田鼠群体(福建、湖南、宁夏、广西)进行遗传结构和遗传分化检测。结果表明:19个微卫星位点显示出高度多态性(PIC=0.83>0.5),平均等位基因数、位点杂合度均显示4个地区野生东方田鼠的遗传多样性丰富。F统计量表明:4个野生东方田鼠群体的遗传差异有13.5%来自于群体间,86.5%的遗传差异主要来源于个体间。遗传距离与UPGMA聚类分析均表明,宁夏群体与广西群体间的遗传分化程度最大,湖南群体与广西群体间的遗传分化程度最小,这在STRUCTURE聚类图分析时(K =2)同样得到提示。分析认为,广西群体与湖南群体间的遗传分化较小,存在一定基因交流的原因是这两个群体间较近的地理距离。
关键词:东方田鼠;微卫星位点;遗传多样性
东方田鼠(Microtus fortis)是我国一种重要的野生实验动物资源,主要分布在我国中东部低海拔地区,以及和我国接壤的俄罗斯、朝鲜、蒙古等地区。东方田鼠具有独特的、天然的抗血吸虫特性[1-2],是一种抗血吸虫病机理研究不可多得的动物材料。近年来的研究表明,东方田鼠还能作为糖尿病、卵巢癌、非酒精性脂肪肝等研究的模型动物[3-5]。20世纪90年代初期,国内的一些实验动物研究机构(中国科学院长沙农业现代化研究所、上海实验动物研究中心等)已将该物种引入实验室进行饲养、净化、繁育[6-7],目标是将其培育成具有中国特色的一种实验动物资源。目前,东方田鼠的亚种分类比较确定的是分布于湖南洞庭湖区的长江亚种(M.f.calamorum)和分布于宁夏地区的指名亚种(M.f.fortis),其他亚种的分类及地理分布区域一直存在较大的争议[8-12]。同时,对于中国东方田鼠各地理群体的分子生物学信息、遗传结构和多样性水平一直未见深入的研究报道。这些都阻碍了东方田鼠实验动物化进程的发展和资源的保护。
微卫星也称短串联重复(short tandem repeats,STR)或简单重复序列(simple sequence repeats,SSR),普遍分布于真核生物基因组中,广泛应用于物种群体遗传结构、亲缘关系分析及遗传资源评价等研究中[13-14]。本试验利用前期的研究方法[15-16]筛选了20个东方田鼠微卫星(STR)分子标记,并结合荧光多重PCR和分型方法和对我国4个地区野生东方田鼠群体的遗传多样性进行检测,旨在揭示该物种野生群体的遗传多样性现状,希望能够从分子地理学角度对该物种的亚种分类、系统进化研究提供研究基础,并为该物种的实验动物资源开发和保护提供帮助。
1 材料与方法
1.1样品采集
63个野生东方田鼠样本(4个地理种群)分别采自宁夏(NX)、湖南(HN)、广西(GX)、福建(FJ)(表1)。样本通过形态学和部分基因序列测序鉴定。

表1 东方田鼠样本采集信息Table 1 Sample collection information of M.fortis in China
1.2基因组总DNA的提取
取0.5—1 cm鼠尾,利用SDS/蛋白酶K裂解,酚氯仿提取总DNA,琼脂糖凝胶电泳检测提取的DNA质量。利用NanoDrop ND-1000浓度仪测定浓度,稀释至100 pmol/uL保存备用。
1.3东方田鼠微卫星标记及其引物
本试验选择前期利用磁珠富集法[15]获得的东方田鼠多态性良好的20个位点进行条件摸索和优化。引物5’端标记有HEX或FAM荧光(表2),引物均由上海生工生物有限公司合成。
1.4多重PCR扩增及产物检测
将20个STR基因座引物根据目的片段长度以及不同的荧光标记分成7组(Panel 1—7,表2)。通过多重PCR扩增试剂盒(TIANGEN BIOTECH,型号KT109)进行多重PCR扩增。PCR扩增总体积为12.5 μL:10×Multi HotStart Buffer 1.25 μL,Super Pure dNTPs(2.5 mmol/L)1 μL,Primer Mix 1 μL(各引物的终物质的量浓度为1 pmol/μL),DNA Template 2 μL(10 ng/μL),Multi HotStart DNA Polymerase 0.25 μL(5 U/μL),ddH2O 7 μL,另加10 μL石蜡油防止挥发。多重PCR扩增在Mastercycler gradient梯度PCR仪上(Eppendorf公司)完成,95℃预变性15 min;94℃变性30 s,54—59℃退火90 s,72℃延伸60 s,35个循环;72℃延伸10 min,4℃保存。反应结束后,将扩增产物经琼脂糖或聚丙烯酰胺凝胶电泳抽检后,在ABI3730自动测序仪上电泳分型,并利用GeneMapper v3.0软件对分型后的数据进行采集处理。
1.5数据统计及分析
将分型后获得的片段长度数据转化成个体的基因型数据,并利用CERVUS 3.0[17]和POPGENE v1.32软件[18]统计各位点的等位基因数、期望杂合度(He)、观测杂合度(Ho)、多态信息含量(PIC)、群体内和群体间的遗传分化系数、各位点的哈迪-温伯格平衡情况、Nei氏遗传距离以及基因流等。利用STRUCTURE v2.3.4[19]进行遗传结构的分析,在混合模型下,K值取2—5,并利用Distruct 1.1程序[20]构建遗传结构图。

表2 微卫星标记及其引物序列Table 2 Microsatellite markers and primers information
2 结果与分析
2.1群体遗传学参数检测
20个STR位点在4个东方田鼠野生群体63个样本中的平均等位基因数为8.523个,其中MFBAC52-29和MFB41位点的有效等位基因数(Ne)最少,分别为1.772和2.729个;而MFBAC52-05、MFBAC67-34、MFA56、MFA178、MFA257、MFA184这6个位点的有效等位基因数(Ne)均大于10,且PIC值相对最高(0.903—0.922),表明在20个STR位点中,该6个位点的遗传多样性最高。在20个位点中,只有MFBAC52-29位点的PIC值为0.370,处于0.25—0.5,属于中等多态性位点,其他19个位点的PIC值均大于0.5,表现为高度多态性位点(表3)。
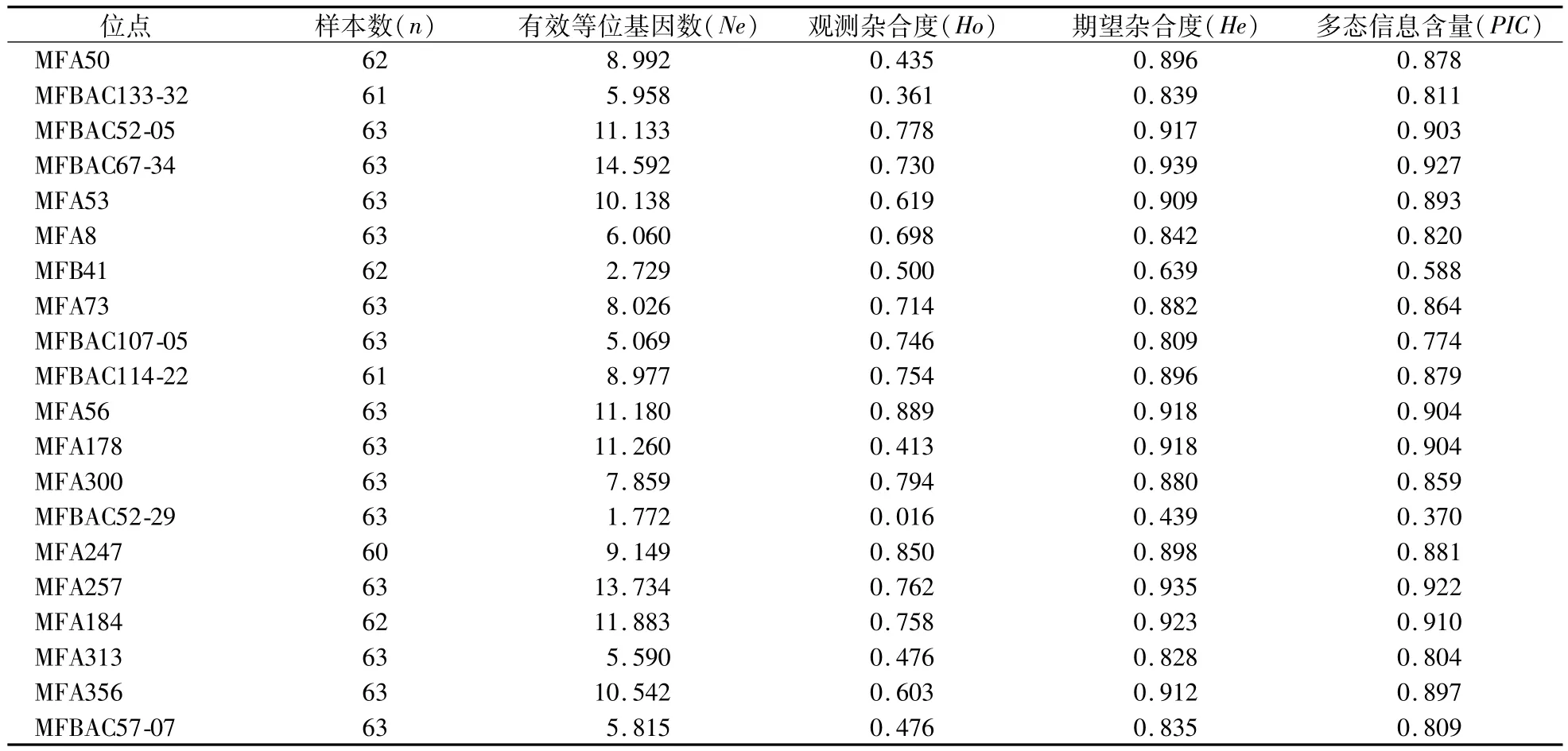
表3 20个微卫星(STR)座位的遗传多样性参数Table 3 Genetic diversity parameters of 20 microsatellite loci(STR)
2.2遗传分化与基因流
通过POPGENE v1.32检测东方田鼠各位点间的遗传分化系数[21]与基因流(表4),种群内的近交系数(Fis)值在-0.041—0.957,平均值为0.153。种群间的近交系数(Fit)值在0.023—0.964,平均值为0.267。其中MFBAC52-29位点的Fis和Fit值在所有20个位点中最高。种群间的遗传分化系数(Fst)值在0.042—0.366,平均值为0.135。各位点的基因流(Nm)的平均值为1.604,20个位点中仅MFA300、MFA313、MFBAC57-07这3个位点的基因流数值低于1;17个位点的Nm>1,表明各群体间还是保持了一定的基因流水平,较高的基因流水平能够抑制种群的遗传分化。
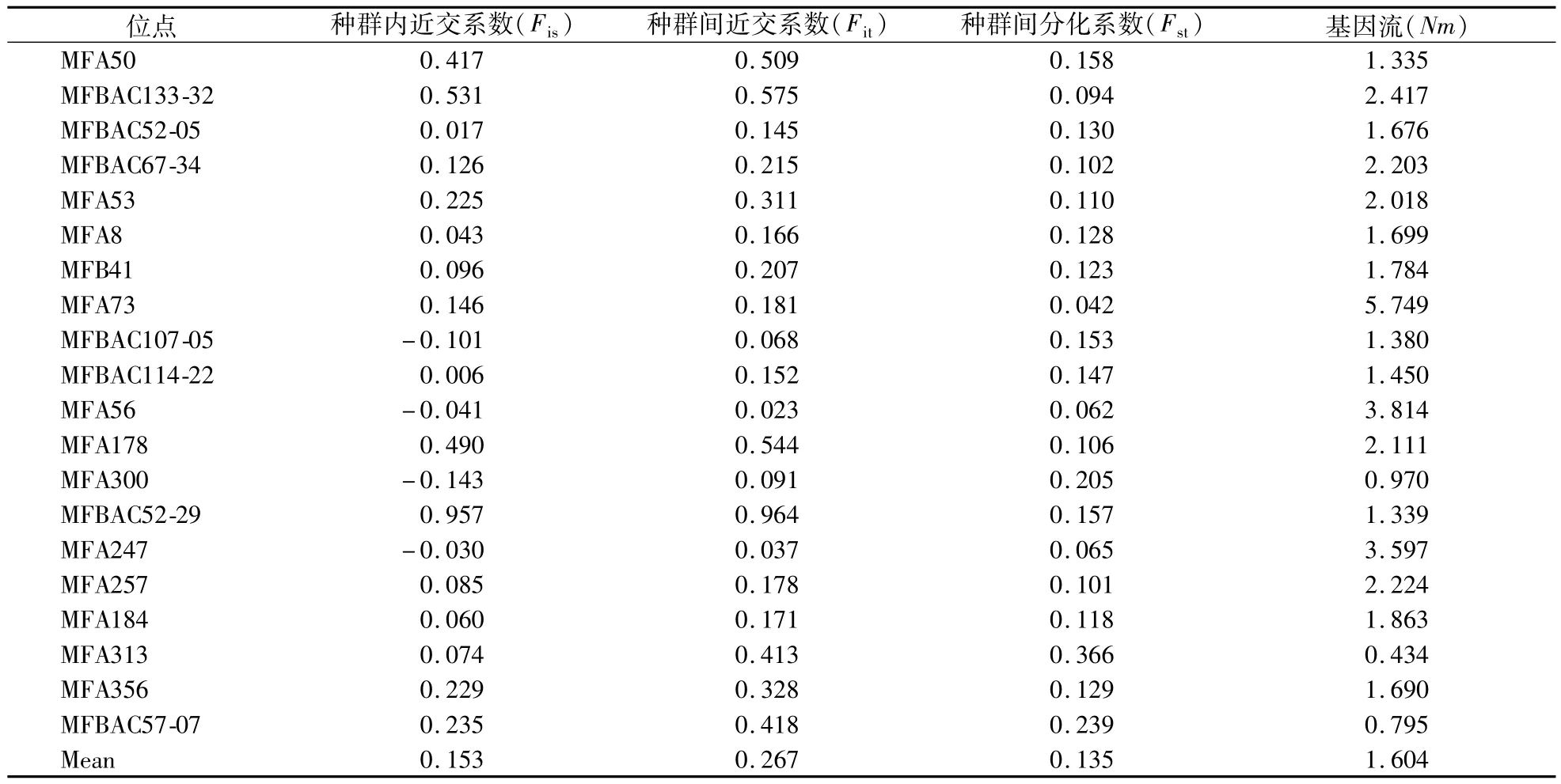
表4 东方田鼠各位点分化系数与基因流检测Table 4 Differentiation coefficient and gene flow detection of 20 Loci of M.fortis
2.3哈迪-温伯格平衡(Hardy-Weinberg equilibrium)检测
对4个群体20个位点进行哈迪-温伯格平衡检测(表5),发现有8个位点(MFBAC52-05、MFBAC67-34、MFA8、MFB41、MFBAC107-05、MFBAC114-22、MFA56、MFA300)在4个群体中均处于平衡状态(P>0.05),有2个位点(MFA178、MFBAC52-29)在4个群体中处于不平衡的状态(P<0.05)。从4个群体的总体看,有32.5%位点处于非平衡状态。
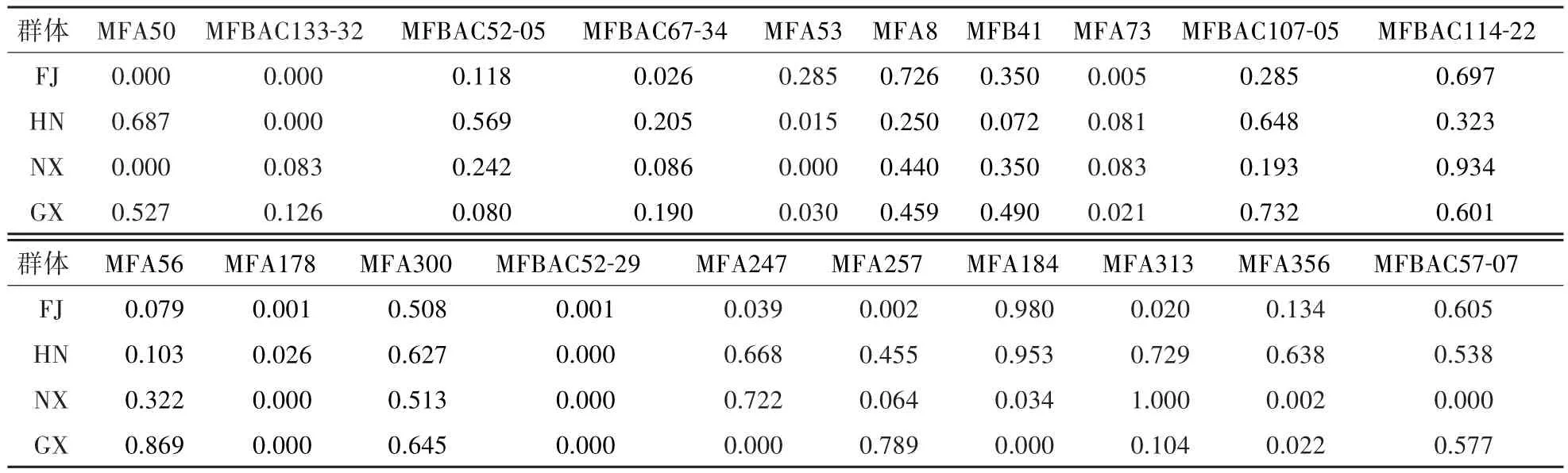
表5 20个位点的哈迪-温伯格平衡检测(P值)Table 5 P value of Hardy-Weinberg equilibrium test of 20 loci
2.4遗传距离与聚类分析
通过POPGENE v1.32统计4个群体东方田鼠的Nei氏遗传距离[22]与遗传一致性系数(表5)显示,宁夏群体(NX)与福建(FJ)、广西(GX)群体之间的遗传距离较远,分别达到了0.983和1.044;而湖南(HN)与广西(GX)、福建(FJ)群体之间的Nei氏遗传距离较近,分别为0.643和0.690(表6)。根据Nei氏无偏差遗传距离,构建了UPGMA聚类树(图1)。STRUCTURE程序推断的群体间的系统发生关系(图2)显示:K =2时,东方田鼠63个样本被分为2个类群,一类是包括宁夏(NX)群体的所有样本,另一类是包含福建(FJ)、湖南(HN)、广西(GX)群体的我国南方群体样本;当K =3时,FJ群体作为独立的群,被从南方群体中细分出来;当K =4时,广西与湖南群体被进一步得到细分;当K =5时,广西群体显示出复杂的基因组成,难以被清晰的划分。
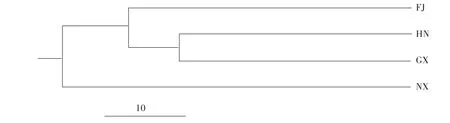
图1 根据Nei氏遗传距离构建的东方田鼠4个群体的UPGMA聚类树Fig.1 UPGMA cluster tree of 4 populations of M.fortis based on Nei’s genetic distances

表6 东方田鼠Nei氏遗传距离(下三角)与遗传一致性系数(上三角)Table 6 Nei’s genetic distance(below diagonal)and genetic consistency coefficient(above diagonal)of M.fortis
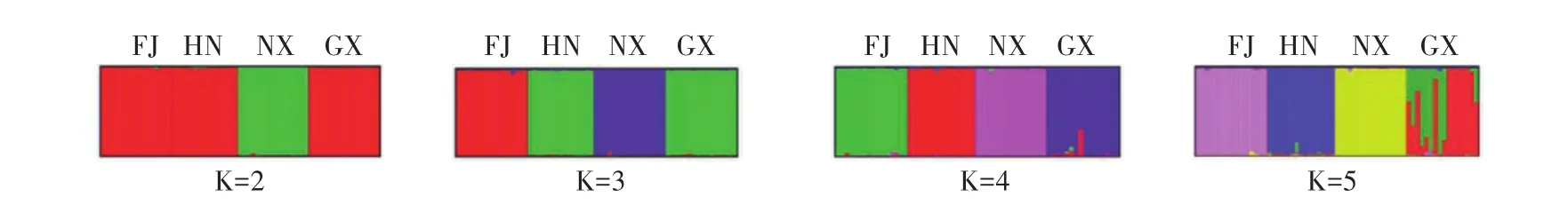
图2 STRUCTURE程序推断的东方田鼠4个群体(K =2—5)的群体遗传结构图Fig.2 The population genetic structure of 4 population of M.fortis inferred by STRUCTURE program(K from 2 to 5)
3 讨论
微卫星序列的分类包括:完美型、非完美型和复合型序列[23]。所谓完美型是指由单一的重复单元所组成的序列,如(AC)n;而非完美型是指重复序列中有其他核苷酸夹杂其中,复合型是由2个或多个重复单元组成的序列。此次选择的20个STR位点中,16个属于完美型,其余的为复合型。本研究选用的20 个STR位点在4个东方田鼠群体样本中表现出较高的遗传多态性,其中19个位点属于高度多态位点,平均多态信息量PIC =0.83>0.5,这些STR位点能够充分反映东方田鼠野生群体的遗传多样性信息。
F统计量的分析表明,从整体而言,4个野生东方田鼠群体存在明显的群体遗传分化,群体间的遗传分化系数为Fst=0.135,表明群体间的遗传差异有13.5%,而另外86.5%的遗传差异主要来源于个体间的差异。群体内的近交系数较高(Fit=0.153),可能是由于4个东方田鼠群体之间由于地理分布的局限性,导致群体间的基因交流受到一定程度的限制。哈迪-温伯格平衡(Hardy-Weinberg equilibrium)检测发现:在4个群体20个位点中,有32.5%的位点处于非平衡状态,表明东方田鼠野生群体其遗传结构并不稳定,可能受到选择、近交、群体规模偏小等因素的影响。其中有8个位点(MFBAC52-05、MFBAC67-34、MFA8、MFB41、MFBAC107-05、MFBAC114-22、MFA56、MFA300)在4个野生自然群体中均处于平衡状态,表明这8个位点可能更适合应用于东方田鼠种群资源的遗传检测。
分析4个东方田鼠野生群体的Nei氏遗传距离和基因流发现,宁夏群体(NX)与广西群体(GX)之间的遗传距离最远(1.044),而广西群体(GX)与湖南群体(HN)之间的遗传距离最近(0.643)。在4个东方田鼠野生群体中,宁夏群体(NX)在地理位置上属于我国北方的群体,而湖南、福建、广西属于我国南部地区的群体,从遗传距离和聚类分析都表明,宁夏群体(NX)与3个南方群体间的遗传分化程度较大。研究认为,湖南与广西在地理距离上比较接近,有助于这两个群体间的基因交流和个体的扩散。而宁夏属于中国北方地区,中国存在较多东西走向的山脉和河流,形成了天然的地理屏障(如秦岭-淮河地理分界线等),使得南北地区的物种种群产生明显的遗传分化。在STRUCTURE程序分析结果中也同样提示(K=2),该4个群体被分为两类,一类是以宁夏(NX)为代表的北方群体,而另3个群体(FJ、HN、GX)可以被划分成南方群体。虽然4个东方田鼠野生群体间的分化程度均较高,但STRUCTURE分析(K =4、5)也表明在广西群体与湖南群体之间存在一定的基因交流,分析认为这可能是因为两个群体间相对较近的地理距离,使得这两个种群在长期的进化过程中存在一定的扩散杂交。
我国是一个物种资源丰富的国家,许多独特的物种蕴含着重要的生物学特性。东方田鼠拥有天然的抗血吸虫感染的能力,使其医学研究价值被得到重视。但是对于在自然环境下的野生东方田鼠群体的分子遗传学研究较少,该物种的亚种分类一直存在较大争议、系统地理学研究也处于起步阶段。本研究所采用的20个STR位点在野生群体中表现出高度的遗传多态性,可以作为今后东方田鼠遗传检测、分子育种有效的遗传分子标记。本研究从基因组分子角度为今后中国东方田鼠亚种分类提供了一定的基因数据参考,为促进东方田鼠的实验动物资源化提供了重要的基础。
参 考 文 献
[1]贺宏斌,左家铮,刘柏香,等.室内繁殖和野生东方田鼠感染日本血吸虫的比较[J].实用寄生虫病杂志,1995,3(2):72-74.
[2]HE Y,LUO X,ZHANG X,et al.Immunological characteristics of natural resistance in Microtus fortis to infection with Schistosoma japonicum [J].Chin Med J(Engl),1999,112(7):649-654.
[3]俞远京,苏志杰,马亚东,等.诱发性东方田鼠糖尿病动物模型的初步研究[J].湖南畜牧兽医,2003(4):8-9.
[4]俞远京,苏志杰,周智君,等.东方田鼠自发上皮性卵巢癌的病理学研究[C]//中国实验动物学会第七届学术年会论文集.2006:210-213.
[5]杨玉琴,冯洁,柏熊,等.东方田鼠非酒精性脂肪肝模型的建立[J].中国实验动物学报,2013,21(2):34-38.
[6]俞远京,胡维新,王勇,等.野生东方田鼠实验动物化及种质资源保护的初步研究[J].实验动物科学与管理,2003,20(z1):13-20.
[7]柏熊,王晓东.东方田鼠净化的研究初报[J].上海畜牧兽医通讯,2006(6):33.
[8]黄文几,陈廷熹,温业新.中国啮齿类[M].上海:复旦大学出版社,1995:231-233.
[9]罗泽洵,陈卫,高武.中国动物志兽纲第六卷啮齿目(下册)仓鼠科[M].北京:科学出版社,2000:221-232.
[10]谭邦杰.哺乳动物分类名录[M].北京:中国医药科技出版社,1992:219-291.
[11]马勇.中国有害啮齿动物分布资料[J].中国农学通报,1986(6):76-82.
[12]洪震藩.东方田鼠的一新亚种:福建亚种[J].动物分类学报,1981,6(4):444-445.
[13]WEBER J L,MAY P E.Abundant class of human DNA polymorphisms which can be typed using the polymerase chain reaction[J].Am J Human Genet,1989,44(3):388-396.
[14]宋卫涛,李慧芳,韩威,等.我国地方蛋鸭品种遗传结构及遗传分化[J].四川农业大学学报,2010,28(1):93-98.
[15]倪丽菊,陶凌云,柏熊,等.东方田鼠微卫星标记的富集筛选与初步应用[J].遗传,2011,33(9):989-995.
[16]汤敏,倪丽菊,高骏,等.东方田鼠部分BAC文库的微卫星筛选[J].中国实验动物学报,2012,20(3):50-55.
[17]KALINOWSKI S T,TAPER M L,MARSHALL T C.Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment[J].Molecular Ecology,2007,16:1099-1106.
[18]YEH F C,YANG R C,BOYLE T.POPGENE version1.32,Microsoft window-base software for population genetic analysis:a quick user’s guide [M].University of Alberta,Center for International Forestry Research,Alberta,Canada,1999.
[19]PRITCHARD J K,STEPHENS M,DONNELLY P.Inference of population structure using multilocus genotype data[J].Genetics,2000,155:945.
[20]ROSENBERG N A.Distruct:a program for the graphical display of population structure[J].Molecular Ecology Notes,2004,4:137-138.
[21]WEIR B S,COCKERHAM C C.Estimation F-statistics for the analysis of population Structure[J].Evolution,1984,38:1358-1370.
[22]NEI M.Genetic distance between populations[J].Am Nat,1972,106:283-292.
[23]WEBER J L.Infornativeness of human(dC-dA)n·(dG-dT)n polymorphisms[J].Genomics,1990,7(4):524-530.
(责任编辑:闫其涛)
Genetic diversity analysis of 4 wild Microtus fortis populations based on microsatellite loci
GAO Jun1,2,NI Li-ju3,SUN Feng-ping2,XIAO Jun-hua1,ZHOU Yu-xun1,LI Kai1*
(1Institute of Biological Sciences and Technology,Donghua University,Shanghai 201620,China;2Institute of Animal Husbandry and Veterinary Sciences,Shanghai Academy of Agricultural Sciences,Shanghai 201106,China;3Shanghai Laboratory Animals Research Center,Shanghai,201203 China)
Abstract:Based on 20 microsatellite loci,the genetic structure and genetic differentiation of 4 wild Microtus fortis populations(Fujian,Hunan,Guangxi and Ningxia)were detected by multiplex fluorescence PCR technique.The results indicated that 19 microsatellite loci showed high polymorphism(PIC =0.83>0.5),and the average number of alleles and loci heterozygosity showed that the genetic diversity was abundant in 4 regions of M.fortis.F-statistic showed that the genetic difference among the populations was 13.5%,and 86.5%of genetic difference was mainly from individuals.Genetic distances and UPGMA cluster analysis showed that the degree of genetic differentiation between Ningxia population and Guangxi population was the largest,the degree of genetic differentiation between Hunan population and Guangxi population was the least,which was also prompted in STRUCTURE clustering graph analysis(K =2).The genetic differentiation between Guangxi population and Hunan population was small,the reason for the existence of a certain gene exchange is the closer geographical distance between the two populations.
Key words:Microtus fortis;Microsatellite loci;Genetic diversity
中图分类号:S865
文献标识码:A
文章编号:1000-3924(2016)03-072-06
DOI:10.15955/j.issn1000-3924.2016.03.15
收稿日期:2015-04-15
基金项目:上海市科委项目(09140900101、09140900102)
作者简介:高骏(1979—),男,博士,助理研究员,研究方向:动物分子遗传
*通信作者,E-mail:likai@dhu.edu.cn
