大翼厚皮橙叶片及种子形态变异研究
2016-06-04牟凤娟李一果李双智杨艳双刘天娥西南林业大学林学院云南昆明6504昆明市生物资源开发创新办公室云南昆明650500
牟凤娟李一果李双智杨艳双刘天娥(.西南林业大学林学院,云南昆明6504;.昆明市生物资源开发创新办公室,云南昆明650500)
大翼厚皮橙叶片及种子形态变异研究
牟凤娟1李一果2李双智1杨艳双1刘天娥1
(1.西南林业大学林学院,云南昆明650224;2.昆明市生物资源开发创新办公室,云南昆明650500)
摘要:对西双版纳分布的野生大翼厚皮橙3个居群进行调查和取样,采用方差分析、相关分析等方法对其叶片、种子的15个形态特征在群体内和群体间的变异进行分析。结果表明:同一居群内各个表型的变异程度不同;9个表型在群体间均存在极显著差异,1个表型差异为显著,其余4个表型为差异不显著;种子形态表现较叶片更为稳定。15个形态特征的变异系数分别为,翼叶宽度29.64%,翼叶长度22.96%,本叶宽度22.46%,本叶长度20.97%,翼叶长/本叶长20.73%,叶总长20.09%,本叶长/翼叶长17.47%,翼叶宽/本叶宽16.97%,种子长/宽14.97%,翼叶长/宽13.39%,种子厚度12.34%,本叶长/宽12.04%,种子宽度11.37%,翼叶长/叶总长10.39%,种子长度10.10%。11个叶片及4个种子形态特征间多数表现为极显著正相关或负相关。
关键词:大翼厚皮橙;叶片;种子;形态;变异
大翼厚皮橙(Citrus macroptera var.kerrii)属芸香科(Rutaceae)柑桔属大翼橙亚属(Subgen.Papeda),是柑橘属中比较原始的种类。该种为拉尼西亚大翼橙(C.macroptera)的1个变种[1],具有较原变种更大的果实及更多心室的特点。主要分布于印度东北部,泰国中北部,越南北部,以及中国云南南部等地区[1-6]。目前,国内仅在云南西双版纳局部地区的湿性季节性雨林中发现有少量大翼厚皮橙野生植株分布[3],成年植株周围幼苗较少,种群更新能力极差。由于大翼厚皮橙的叶片及果实形状与柚(C.maxima)极为相似,在当地被称为“野泡果”,误为“野柚子”。大翼厚皮橙与柚除在叶形、果实形状上有一定差异外,花和种子的差异也较为明显,前者为小花型,花冠直径仅1.0~1.5 cm,种子较小;后者为大花型,花冠直径2.5~4.0 cm,且种子较大[3]。
目前,对美拉尼西亚大翼橙及其变种的研究报道主要集中在化学成分[7-9]及其生物活性,包括抗氧化[10-11]、抗菌[12]、抗寄生[13]及果实毒性等[14]方面,其他零星报道还涉及珠心细胞培养植株再生[15]、用途和社会经济价值[4,16],以及局部地区(印度东北部Tripura和Mizoram)分布居群的分子遗传多样性[17],印度东北部Mizoram和Meghalaya地区分布居群的叶片、果实及种子形态特征表现[6]。虽然陈进等[3]曾对西双版纳地区大翼厚皮橙的分布、植物学性状、花粉形态、果实营养成分、果皮精油含量及种子的萌发特点等进行了报道,但目前对国内分布居群的遗传多样性及形态特征变异方面的研究未见报道。在此拟通过研究大翼厚皮橙国内分布居群的表型变异,了解其种群的遗传多样性,以期为保护和利用该野生柑橘种质资源提供理论依据。
1 材料与方法
1.1试验材料
2015年1月对西双版纳分布的野生大翼厚皮橙3个居群进行调查和取样。由于每个居群内植株数量极少,其中有果实的植株更少,随机采集每个居群中所有植株的成熟叶片、果实等试验材料。其中景洪1(JH-1)、景洪2(JH-2)和勐腊(ML)采集叶片分别为4、3、8株,采集果实分别为1、2株和1株。每个居群至少采集1号凭证标本,保存于西南林业大学标本馆(SWFC)。
1.2研究方法
1.2.1取样方法
1)叶片取样。从每个植株上、中、下不同部位采集的叶片混匀后采用随机抽样的方法,取至少10片成熟新鲜叶备用。
2)种子取样。由于每个居群内结果植株极少,取样时大部分果实已成熟落地,部分果实又被动物啃食,难以区分其植株来源。为此,采集每个居群内所有果实,处理种子,所有种子混匀后随机取样。
1.2.2测定方法
1)叶片形态特征测定。大翼厚皮橙叶片为单身复叶,且总叶柄两侧的翅较为发达,即具有发达的翼叶(图1)。对每个居群中每个植株的叶片随机抽样并编号;测量叶片的叶总长、翼叶长、翼叶宽、本叶长、本叶宽等各项叶片形态指标。

图1 大翼厚皮橙叶片及种子形态特征Fig.1 The leaf and seed characters of Citrrus macroptera var.kerrii
2)种子形态特征测定。种子清洗后于室内自然干燥。每个居群采用随机抽样方法抽取30粒种子,编号;用精度为0.1 mm的游标卡尺测量每粒种子的种子长度、宽度及厚度(图1)。
1.2.3数据分析
用Excel 2007和SPSS 19.0等软件对大翼厚皮橙叶片及种子的多项形态指标进行差异分析、单因素方差分析F检验和相关分析。
2 结果与分析
2.1形态特征变异
2.1.1居群内形态变异分析
从表1的变异系数可知,同一居群内各个形态性状的变异程度不同,如景洪1居群翼叶长/本叶长的变异系数最大(25.28%),翼叶宽/本叶宽的变异系数最小(9.65%);景洪2居群中变异系数最大的是翼叶宽度(28.30%),种子长度的变异系数最小(7.51%);勐腊居群中翼叶宽度的变异系数最大(23.87%),而种子宽度的变异系数最小(7.85%)。
2.1.2居群间形态变异分析
从表1各表型平均值可知,居群间翼叶长/本叶长、本叶长/宽、种子长度、种子长/宽等4个表型特征平均值为:景洪1居群>勐腊居群>景洪2居群,而叶总长、翼叶长度、翼叶宽度、翼叶长/宽等4个表型特征平均值为:景洪2居群>勐腊居群>景洪1居群。本叶长度、本叶宽度、种子厚度等3个表型特征平均值为:景洪2居群>景洪1>勐腊居群;翼叶长/本叶长、本叶长/翼叶长2个表型特征平均值为:勐腊居群>景洪1居群>景洪2居群,翼叶宽/本叶宽表型特征平均值为:勐腊居群>景洪2居群>景洪1居群。种子宽度的表型特征平均值为:景洪1居群>景洪2居群>勐腊居群。
同一形态性状的变异系数在不同居群中存在一定程度的变化,如叶总长的变异系数为:景洪居群2>景洪居群1=勐腊居群,本叶长度、本叶宽度、种子厚度的变异系数是景洪居群2>景洪居群1>勐腊居群,而翼叶长度、翼叶宽度的变异系数是景洪居群2>勐腊居群>景洪居群1;种子长度的变异系数是景洪居群1>勐腊居群>景洪居群2,种子宽度的变异系数是景洪居群1>景洪居群2>勐腊居群。不同群体内每一性状的变异系数均有一定差异,说明不同居群的环境异质性导致了居群表型变异的差异。
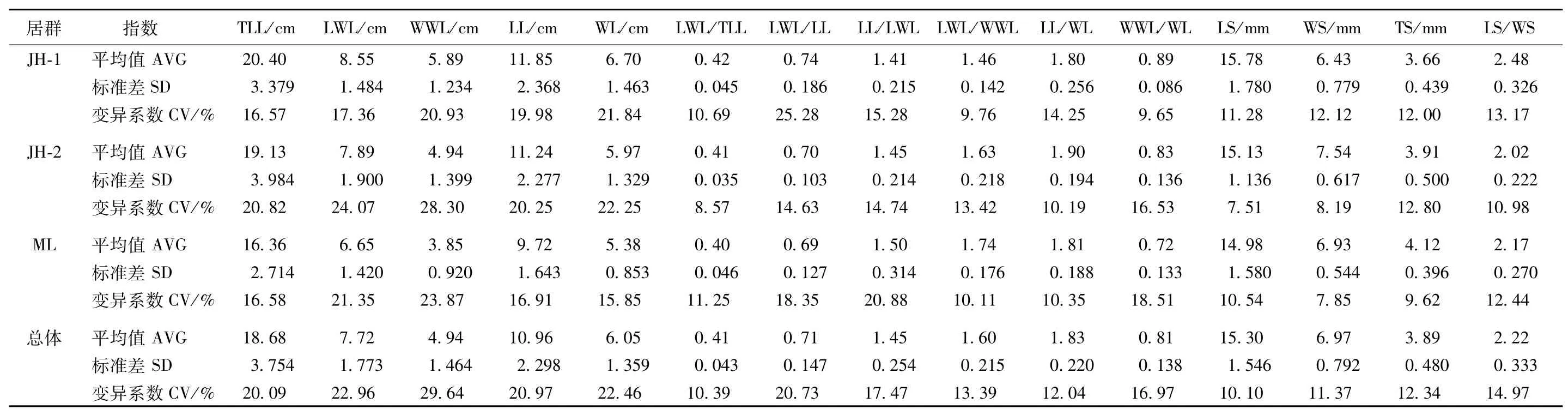
表1 大翼厚皮橙叶片、种子变异分析Table1 Thevarianceanalysisofleafand seed ofCitrusmacroptera var.kerrii
居群间形态变异单因素方差分析F检验结果表明(表2),在调查的15个表型中,叶总长、翼叶长度、翼叶宽度、本叶宽度、翼叶长/宽、翼叶宽/本叶宽、种子宽度、种子厚度、种子长/宽等9个性状在居群间的差异达极显著水平(P<0.01),本叶长度在居群间的差异为显著水平,其他4个性状在居群间的差异不显著。比较F值发现,翼叶宽度在居群间的差异最大(F = 29.008),翼叶长/本叶长在居群间的差异最小(F = 1.031)。检验结果说明,翼叶宽度在3个居群间的变异最大,而翼叶与本叶的长度比值在3个居群间的变异最小,表现最稳定。
2.1.3种内形态变异分析
大翼厚皮橙种内叶片、种子形态变异较大(表1)。叶片形态的变异系数为10.39%~29.64%,变异系数最大的是翼叶宽度(29.64%),其次依次为翼叶长度(22.96%)、本叶宽度(22.46%)、本叶长度(20.97%)、翼叶长/本叶长(20.73%)、叶总长(20.09%),而本叶长/翼叶长(17.47%)、翼叶宽/本叶宽(16.97%)、翼叶长/宽(13.39%)、本叶长/宽(12.04%)、翼叶长/叶总长(10.39%)等的变异相对较小。种子形态的变异系数为10.10%~14.97%,种子长度(10.10%)、种子宽度(11.37%)、种子厚度(12.34%)和种子长/宽(14.97%)的变异程度依次增大。翼叶宽度在种内的变异最大,而翼叶长/叶总长比值的变异最小,说明翼叶长/叶总长这个叶片性状的表现最稳定,可以作为种类鉴定的可靠指标之一。
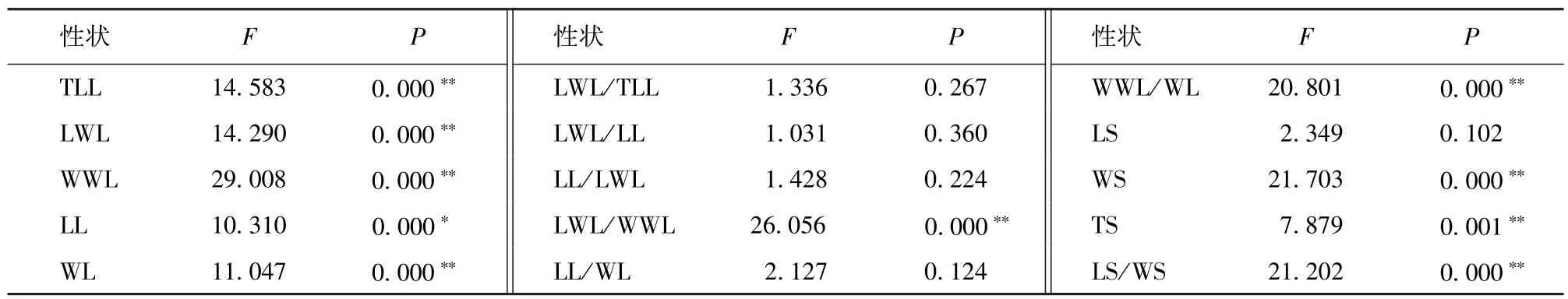
表2 单因素方差分析F检验Table 2 F-test of one-factor analysis of variance
2.2形态特征相关性
2.2.1叶片各形态特征相关性
相关分析发现,许多形态特征间表现为极显著相关(表3),其中21对特征为极显著正相关,如叶总长与翼叶长度、翼叶宽度、本叶长度、本叶宽度、翼叶宽/本叶宽(R分别为0.898、0.850、0.941、0.858、0.372),说明叶片越长,此5个特征值也越大;仅有本叶长度与本叶长/翼叶长、本叶长/宽表现为显著正相关(R均为0.191)。另外,17对叶片形态特征表现为极显著负相关,如翼叶长/宽与叶总长、翼叶长度、翼叶宽度、本叶长度、本叶宽度(R分别为-0.364、-0.311、-0.657、-0.356、-0.506);仅有本叶长度与叶长/叶总长比值为显著负相关(R=-0.239)。

表3 大翼厚皮橙叶片特征相关分析Table 3 The correlation analysis on leaves traits of Citrus macroptera var.kerrii
2.2.2种子各形态特征相关性
相关性分析发现,大翼厚皮橙的种子长度、种子宽度及种子厚度彼此间相关性不显著;而种子长/宽比值与种子长度为极显著正相关(R=0.630),与种子宽度为极显著负相关(R=-0.719)(表4)。
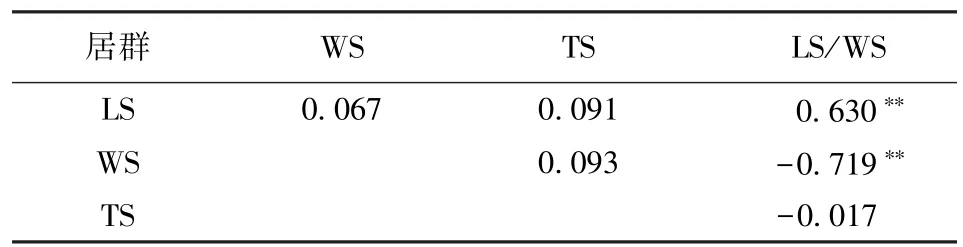
表4 大翼厚皮橙种子特征相关分析Table 4 The correlation analysis on seeds traits of Citrus macroptera var.kerrii
3 结论与讨论
3.1叶片、种子形态特征多样性
物种的遗传变异可体现在居群、个体、组织细胞及分子等不同水平[18],同一物种的不同个体或居群间存在各种形式及程度不同的形态变异,这些变异往往是各种生物、非生物因子所导致的自然选择在强度和方向上差异的综合产物,即物种的表型多样性是遗传多样性与环境多样性的综合体现[19],使形态变异往往具有一定生态适应意义。因而,从形态学或表型性状上估计物种的遗传变异是一种有效的方法[20-22]。
大翼橙类柑橘为单身复叶,且翼叶较为发达。翼叶的绝对大小(包括长度、宽度)、形状(翼叶长/宽比值)及其相对大小(即翼叶长/叶总长、翼叶长/本叶长比值)表现对于大翼橙种类的鉴定具有极其重要的意义。调查分析发现,大翼厚皮橙的翼叶宽度在居群间具有较大的变异,而翼叶长/总叶长比值和翼叶长/本叶长比值这两个特征的变异较小,但在Malik等的研究中仅探讨了本叶的相关表现[6],没有涉及翼叶这个比较重要的性状。在本研究中,大翼厚皮橙的叶片、种子性状的多个表现在物种水平有较为丰富的变异,CV值变动于10.10%~29.64%。每个表型在不同居群的变异程度明显不同,且营养性状(叶片)的变异程度普遍高于繁殖性状(种子)。与其他研究相比,本研究发现,西双版纳地区分布的大翼厚皮橙本叶长度、本叶宽度、本叶长/宽3个性状的平均值分别为10.96 cm、6.05 cm、1.83,范围分别为6.80~18.00 cm、3.30~10.90 cm、0.70~2.57,变异系数分别为20.97%、22.46%、12.04%,与印度东北部分布的(平均值分别为8.97 cm、5.12 cm、1.75,范围分别为7.2~11.5 cm、3.9~7.3 cm、1.60~2.06,变异系数分别为22.5%、16.72%、18.45%)相比[6],本地区的本叶长度、本叶宽度及本叶长/宽的平均值均更大、范围也更大;种子长度、种子宽度2个性状的平均值分别为15.30、6.97 mm,范围分别为11.16~18.14、4.88~8.82 mm,变异系数分别为10.10%、11.37%,与印度东北部分布的(平均值分别为14.41、6.40 mm,范围分别为12~17、4~8 mm,变异系数分别为12.61%、17.81%)相比[6],西双版纳地区的种子长宽值均更大,但变异更小,主要原因可能是这3个居群间的地理距离较近。
分布在生物居群间的变异可真实反映物种在不同生境中的适应状况,其大小在某种程度上说明了该物种对不同生态环境适应的广泛程度,即值越大,其适应的环境就越广[23-24]。Malik等利用RAPD分子标记研究发现,分布于印度东北部的拉尼西亚大翼橙具有较高的遗传多样性,且Mizoram居群较Tripura居群具有更高的多样性[17],本研究也发现,大翼厚皮橙同一形态特征的变异系数在不同居群中存在一定程度的变化,即居群间的变异程度不同。虽然大翼厚皮橙群体间距离较小,但许多表型在群体间的变异依然明显大于群体内,说明该物种对环境的适应能力较大。导致该物种个体数量极少的原因还需从其他方面予以探讨,如果实成熟落地后种子易被小动物啃食,成年植株周围极少有幼苗,种群更新困难,这也可能是导致其数量较少的重要原因之一。
3.2形态特征相关性
一种植物的形态特征外在表现是长期适应进化的结果,各个性状之间往往存在一定联系,一个性状的变异可能导致其他相关性状的相应变异。本研究发现,大翼厚皮橙叶片各性状之间,以及种子性状之间均存在较高的相关性,这说明各性状在共同维持叶片、种子结构的稳定性方面具有协调一致的适应性。
[参考文献]
[1]Swingle W T.The botany of Citrus and its relatives of the orange subfamily[M]/ / Webber H J,Batchelor L D.The Citrus Industry.Vol.1.Berkeley and Los Angeles:University of California Press,1943:129-474.
[2]Swingle W T,Reece P C.The botany of Citrus and its wild relatives(Family Rutaceae,Subfamily Aurantioideae)[M]/ / Reuther W,Webber H,Batchelor L.The Citrus Industry,Vol.I.History,world distribution,botany and varieties.Berkeley:University of California Press,1967:190-430.
[3]陈进,苏应春,陈贵清,等.云南西双版纳大翼厚皮橙的研究[J].中国柑桔,1993,22(3):3-5.
[4]黄成就.芸香科[M]/ /《中国植物志》编辑委员会.中国植物志,第43卷第2分册.北京:科学出版社,1997:175-184.
[5]Sharma B D,Hore D K,Gupta S G.Genetic resources of Citrus of north-eastern India and their potential use [J].Genetic Resources and Crop Evolution,2004,51 (4):411-418.
[6]Malik S K,Chaudhury R,Dhariwal O P,et al.Collection and characterization of Citrus indica Tanaka and C.macroptera Montr.:wild endangered species of northeastern India[J].Genetic Resources and Crop Evolution,2006,53(7):1485-1493.
[7]Dreyer D L,Huey P F.Coumarins of Citrus macroptera [J].Phytochemistry,1973,12(12):3011-3013.
[8]Gaillard E,Muyard F,Bevalot F,et al.Rutacea from New Caledonia.Chemical composition of stem bark of Citrus macroptera Montr.[J].Annales Pharmaceutiques Francaises,1995,53(2):75-78.
[9]Miah M N,Bachar S C,Nahar L.Composition of the volatiles of Citrus macroptera var.annamensis and evaluation of bioactivity[J].Journal of Essential Oil Bearing Plant,2010,13(2):211-218.
[10]Chowdhury S A,Sohrab M H,Datta B K,et al.Chemical and antioxidant studies of Citrus macroptera[J].Bangladesh Journal of Scientific and Industrial Research,2008,43(4):449-454.
[11]Rahman H,Eswaraiah M C,Dutta A M.In-vitro anti-oxidant activity of Citrus macroptera(var.annamensis)fruit peels extracts[J].International Journal of Pharmacy and Pharmaceutical Sciences,2014,6(1):364-371.
[12]Waikedre J,Dugay A,Barrachina I,et al.Chemical composition and antimicrobial activity of the essential oils from New Caledonian Citrus macroptera and Citrus hystrix[J].Chemistry&Biodiversity,2010,7(4):871-877.
[13]Desrivot J,Waikedre J,Cabalion P.Antiparasitic activity of some New Caledonian medicinal plants[J].Journal of Ethnopharmacology,2007,112(1):7-12.
[14]Uddin N,Hasan M R,Hasan M M,et al.Assessment of toxic effects of the methanol extract of Citrus macroptera Montr.fruit via biochemical and hematological evaluation in female sprague-dawley rats[J].PLoS One,2014,9(11):1-7.
[15]Miah M N,Islam S,Hadiuzzaman S.Regeneration of plantlets through somatic embryogenesis from nucellus tissues of Citrus macroptera Mont.var.anammensis(‘Sat Kara’)[J].Plant Tissue Culture,2002,12(2):167-172.
[16]Malik S K,Chaudhury R.The cryopreservation of embryonic axes of two wild and endangered Citrus species [J].Plant Genetic Resources:Characterization and U-tilization,2006,4(3):204-209.
[17]Malik S K,Uchoib A,Kumara S,et al.Molecular characterization of Citrus macroptera Montr.(Satkara):An endangered wild species from northeast India[J].Plant Biosystems,2013,147(4):857-863.
[18]Moritz C,Hillis D M.Molecular systematics context and controversies[M]/ / Hillis D M,Moritz C,Mable B K.Molecular Systematics.Sunderland:Sinauer Associates,1996:1-13.
[19]顾万春.统计遗传学[M].北京:科学出版社,2004.
[20]Schaal B A,Leverich W J,Rogstad S H.Comparison of methods for assessing genetic variation in plant conservation biology[M]/ / Falk D A,Holsinger K E.Genetics and Conservation of Rare Plants.New York:Oxford University Press,1991:123-134.
[21]葛颂,洪德元.遗传多样性及其检测方法[M]/ /钱迎倩,马克平.生物多样性研究的原理与方法.北京:中国科学技术出版社,1994:123-140.
[22]葛颂.遗传多样性[M]/ /蒋志刚,马克平,韩兴国.保护生物学.杭州:浙江科学技术出版社,1997:11-19.
[23]李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[24]李梅,韩海荣,康峰峰,等.山西灵空山辽东栎种群叶性状表型变异研究[J].北京林业大学学报,2005,27(5):10-16.
(责任编辑赵粉侠)
第1作者:牟凤娟(1977—),女,博士,副教授。研究方向:植物分类及植物地理。Email:moufengjuan@126.com。
Studies on Morphological Variation of Leaf and Seed of Citrus macroptera var.kerrii
Mou Fengjuan1,Li Yiguo2,Li Shuangzhi1,Yang Yanshuang1,Liu Tiane1
(1.College of Forestry,Southwest Forestry University,Kunming Yunnan 650224,China;2.Biological Resources Development Innovation Office of Kunming,Kunming Yunnan 650500,China)
Abstract:Investigation and sampling of three populations of Citrus macroptera var.kerrii distributed in Xishuangbanna.The variance analysis,the correlation analysis and other methods were used to discuss the variation of 15 morphological characters of leaf and seed in the population and among the populations of C.macroptera var.kerrii.The results showed that the variance degree of all traits in the same population were different,the difference of 9 traits among populations was very significant,that of one trait was significant,and that of other 4 traits were not significant,the morphology of seeds was more stable than that of leaves.the coefficients of variance of 15 traits were respectively as follows:the WWL(width of wing leaf)29.64%,the LWL(length of wing leaf)22.96%,the WL (width of leaflet)22.46%,the LL(length of leaflet)20.97%,the LWL/ LL 20.73%,the TLL(total length of leaf)20.09%,the LL/ LWL 17.47%,the WWL/ WL 16.97%,the LS/ WS(length of seed/ width of seed)14.97%,the LWL/ WWL 13.39%,the TS(thickness of seed)12.34%,the LL/ WL 12.04%,the WS 11.37%,the LWL/ TLL 10.39%,the LS 10.10%.Most correlation efficient of 11 leaf traits and 4 seed traits were obvious positive or negative.
Key words:Citrus macroptera var.kerrii,leaf,seed,morphology,variation
基金项目:国家自然科学基金地区基金项目(31360048)资助;西南林业大学云南省省级重点学科(林学)资助;西南地区生物多样性保育国家林业局重点实验室开放基金资助。
收稿日期:2015-11-25
doi:10.11929/ j.issn.2095-1914.2016.03.010
中图分类号:S722.3
文献标志码:A
文章编号:2095-1914(2016)03-0054-06
