云南地方水稻品种抗稻瘟病基因Pita功能区段序列变异分析
2016-06-17马继琼杨奕孙一丁王炎炎许明辉
马继琼 杨奕 孙一丁 王炎炎 许明辉
(云南省农业科学院 生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室, 昆明 650223; #共同第一作者; *通讯联系人, E-mail: xuminhui@sohu.com)
云南地方水稻品种抗稻瘟病基因Pita功能区段序列变异分析
马继琼#杨奕#孙一丁王炎炎许明辉*
(云南省农业科学院 生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室, 昆明 650223;#共同第一作者;*通讯联系人, E-mail: xuminhui@sohu.com)
马继琼, 杨奕,孙一丁,等. 云南地方水稻品种抗稻瘟病基因Pita功能区段序列变异分析. 中国水稻科学, 2016, 30(3): 265-272.
摘要:稻瘟病抗性基因序列变异的分析与认识是挖掘水稻抗稻瘟病基因资源的基础。对来源于云南省56个县(市)的218份地方稻品种稻瘟病抗性基因Pita第二编码区功能位点区段的609个碱基进行了测序分析。结果表明,所有供试品种在此序列上有18个变异位点,存在46种DNA单倍型和28种蛋白类型,其中DNA单倍型Pita-H1、Pita-H2、 Pita-H6频率较高,三者比例分别为17.89%、29.82%、20.18%,合计67.89%,是云南的优势单倍型,而其他单倍型频率较低,频率0.46%~3.67%。携带Pita抗性基因(编码区第2752位功能位点碱基为G)的品种共61个,占供试材料的27.98%,其中39个品种(占63.93%)与越南抗性品种Tetep序列一致。Pita抗性基因在水稻各亚种和生态型中的分布是不平衡的,籼稻中的频率为35.06%,粳稻中为24.11%,水稻中31.03%,陆稻中21.92%,黏稻中30.77%,糯稻中18.37%。Pita抗性基因在云南省广泛分布,36县(市)检测到了Pita抗性基因,但南部地区频率高于其他地区,且有的县频率较高,呈小集中的特点,海拔2200m以上的7个品种未能检测到Pita抗性基因。
关键词:稻; 云南; 稻瘟病; Pita
稻瘟病是全世界各稻区最为严重的病害之一。由于稻瘟病菌的高度变异性和不稳定性,新育成的具有单一抗性基因的抗性品种在推广种植几年后就成为感病品种,这已成为水稻生产可持续发展的重要障碍,挖掘水稻抗性基因资源是抗稻瘟病育种的基础。
主效抗稻瘟病基因Pita是最早被克隆和研究最多的基因之一[1-4],在我国很多稻区表现高水平的稻瘟病抗性,被广泛应用于我国的水稻育种和生产[5]。地方品种Tadukan和越南品种Tetep的Pita基因被广泛应用于亚洲和美国的抗稻瘟病育种中[6,7]。李进斌等[8]的研究表明Pita抗性基因仍可在云南粳稻区利用。为了准确鉴定该基因,Jia等[9,10]已开发了引物YL155/YL87和YL183/YL87,并已广泛应用于资源的鉴定和育种中。杨杰等[11]利用Pita基因的功能标记,检测和分析了我国115份水稻地方品种的Pita基因型,结果表明Pita在我国浙江、福建、广东、广西、贵州、四川、安徽、江西、河南、河北、吉林均有分布。
Pita是单拷贝抗稻瘟病基因,包含2个外显子和1个1463 bp的内含子,编码一个NBS-LRR 蛋白,缺乏经典的富亮氨酸重复(LRR)区域,但含有一长度不等的不完全重复的氨基酸序列组成的富亮氨酸结构域(LRD)。在Pita蛋白LRD区域的单个氨基酸的差异,决定了等位基因属于抗病基因还是感病基因,即第918氨基酸是丙氨酸(A)则抗病,丝氨酸(S)则表现为感病反应[12]。Wang等[4]研究表明,除抗性位点外,Pita基因在栽培稻和野生稻中都存在大量的单核苷酸多态性位点和插入/缺失位点,存在15种Pita变异体和9种变异蛋白,而Thakur等[13]对印度地方稻种的分析表明存在64种单倍型和47种蛋白变异体。
云南地形气候复杂,海拔76-2700 m区域均有亚洲栽培稻地方品种分布,是世界上最大的稻种遗传多样性中心之一,基因多样性大于中国其他地理群[14]。赵国珍等[15]在云南景洪普通野生稻中发现存在Pita抗性基因,推测云南可能是抗稻瘟病基因Pita的起源地之一。李进斌等[16]曾用引物YL155/YL87和YL183/YL87对云南地方稻种进行分析,发现云南地方稻种携带Pita抗性基因的比例较高。
第二代测序技术的成熟与普及,为直接鉴定该位点DNA碱基提供了可能性,DNA测序同时可提供除功能位点外其他碱基变异的信息。本研究拟对云南地方稻种资源Pita基因包含功能位点的区段进行测序分析,明确Pita抗性基因的多样性,以及品种携带Pita抗性基因的情况,积累种质资源基因信息,为挖掘和利用基因资源提供指导。
1材料与方法
1.1材料
来源于云南省14个州市56个县(市)的218份地方稻品种作为试验材料。 按籼粳稻分类,籼稻77份,粳稻141份;按水陆稻分类,水稻175份,陆稻73份;按黏糯稻分类,黏稻169份,糯稻49份。材料均由云南省农业科学院生物技术与种质资源研究所保存。以日本晴作为测序对照品种,与从GenBank查询到的日本晴Pita基因序列AF207842进行比对,以检验测序的可靠性。
1.2DNA提取
采用CTAB法提取基因组DNA[17]。
1.3PCR扩增及测序
根据Jia等(私人通讯)提供的160余份水稻Pita基因序列,设计一对引物。扩增的是第二编码区后半段(包含功能位点)和终止子后的部分序列,大约1449 bp。前引物pita2-F位于基因编码区1344-1364位(第二编码区内),序列为TACTTTGCAGGGGATGAGGC,后引物pita2-R 位于基因终止子后20 bp,序列为TGATTCAGGGTCACAACAATAACA,由华大基因公司合成。
20 μL反应体系包括缓冲液2 μL,dNTPs 1 μL,前后引物各1 μL,ExTaq酶 0.4 μL,ddH2O 13.6 μL,DNA模板1 μL。反应程序如下:95℃下5 min;95℃下40 s,57℃下30 s,72℃下50 s,41个循环,72℃下10 min。
PCR扩增产物送至华大基因公司用后引物pita2-R进行测序,每个材料重复测序2次,若两次的测序结果不一致,则进行第3次测序,取其中2个相同的测序结果用软件BioLign进行比较分析,用软件MEGA 5.1进行系统树分析。
2结果与分析
2.1Pita基因功能区段变异与编码蛋白变异
引物pita2-F/pita2-R对所有供试材料基因组DNA进行扩增,均可扩增出一条大约1449 bp DNA片段,表明所有材料均携带Pita/pita基因片段。扩增片段用后引物pita2-R进行测序,每批次测序实验所获得的日本晴序列长度和碱基序列与从GenBank获得的AL772419.4序列的对应序列一致,说明测序资料可靠。测序实验一般可得到750 bp左右的正确序列,超出此范围测序准确性会受到影响。为了保证资料的准确性,我们只对所获得的序列中的609个碱基序列进行比较分析。通过比对,该序列位于Pita基因编码区2179-2787(终止子)位,包含了Pita基因功能位点2752位碱基和终止子。
表1云南水稻地方品种Pita基因功能序列变异位点与单倍型
Table 1. Sequence variant loci and haplotypes of Pita gene in rice landraces from Yunnan.
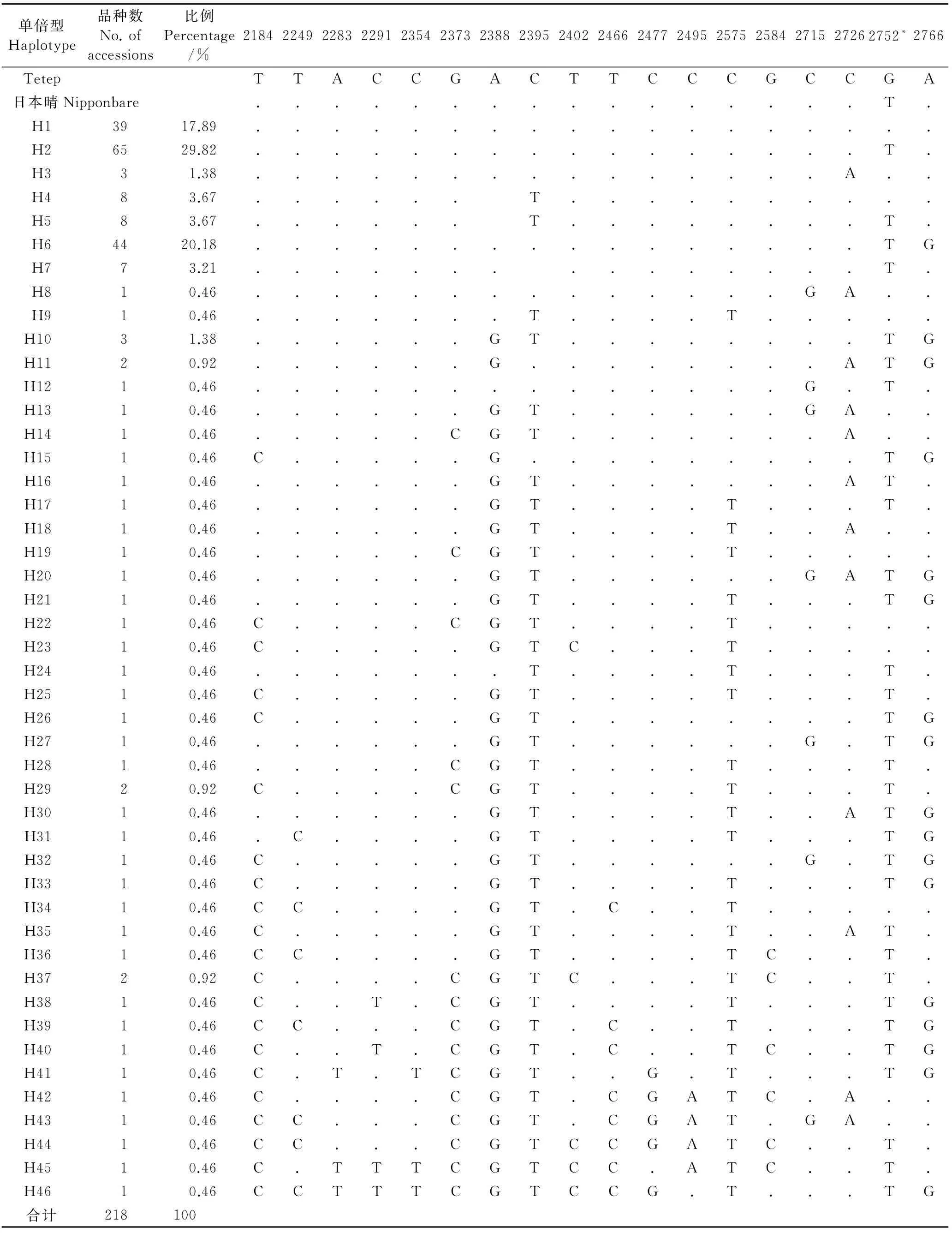
单倍型Haplotype品种数No.ofaccessions比例Percentage/%21842249228322912354237323882395240224662477249525752584271527262752*2766TetepTTACCGACTTCCCGCCGA日本晴Nipponbare................T.H13917.89..................H26529.82................T.H331.38...............A..H483.67......T..........H583.67......T........T.H64420.18................TGH773.21...............T.H810.46..............GA..H910.46.......T....T.....H1031.38......GT........TGH1120.92......G........ATGH1210.46..............G.T.H1310.46......GT......GA..H1410.46.....CGT.......A..H1510.46C.....G.........TGH1610.46......GT.......AT.H1710.46......GT....T...T.H1810.46......GT....T..A..H1910.46.....CGT....T.....H2010.46......GT......GATGH2110.46......GT....T...TGH2210.46C....CGT....T.....H2310.46C.....GTC...T.....H2410.46.......T....T...T.H2510.46C.....GT....T...T.H2610.46C.....GT........TGH2710.46......GT......G.TGH2810.46.....CGT....T...T.H2920.92C....CGT....T...T.H3010.46......GT....T..ATGH3110.46.C....GT....T...TGH3210.46C.....GT......G.TGH3310.46C.....GT....T...TGH3410.46CC....GT.C..T.....H3510.46C.....GT....T..AT.H3610.46CC....GT....TC..T.H3720.92C....CGTC...TC..T.H3810.46C..T.CGT....T...TGH3910.46CC...CGT.C..T...TGH4010.46C..T.CGT.C..TC..TGH4110.46C.T.TCGT..G.T...TGH4210.46C....CGT.CGATC.A..H4310.46CC...CGT.CGAT.GA..H4410.46CC...CGTCCGATC..T.H4510.46C.TTTCGTCC.ATC..T.H4610.46CCTTTCGTCCG.T...TG合计218100
“.”代表与第一行碱基相同; “*”代表功能位点。下同。
“.”, Indicates the same as the bases of the first line; “*” ,Indicates the functional sites. The same as in tables below.
与越南抗性品种Tetep的Pita基因序列进行比对分析,所有供试品种在609个碱基序列上存在18个变异位点,根据18个变异位点,此段序列存在46种单倍型(表1),其中日本晴归为Pita-H2,Tetep归为Pita-H1。各单倍型与抗性品种Tetep对应片段比较,每个单倍型存在0~13个位点的差异。各个单倍型出现频率存在极大的差异,其中Pita-H1、Pita-H2、Pita-H6频率较高,三者比例分别为17.89%、29.82%、20.18%,合计67.89%,是云南的优势单倍型,而其他单倍型频率较低,频率为0.46%~3.67%。综上所述,云南水稻地方品种在Pita基因功能位点序列上存在丰富的变异,单倍型多,其中优势单倍型为3种,而多数单倍型频率较低。
表2云南水稻地方品种Pita基因编码蛋白序列变异位点与蛋白质类型
Table 2. Sequence variant loci and type of Pita gene encoded proteins in rice landraces from Yunnan.
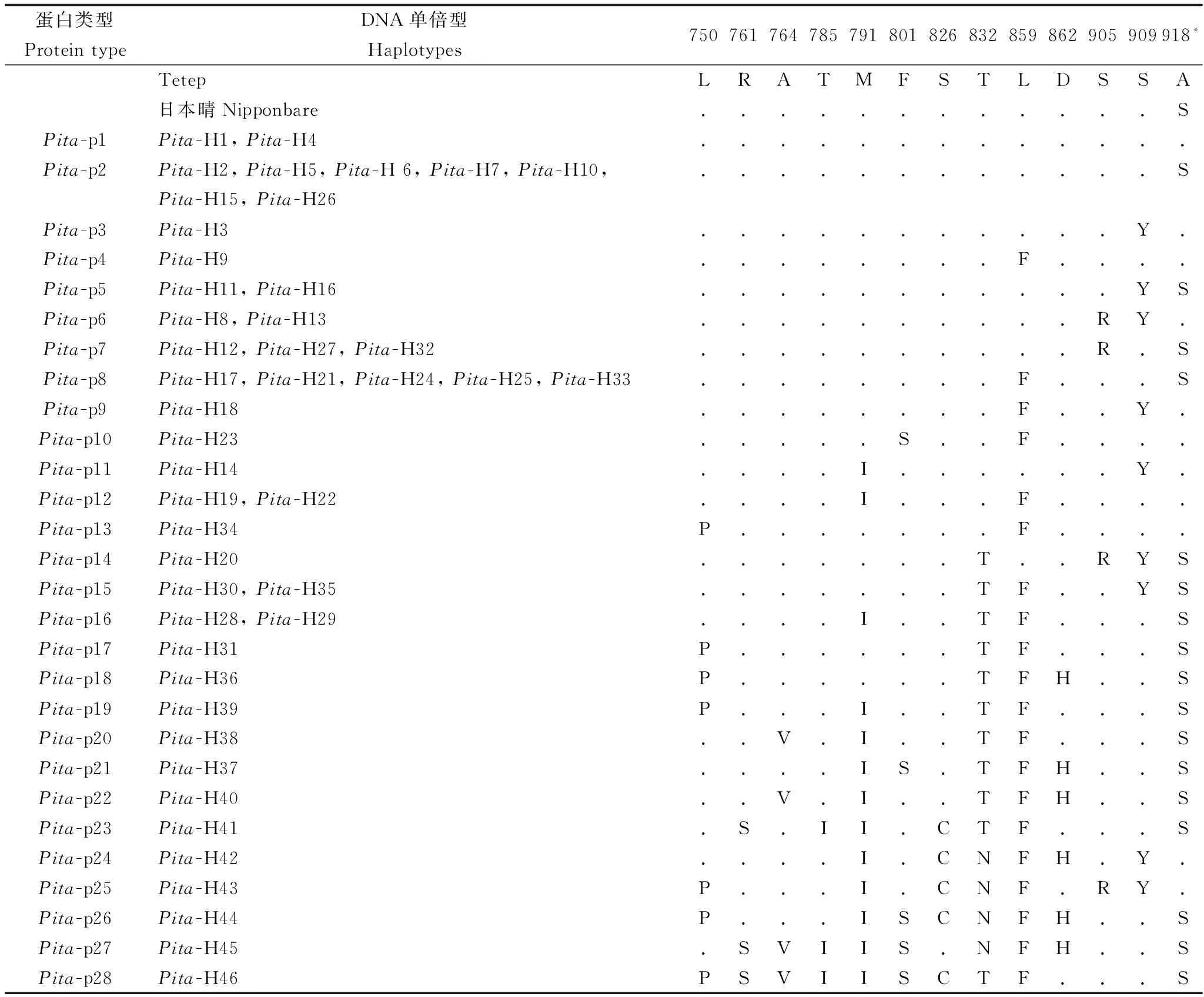
蛋白类型ProteintypeDNA单倍型Haplotypes750761764785791801826832859862905909918*TetepLRATMFSTLDSSA日本晴Nipponbare............SPita-p1Pita-H1,Pita-H4.............Pita-p2Pita-H2,Pita-H5,Pita-H6,Pita-H7,Pita-H10,Pita-H15,Pita-H26............SPita-p3Pita-H3...........Y.Pita-p4Pita-H9........F....Pita-p5Pita-H11,Pita-H16...........YSPita-p6Pita-H8,Pita-H13..........RY.Pita-p7Pita-H12,Pita-H27,Pita-H32..........R.SPita-p8Pita-H17,Pita-H21,Pita-H24,Pita-H25,Pita-H33........F...SPita-p9Pita-H18........F..Y.Pita-p10Pita-H23.....S..F....Pita-p11Pita-H14....I......Y.Pita-p12Pita-H19,Pita-H22....I...F....Pita-p13Pita-H34P.......F....Pita-p14Pita-H20.......T..RYSPita-p15Pita-H30,Pita-H35.......TF..YSPita-p16Pita-H28,Pita-H29....I..TF...SPita-p17Pita-H31P......TF...SPita-p18Pita-H36P......TFH..SPita-p19Pita-H39P...I..TF...SPita-p20Pita-H38..V.I..TF...SPita-p21Pita-H37....IS.TFH..SPita-p22Pita-H40..V.I..TFH..SPita-p23Pita-H41.S.II.CTF...SPita-p24Pita-H42....I.CNFH.Y.Pita-p25Pita-H43P...I.CNF.RY.Pita-p26Pita-H44P...ISCNFH..SPita-p27Pita-H45.SVIIS.NFH..SPita-p28Pita-H46PSVIISCTF...S
“.”代表与第一行氨基酸相同; “*”功能位点。
“.” Indicates the same as the amino acid of the first line; “*” Indicates the functional locus.
表3Pita抗性基因在云南地方水稻品种各亚种和生态类型中的分布
Table 3. Distribution of Pita resistance genes in different subspecies and ecotypes.

群体类型Grouptype材料数No.ofaccessions携带G样品数No.ofvarietiescarryingGlocus频率Frequency/%总体Totalpopulation2186127.98籼稻indica772735.06粳稻japonica1413424.11水稻Rice1454531.03陆稻Uplandrice731621.92黏稻Non-glutinousrice1695230.77糯稻Glutinousrice49918.37
根据前人结果,2752位出现碱基G的为抗性基因,出现T的为感病基因[12]。在46种单倍型中,Pita-H1、Pita-H3、Pita-H4、Pita-H8、Pita-H9、Pita-H13、Pita-H14、Pita-H18、Pita-H19、Pita-H22、Pita-H23、Pita-H34、Pita-H42、Pita-H43等14种为抗性单倍型,在2752位为G,携带抗性基因,合计61个品种,占27.98%,比中国其他地区高[11],但比我们用分子标记YL155/YL87和YL183/YL87检测的低[16]。值得注意的是,61个携带抗性基因的品种中有39个(占63.93%)与越南抗性品种Tetep序列一致,表明它们可能是同源的,而与越南抗性品种Tetep序列不一致的品种可能是云南特有或Tetep型的变异类型。
根据此段序列,理论上可翻译成202个氨基酸,所有供试材料间存在13个氨基酸位点的差异,与越南抗性品种Tetep对比,存在0~10个氨基酸的差异,可归为28种蛋白质类型,类型较丰富(表2)。
为了了解各蛋白类型间的关系,根据202个氨基酸序列进行了系统进化分析。利用软件MEGA(版本5.1)中的Maxinum-Likelihood, Mininum-Evolution, Neighbor-Joining等3种分析方法得到的系统树完全一致,本研究给出Neighbor-Joining法Pita基因系统进化树(图1)。由图1可以看出,云南水稻地方品种可分两大枝,第一大枝包含了不携带Pita抗性基因的单倍型Pita-p16、Pita-p17、Pita-p18、Pita-p19、Pita-p20、Pita-p21、Pita-p22、Pita-p23、Pita-p26、Pita-p27、Pita-p28,品种数量少;其余单倍型归为第二枝,包含了云南大多数品种,是云南的主体,有携带抗性基因的,也有不抗的。第二枝中抗感品种混合在一起,这意味着Pita抗性基因可能来源于野生稻,也可能来源于感病品种的突变,即二次起源。在第二枝中,抗性单倍型Pita-p4、Pita-p10、Pita-p12、Pita-p13比Tetep更接近树基,说明Tetep型(云南的多数抗性品种)起源可能更晚些。
2.2Pita抗性基因在水稻亚种和生态型中的分布
功能位点2752位携带G的云南地方稻品种在籼粳亚种间、水陆稻间、黏糯稻间的分布存在明显的差异(表3),籼稻中的频率为35.06%,粳稻中为24.11%,水稻中31.03%,陆稻中21.92%,黏稻中30.77%,糯稻中18.37%。说明Pita抗性基因在水稻各亚种和生态型中的分布是不平衡的。
2.3Pita抗性基因在云南的地理分布
218个材料来源14个州市58县市(表4),其中13个州市36县(市)检测到了Pita抗性基因,而西双版纳州、红河州、文山州等南部边缘地区频率高于其他地区。在检测到Pita抗性基因的县(市)中,有的县频率较高,呈小集中的特点。如永胜县、盐津县、广南县、墨江县、西盟县等较为集中,特别是保山市的各县都检测到了抗性基因。可见,Pita抗性基因在云南呈现广泛分布,小集中的特点。在云南,海拔是稻种类型分布最重要的影响因子,Pita抗性基因在不同海拔地区的分布见表5。海拔1200-2200 m地区Pita抗性基因分布频率相对较高。值得一提的是,海拔2200 m上有7个品种,却没有1个品种携带抗性基因,可能是材料少造成的,也可能是这一区域的品种不携带抗性基因,需要加大材料数目研究后才可得知。
3讨论
Pita抗性基因标记YL155/YL87和YL183/YL87设计在内含子中,利用了分子标记与功能位点的连锁关系,鉴定一般需2个PCR。Wang等[18]用该Pita-InDel分子标记筛选来自全球的1790份水稻种质资源,其中183份携带类抗病基因,经测序证实148份材料功能位点为G,携带抗性基因,而29份携带T,为感病基因,鉴定准确率为80.87%,存在一定的误差。李进斌等[16]也曾用Pita引物YL155/YL87和YL183/YL87对云南稻种资源Pita基因进行鉴定,携带抗病基因材料高达40.9%,而本研究对功能位点测序结果表明抗病基因材料占27.98%,用Pita-InDel分子标记筛选存在较大误差。Wang等[18]对163份功能位点为G的材料进行接种鉴定,发现有4份尽管携带抗病基因但表现感病,可能由抗病信号转导元件的突变所致,准确率为97.55%。可见,利用测序方法直接鉴定功能位点,结果直观准确,在测序技术成熟与普及的今天不失为Pita基因鉴定首选方法。
表4Pita抗性基因在云南不同地区和县(市)的分布
Table 4. Distribution of Pita resistance genes in Yunnan different regions.
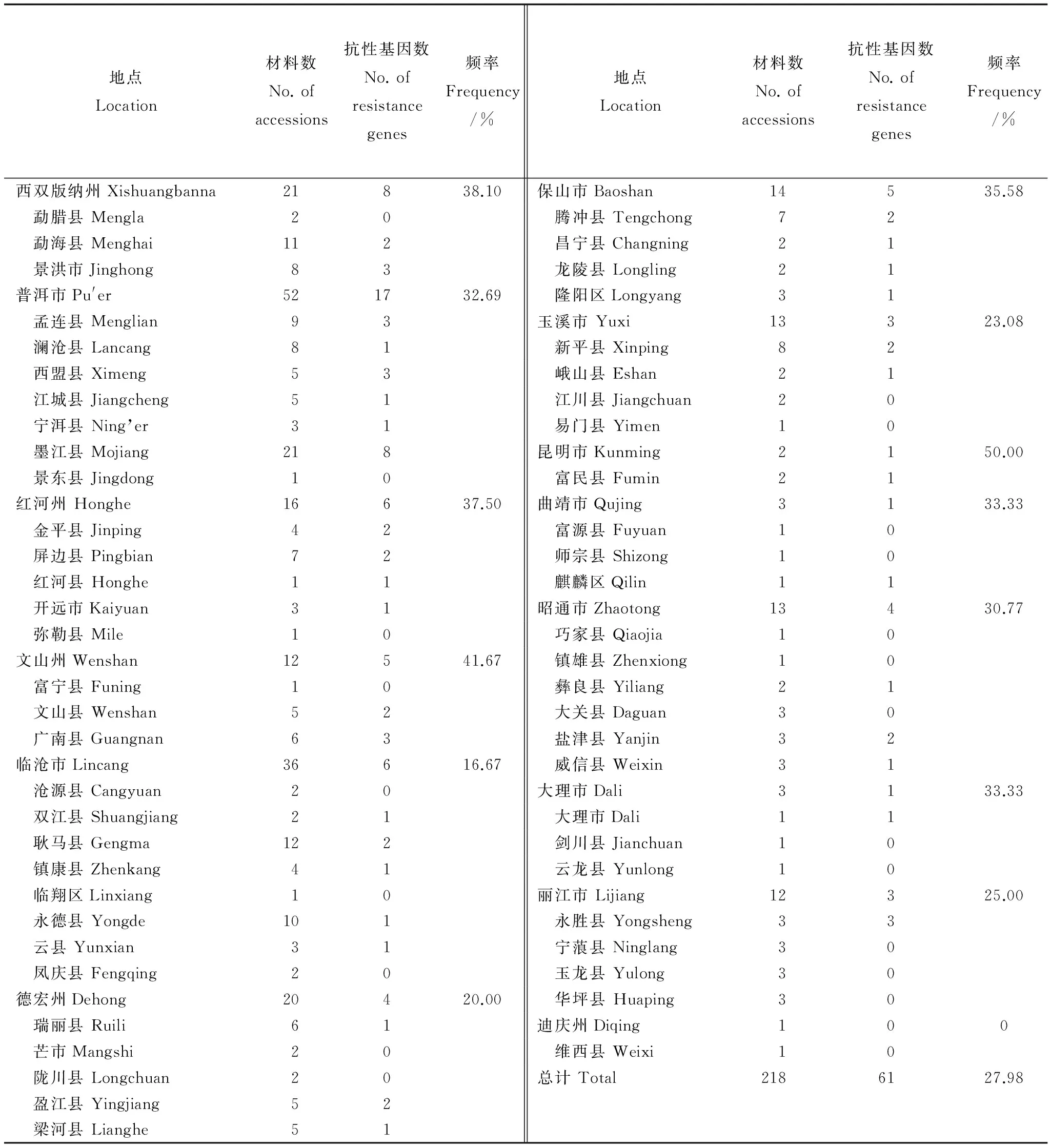
地点Location材料数No.ofaccessions抗性基因数No.ofresistancegenes频率Frequency/%地点Location材料数No.ofaccessions抗性基因数No.ofresistancegenes频率Frequency/%西双版纳州Xishuangbanna21838.10保山市Baoshan14535.58 勐腊县Mengla20 腾冲县Tengchong72 勐海县Menghai112 昌宁县Changning21 景洪市Jinghong83 龙陵县Longling21普洱市Pu'er521732.69 隆阳区Longyang31 孟连县Menglian93玉溪市Yuxi13323.08 澜沧县Lancang81 新平县Xinping82 西盟县Ximeng53 峨山县Eshan21 江城县Jiangcheng51 江川县Jiangchuan20 宁洱县Ning’er31 易门县Yimen10 墨江县Mojiang218昆明市Kunming2150.00 景东县Jingdong10 富民县Fumin21红河州Honghe16637.50曲靖市Qujing3133.33 金平县Jinping42 富源县Fuyuan10 屏边县Pingbian72 师宗县Shizong10 红河县Honghe11 麒麟区Qilin11 开远市Kaiyuan31昭通市Zhaotong13430.77 弥勒县Mile10 巧家县Qiaojia10文山州Wenshan12541.67 镇雄县Zhenxiong10 富宁县Funing10 彝良县Yiliang21 文山县Wenshan52 大关县Daguan30 广南县Guangnan63 盐津县Yanjin32临沧市Lincang36616.67 威信县Weixin31 沧源县Cangyuan20大理市Dali3133.33 双江县Shuangjiang21 大理市Dali11 耿马县Gengma122 剑川县Jianchuan10 镇康县Zhenkang41 云龙县Yunlong10 临翔区Linxiang10丽江市Lijiang12325.00 永德县Yongde101 永胜县Yongsheng33 云县Yunxian31 宁蒗县Ninglang30 凤庆县Fengqing20 玉龙县Yulong30德宏州Dehong20420.00 华坪县Huaping30 瑞丽县Ruili61迪庆州Diqing100 芒市Mangshi20 维西县Weixi10 陇川县Longchuan20总计Total2186127.98 盈江县Yingjiang52 梁河县Lianghe51
目前的研究表明普通野生稻中有携带Pita抗

*抗性单倍型。
* Indicates the resistant haplotypes.
图1云南地方水稻品种Pita基因功能片段编码的29种蛋白单倍型系统进化关系
Fig. 1. Phylogenetic tree constructed with the 29 protein haplotypes encoded by Pita genes from Yunnan rice landraces.
表5Pita抗性基因在云南不同海拔地区的分布
Table 5. Distribution of Pita resistance genes at different elevation in Yunnan.

海拔Elevation/m材料数No.ofaccessions抗性基因数No.ofresistancegenes比例Percentage/%<80020630.0801-1200852023.51201-1600732331.51601-2000281035.72001-22005360.0>2200700.0
性基因的材料[15,19],普通野生稻是栽培稻Pita抗性基因的来源之一,栽培稻与野生稻进行基因交流可能是重要的原因之一。云南南部的景洪普通野生稻携带Pita抗性基因,巧合的是西双版纳州及周边的热带地区红河州、文山州Pita抗性基因频率明显高于其他地区,可能是栽培稻与野生稻进行基因交流造成的。据此推测云南东南部以南地区(越南、老挝等)稻种中可能也存在较高频率的Pita抗性基因,这需要进一步的分析才可得知。
R基因的进化是对病原强加到植物上的选择压力所做出的回应[20]。云南稻瘟病菌群体遗传多样性水平较高[21,22],云南水稻地方品种在Pita基因上存在丰富的变异类型,这可能与云南稻瘟病菌复杂相关。
参考文献:
[1]Huang E, Hwang S, Chiang Y, Lin T. Molecular evolution of thePi-tagene resistant to rice blast in wild rice (Oryzarufipogon).Genetics, 2008, 179: 1527-1538.
[2]Jia Y, Martin R. Identification of a new locusPtr(t) required for rice blast resistance genePi-ta-mediated resistance.MolPlantMicrobeInteract, 2008, 21: 396-403.
[3]Lee S, Costanzo S, Jia Y, Olsen K M, et al. Evolutionary dynamics of the genomic region around the blast resistance genePi-tain AA genomeOryzaspecies.Genetics, 2009, 183: 1315-1325.
[4]Wang X, Jia Y, Shu Q Y, et al. Haplotype diversity at thePi-talocus in cultivated rice and its wild relatives.Phytopathology, 2008, 98: 1305-1311.
[5]时克, 雷财林, 程治军, 等. 稻瘟病抗性基因Pita和Pib在我国水稻主栽品种中的分布. 植物遗传资源学报, 2009, 10(1): 21-26.
Shi K, Lei C L, Cheng Z J, et al. Distribution of two blast resistance genesPitaandPibin major rice cultivars in China.JPlantGeneticResour, 2009, 10(1): 21-26.(in Chinese with English abstract)
[6]Moldenhauer K A K, Lee F N, Gibbons J W, et al. Registration of ‘Ahrent’ rice.CropSci, 2007, 47: 446-447.
[7]Rybka K, Miyamoto M, Ando I, et al. High resolution mapping ofindica-derived rice blast resistance genesPi-ta2 andpi-taand a consideration of their origin.MolPlantMicrobe-Interact, 1997, 10: 517-524.
[8]李进斌, 李成云, 陈艳, 等. 二十二个抗稻瘟病基因在云南的利用价值评价. 植物保护学报, 2005, 32(2): 113-119.
Lin J B, Li C Y, Chen Y, et al. Evaluation of twenty-two blast resistance genes inYunnan using monogenetic rice lines.ActaPhytophylSin, 2005, 32(2): 113-119. (in Chinese with English abstract)
[9]Jia Y, Wang Z, Singh P. Development of dominant rice blastPi-taresistance gene markers.CropSci, 2002, 42: 2145-2149.
[10]Jia Y, Wang Z, Robert G. RicePi-tagene confers resistance to the major pathotypes of the rice blast fungus in the United States.Phytopathology, 2004, 94: 296-301.
[11]杨杰, 杨金欢, 王军, 等. 稻瘟病抗病基因Pita和Pib在中国水稻地方品种中的分布. 华北农学报, 2011, 26(3): 1-6.
Yang J, Yang J H, Wang J, et al. Distribution of two blast resistant genesPitaandPibin landrace rice in China.ActaAgricBoreali-Sin, 2011, 26(3): 1-6.(in Chinese with English abstract)
[12]Bryan G T, Wu K S, Farrall L, et al. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance genePi-ta.PlantCell, 2000, 12(11): 2033-2046.
[13]Thakur S, Gupta Y K, Singh P K, et al. Molecular diversity in rice blast resistance genePi-tamakes it highly effective against dynamic population ofMagnaportheoryzae.FunctIntegrGenom, 2013, 13: 309-322.
[14]黄燕红,孙新立,王象坤. 中国栽培稻遗传多样性中心和起源研究. 植物遗传资源学报, 2005, 6(2): 125-129.
Huang Y H, Sun X L, Wang X K. Study on the center of genetic diversity and its origin of cultivated rice in China.JPlantGeneticResour, 2005, 6(2): 125-129.(in Chinese with English abstract)
[15]赵国珍, 蒋春苗, 刘吉新, 等. 云南野生稻抗稻瘟病Pi-ta基因的检测及分析. 中国水稻科学, 2014, 28(6): 675-680.
Zhao G Z, Jiang C M, Liu J X, et al. Identification and analysis on the rice blast resistance genePi-tain wild rice from Yunnan, China.ChinJRiceSci, 2014, 28(6): 675-680.(in Chinese with English abstract)
[16]李进斌, 王甜, 许明辉. 云南地方稻种抗稻瘟病基因Pi-ta和Pib的鉴定. 中国水稻科学, 2012, 26(5): 593-599.
Li J B, Wang T, Xu M H. Identification ofPitaandPi-bgenes for rice blast resistance of rice landraces from Yunnan Province.ChinJRiceSci, 2012, 26(5): 593-599.(in Chinese with English abstract)
[17]Doyle J J, Doyle J L. Isolation of plant DNA from fresh tissue.Focus, 1990, 12(1): 13-15.
[18]Wang X, FjeIlsrom R, Jia Y, et al. Characterization ofPitablast resistance gene in an international rice core collection.PlantBreeding, 2010(129): 491-501.
[19] Huang C L, Hwang S Y, Chiang Y C, et al. Molecular evolution of thePi-tagene resistant to rice blast in wild rice (Oryzarufipogon).Genetics, 2008, 179(3): 1527-1538.
[20]Rausher M D. Co-evolution and plant resistance to natural enemies.Nature, 2001, 41: 857-864.
[21]Li J B, Yang J, Liu L, He H M, He X H, Zhu Y Y, Li C Y. Geographic distribution of avirulence genes in rice blast fungus in Yunnan Province, China.JResourEcol, 2011, 2(2): 181-185.
[22]李进斌, 罗朝喜, 李成云, 等. 云南省近年稻瘟病菌生理小种的组成和分布. 植物保护, 2001, 27(2): 12-14.
Li J B, Luo C X, Li C Y, et al. Race composition and distribution ofMagnaporthegriseaCavara in Yunnan Province in recent years.PlantProt, 27(2): 12-14.(in Chinese with English abstract)
Sequence Variation of Functional Fragment inPitaGene for Rice Blast Resistance in Yunnan Rice Landraces
MA Ji-qiong#, YANG Yi#, SUN Yi-ding, WANG Yan-yan, XU Ming-hui*
(InstituteofBiotechnologyandGeneticResources,YAAS,KeyLaboratoryofBiotechnologyResearchofYunnanProvince/KeyLaboratoryofSouthwesternCropGeneResourcesandGermplasmInnovationofMinistryofAgriculture,Kunming650223,China;#These authors contributed equally to this work;*Corresponding author, E-mail: xuminhui@sohu.com)
MA Jiqiong, YANG Yi, SUN Yiding, et al. Sequence variation of functional fragment inPitagene for rice blast resistance in Yunnan rice landraces. Chin J Rice Sci, 2016, 30(3): 265-272.
Abstract:Identification and analysis of the variations of rice blast resistance gene sequence is the basis to dig into the genetic resources of the rice blast resistance. In this paper, 609 bases in the second code functional region of the rice blast resistance gene Pita were analyzed using 218 rice landraces from 56 counties of Yunnan Province. A total of 18 nucleotide variations were found in the sequence and the ORFs of 218 rice accessions were grouped into 46 haplotypes and 28 proteins types. Pita-H1 (17.89%), Pita-H2(29.82%) and Pita-H6(20.18%) were high-frequency types in Yunnan, while other types were at low frequency (0.46%-3.67%). So the advantage haplotypes in Yunnan were Pita-H1, Pita-H2 and Pita-H6 (totally 67.89%). A total of 61 rice landraces carrying the resistance gene Pita, the base functional site 2752 was G with the rate of 27.98%. The sequence of Pita in 39 resistance varieties was identical with the sequence obtained from the Vietnamese resistance variety Tetep. The distribution of resistance gene Pita in rice subspecies or ecotypes was unbalanced. The distribution frequency of indica was 35.06%, japonica 24.11%, rice 31.03%, upland rice 21.92%, no-glutinous rice 30.77%, glutinous rice 18.37%. The distribution of blast resistance gene Pita in Yunnan Province featured wide distribution and small concentration. Pita gene was detected in 36 counties of Yunnan Province. The distribution frequency in southern Yunnan was higher than other area. Moreover, some counties were of high distribution frequency. The resistance gene Pita was not detected in seven rice varieties growing over 2200 m elevation.
Key words:rice; Yunnan; blast; Pita
DOI:10.16819/j.1001-7216.2016.5188
收稿日期:2015-12-22; 修改稿收到日期: 2016-01-08。
基金项目:国家基础研究计划资助项目(2012FY110200); 国家自然科学基金资助项目(31360331)。
中图分类号:S435.111.4+1;S511.034
文献标识码:A
文章编号:1001-7216(2016)03-0265-08
中国水稻科学(Chin J Rice Sci),2016,30(3):265-272
http://www.ricesci.cn
