干旱胁迫对云南松苗木生理特征的影响
2016-06-04琪蔡年辉诗王大玮段安安许玉兰西南林业大学云南省高校林木遗传改良与繁育重点实验室云南昆明6504西南林业大学西南山地森林保育与利用省部共建教育部重点实验室云南昆明6504
孙 琪蔡年辉,陈 诗王大玮段安安许玉兰(.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南昆明6504;.西南林业大学西南山地森林保育与利用省部共建教育部重点实验室,云南昆明6504)
干旱胁迫对云南松苗木生理特征的影响
孙 琪1蔡年辉1,2陈 诗2王大玮1段安安1许玉兰1
(1.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南昆明650224;2.西南林业大学西南山地森林保育与利用省部共建教育部重点实验室,云南昆明650224)
摘要:对云南松苗木进行干旱胁迫处理,探讨不同干旱胁迫天数对云南松苗木渗透调节物质与保护酶活性的影响。结果表明:在一定范围内,随着干旱胁迫天数的增加,云南松苗木质膜相对电导率、丙二醛、脯氨酸、可溶性糖含量及其超氧化物歧化酶活性均有不同程度的上升,但变化幅度因干旱胁迫程度、渗透调节物质类型的不同而有差异。不同干旱胁迫天数处理间的可溶性糖含量和超氧化物歧化酶活性差异极显著,丙二醛含量差异显著,脯氨酸与质膜相对电导率含量差异不显著。渗透物质调节能力表现为可溶性糖>脯氨酸,云南松苗木抗旱性相关性次序为丙二醛>质膜相对电导率,云南松苗木脯氨酸与质膜相对电导率变化对干旱胁迫处理响应较弱。在干旱胁迫下云南松苗木膜伤害与渗透调节、酶保护同步进行,具有一定的耐旱性。
关键词:云南松;干旱胁迫;渗透调节;酶活性;耐旱性
云南松(Pinus yunnanensis)为云南省造林的主要树种,因为其适应性强,耐干旱、耐贫瘠的特点,在云南有广泛的分布面积[1]。云南松是良好的建筑用材树种,工业用材树种和采脂树种,造纸性能也很好,其根、叶、树脂、花粉与幼果皆可入药,具有良好的经济价值[2],因此,对云南松进行研究十分必要。近些年来,西南地区干旱频发,甚至发生了几十年不遇的大旱与秋冬连旱[3],给农林与林业的生产生活造成了严重的经济损失,大面积的云南松林因旱灾而出现个体或成片死亡的情况[4-5]。水分是影响植物生长发育的重要环境因素之一[6-8],旱灾造成的损失是自然灾害造成损失中较为严重的[9]。云南松作为云南省重要的乡土树种之一,在云南林业经济和生态建设中占有举足轻重的作用[10]。但由于旱灾或干旱胁迫的影响,云南松林分表现出死亡现象。因此,如何充分利用有限的水资源进行森林植被的恢复和重建,成为改善生态环境建设中亟待解决的问题。所以,开展云南松抗旱性研究尤为重要。目前,对云南松的抗旱研究大部分是利用PEG(高分子渗透调节剂聚乙二醛)模拟干旱胁迫对种子萌发及幼苗生长的影响[11-12],而对云南松苗木在不同干旱胁迫程度下苗木生理特征变化的研究相对较少。鉴于此,本试验通过控制干旱胁迫天数开展云南松苗木渗透调节物质与保护酶活性变化研究,以期找出各项生理指标在不同干旱胁迫条件下的变化,探究云南松苗木的抗旱性,为云南松苗木在干旱逆境下的生长与评价提供信息。
1 材料与方法
1.1试验材料
试验所用种子于2010年1月采自云南省楚雄州白马河林场,千粒质量25.39 g,均采自云南松优良母树,混合采种,混合制种,播种育苗,干旱胁迫试验选取2年生,生长较为一致的盆栽苗进行。
1.2试验方法
试验在西南林业大学塑料温棚内进行,塑料温棚内的湿度在8:00、14:00与20:00分别为67%、29%与59%。参考相似的研究[13-14],并结合预试验确定试验的3个梯度(处理),分别为干旱胁迫0 d(对照)、干旱胁迫5 d和干旱胁迫10 d,试验设置3次重复。每次浇水前采样,测定苗木的渗透调节物质与保护酶活性含量,所取样品均为顶端向下的第1轮枝与第2轮枝中部生长正常的针叶,测定其各项生理指标。
1.3苗木生理特征的测定
电导率(EI)采用电导仪测定[15],丙二醛(MDA)含量采用硫代巴比妥酸法测定[16-18],超氧化物歧化酶(SOD)活性采用核黄素-NBT法测定[17],脯氨酸(Pro)含量采用茚三酮比色法测定[18-19],可溶性糖(SS)含量采用蒽酮乙酸乙酯比色法测定[20]。
1.4数据的处理
采用Excel和SPSS13.0进行统计分析,对生理生化指标进行Excel统计分析整理、绘图,SPSS差异显著性检验。
2 结果分析
2.1不同胁迫天数对苗木质膜相对电导率的影响
质膜相对电导率是细胞膜透性的反映,可以直接反映出细胞被伤害的程度[21]。不同干旱胁迫天数处理下,云南松苗木相对电导率变化见图1。由图1可知,正常浇水处理下相对电导率最小,随着干旱胁迫天数的增加质膜相对电导率不断上升,干旱胁迫10 d时较高,达0.96,表明随着干旱胁迫的加剧,云南松苗木质膜受到的伤害程度增加。经方差分析表明,各处理间的差异未达到检验水平(P= 0.211>0.05),表明不同干旱胁迫天数对云南松苗木质膜相对电导率的影响差异不显著。
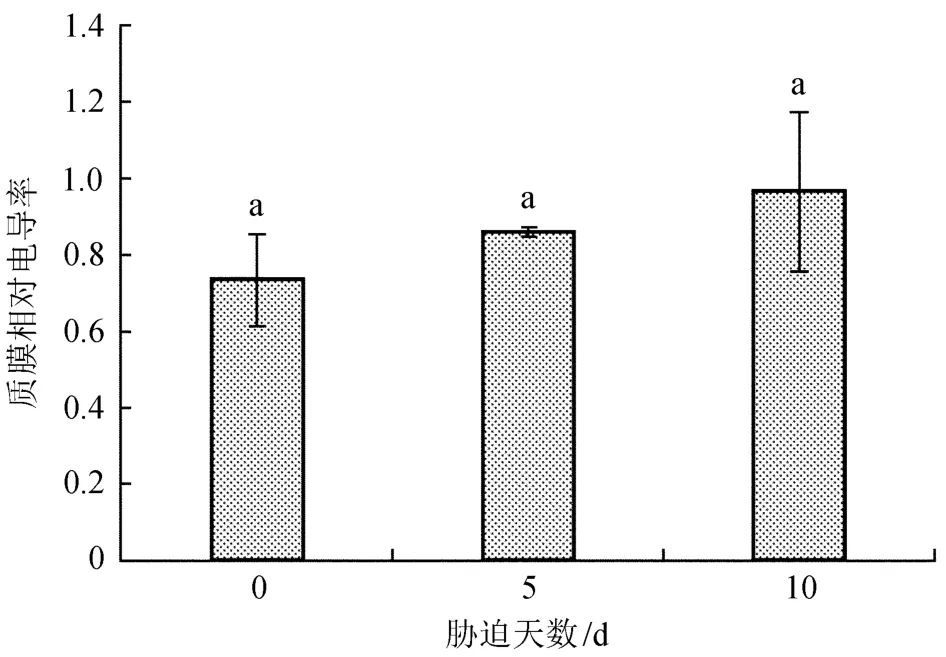
图1 不同胁迫天数云南松苗木质膜相对电导率的变化Fig.1 Changes of plasma membrane relative conductivity of Pinus yunnanensis seedlings under different stress days
2.2不同胁迫天数对苗木丙二醛含量的影响
MDA的积累量也是反映细胞被伤害程度的指标之一[21],由图2可知,随着干旱胁迫天数的增加,云南松苗木中丙二醛含量呈现上升趋势,干旱胁迫10 d时丙二醛的含量较高,分别比干旱胁迫5 d与对照的丙二醛含量高出8.336 μmol/ g与1.941 μmol/ g,且干旱胁迫前期丙二醛增加的速度明显快于胁迫后期。方差分析结果可知,各处理间的差异显著(P = 0.024<0.05),表明不同干旱胁迫天数对云南松苗木质膜相对电导率的影响显著,其中干旱胁迫10 d的丙二醛含量显著高于对照的丙二醛含量。

图2 不同胁迫天数云南松苗木丙二醛含量的变化Fig.2 Changes of malondialdehyd(MDA)content of Pinus yunnanensis seedlings under different stress days
2.3不同胁迫天数对苗木超氧化物歧化酶活性的影响
超氧化物歧化酶是植物抗氧化系统的第一道防线[22],没有胁迫发生时,细胞内的自由基含量变化不大,处于平衡状态。随着干早胁迫程度的加大(图3),云南松苗木中超氧化物歧化酶活性呈现上升的趋势,干旱胁迫10 d时,超氧化物歧化酶含量是干旱胁迫5 d的1.061倍,是对照的1.189倍。方差分析表明,各处理间的差异极显著(P=0.000<0.01),表明不同干旱胁迫天数对云南松苗木超氧化物歧化酶活性有极显著的影响,3个处理两两间均存在显著差异。

图3 不同胁迫天数云南松苗木超氧化物歧化酶活性的变化Fig.3 Changes of superoxide dismutase activity of Pinus yunnanensis seedlings under different stress days
2.4不同胁迫天数对苗木脯氨酸含量的影响
脯氨酸的溶解度很强,具有保持细胞渗透平衡与持水的作用[23]。由图4可知,随着干旱胁迫天数的增加,云南松苗木中脯氨酸含量呈不断上升的趋势,干旱胁迫天数为10 d时,脯氨酸的含量是对照的125%。与此同时,干旱胁迫前期(对照至胁迫5 d)脯氨酸的增加速度明显快于胁迫后期(胁迫5~10 d)。方差分析可知各处理间的差异未达到检验水平(P = 0.071>0.05),即不同干旱胁迫天数对云南松苗木中脯氨酸含量差异不显著,但总体表现为:在本研究处理范围内,随着胁迫程度的增加,脯氨酸含量也随之增加。

图4 不同胁迫天数云南松苗木脯氨酸含量的变化Fig.4 Changes of proline content of Pinus yunnanensis seedlings under different stress days
2.5不同胁迫天数对苗木可溶性糖含量的影响
可溶性糖是细胞中重要的渗透调节物质之一,植物通过增加体内的可溶性糖含量来调节植物组织的渗透势[23]。由图5可知,随着干早胁迫天数的增加,云南松苗木中可溶性糖含量呈现增加的趋势,与对照相比,各干旱胁迫处理中可溶性糖含量均增大。干旱胁迫5 d时可溶性糖的含量比对照有明显升高,当干旱胁迫增加到10 d时,可溶性糖含量增至对照的1.62倍。进一步方差分析表明,各处理间的差异极显著(P = 0.000<0.01),也就是说不同干旱胁迫天数对云南松苗木可溶性糖含量的影响较为明显,且3个处理两两间均存在显著差异。
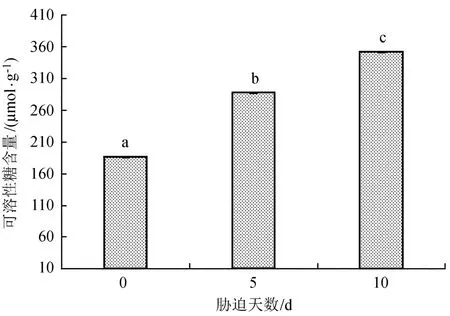
图5 不同胁迫天数云南松苗木可溶性糖含量的变化Fig.5 Changes of soluble sugar content of Pinus yunnanensisseedlings under different stress days
随着干旱胁迫天数的增加质膜相对电导率、丙二醛、超氧化物歧化酶、脯氨酸与可溶性糖都呈上升趋势,本研究中,各指标的峰值均出现在干旱胁迫10 d时。不同胁迫程度下各指标的变化存在差异,干旱胁迫5 d时质膜相对电导率、丙二醛、超氧化物歧化酶、脯氨酸与可溶性糖较对照的升幅分别为19.49%、39.05%、12.13%、17.61%和54.86%;干旱胁迫10 d时质膜相对电导率、丙二醛、超氧化物歧化酶、脯氨酸与可溶性糖较干旱胁迫5 d时的升幅分别为12.15%、8.66%、6.15%、4.27%和23.76%,这些指标均表现为胁迫前期值的变化幅度大于胁迫后期值的变化幅度。
3 结论与讨论
脯氨酸、可溶性糖等渗透调节物质与超氧化物歧化酶是植物体在长期进化过程中形成保护系统,丙二醛与质膜相对电导率反映细胞受伤害程度,细胞中脯氨酸、可溶性糖、超氧化物歧化酶、丙二醛和质膜相对电导率含量的变化与植物的抗旱强弱有一定的关系[24-27]。本研究中,不同干旱胁迫对云南松苗木渗透调节物质及保护酶活性的影响在处理间可溶性糖含量、超氧化物歧化酶呈极显著差异,丙二醛含量差异显著,而脯氨酸与质膜相对电导率差异不显著,引起差异的主要原因是不同生理指标变化对干旱胁迫的响应不同,云南松苗木可溶性糖、超氧化物歧化酶与丙二醛对干旱胁迫变化较为敏感,而质膜相对电导率与脯氨酸含量对干旱胁迫变化不敏感。云南松苗木在干旱胁迫下渗透调节能力和保护酶活性的差异表明,云南松苗木在干旱胁迫环境下各项生理指标的抗旱相关性与抗旱能力不同。
质膜相对电导率是衡量细胞膜透性的重要指标,电解质的渗漏量越多,细胞膜受害程度越重。植物器官衰老或在逆境条件下遭受伤害往往发生膜脂过氧化作用[28],丙二醛是细胞膜脂过氧化的产物,所以丙二醛的积累会对细胞造成伤害,二者同时反映了细胞受伤害的程度。本研究中在干旱胁迫条件下云南松苗木抗旱性相关性次序为丙二醛>质膜相对电导率。由于脯氨酸和可溶性糖都具有维持细胞持水的作用,所以两者之间具有相互补偿的作用[26],且渗透调节物质对干旱胁迫的响应因胁迫程度的不同而表现不同。由渗透调节物质较对照的升幅可以看出,可溶性糖含量变化对干旱胁迫程度变化的响应较强烈,而脯氨酸含量响应较弱,可见渗透调节类型对干旱胁迫调节作用存在差异,调节能力表现为可溶性糖>脯氨酸,可溶性糖和脯氨酸为主要的渗透调节因子,在云南松苗木细胞内可溶性糖渗透调节能力大于脯氨酸,在干旱胁迫后期较前期的升幅也可以看出干旱胁迫后期各种渗透调节物质含量增加的幅度也表现为可溶性糖高于脯氨酸。干旱胁迫后期与前期较对照的升幅相比可知从对照到干旱胁迫前期时各项生理指标的升幅较大,在干旱胁迫后期,云南松苗已经逐渐适应了干旱胁迫的环境,其生理指标的变化幅度降低,这表明云南松具有一定的耐旱性。超氧化物歧化酶的活性可以间接反映植物的抗旱性[29],在本研究中,不同干旱胁迫天数下,云南松苗木超氧化物歧化酶活性上升,细胞通过提高体内超氧化物歧化酶的活性来抵御干旱对自身生长造成的不利影响。
膜的伤害与渗透调节是同步进行的,云南松苗处于干旱逆境时,细胞内的渗透调节物质与保护酶活性的变化说明云南松通过调节细胞内渗透调节物质与保护酶活性抵御干旱胁迫,当遭遇干旱胁迫,云南松苗木细胞内的渗透调节物质与保护酶共同作用,以降低云南松苗木受干旱胁迫影响的程度。综上所述,云南松苗木在干旱胁迫后一系列生理指标的变化表明云南松苗木具有一定的耐旱性。
[参考文献]
[1]陈飞,王健敏,孙宝刚,等.云南松的地理分布与气候关系[J].林业科学研究,2012,25(2):163-168.
[2]中国科学院昆明植物所.云南植物志:第4卷[M].北京:科学出版社,1986:54-55.
[3]吴红军,谢耀坚,王志华.西南地区特大干旱与桉树种植关系分析[J].林业经济,2010(6):120-122.
[4]韩兰英,张强,马鹏里,等.西南地区农业干旱灾害综合风险空间特征[C]/ /中国气象学会.第31届中国气象学会年会S5干旱灾害风险评估与防控.中国气象学会,2014:9.
[5]韩兰英,张强,马鹏里,等.中国西南地区农业干旱灾害风险空间特征[J].中国沙漠,2015,35(4):1015-1023.
[6]许建定.土壤水分和氮磷营养对植物根系生长的影响[J].山西水土保持科技,2010(3):12-15.
[7]郭京衡,李尝君,曾凡江,等.2种荒漠植物根系生物量分布与土壤水分、养分的关系[J].干旱区研究,2016,33(1):166-171.
[8]吕桂英.水分对作物生长发育和产品质量的影响[J].中国种业,2007(3):61-62.
[9]孙振蓉,陈斌,张学霞.锡林郭勒盟农牧业干旱风险评价[J].自然灾害学报,2014,23(5):129-139.
[10]黄瑞.议乡土树种在云南林业生态建设中的作用[J].城市建设理论研究(电子版),2013(12):1-4.
[11]赵丽芝,蔡年辉,夏清柱,等.干旱胁迫对云南松种子萌发及渗透调节物质的影响[J].西南林业大学学报,2012,32(2):21-25.
[12]王晓丽,曹子林,朱霞.PEG处理对云南松种子发芽及其生理生化特性的影响[J].西北林学院学报,2012,27(6):16-19.
[13]韩瑞宏.野生假俭草抗旱性研究[D].成都:四川农业大学,2003.
[14]卢素锦,周青平,刘文辉,等.披碱草属6种牧草苗期抗旱性鉴定[J].湖北农业科学,2013,52(8):1889-1892.
[15]Blum A,Ebercon A.Cell Membrane stability as a measure of drought and heat tolerance in wheat1[J].Crop Science,1981,21(1):43-47.
[16]西北农业大学植物生理生化教研室.植物生理实验指导[M].西安:陕西科学技术出版社,1987:51-55.
[17]Giannopolitis C N,Ries S K.Superoxide dismutases:I.Occurrence in higher plants[J].Plant Physiology, 1977,59(2):309-314.
[18]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000:161-162.
[19]朱广廉.植物体游离脯氨酸的测定[J].植物生理学通讯,1986,22(1):35-37
[20]化黎玲,胡明月,袁俊云,等.香樟大树相对电导率测定试验[J].山东林业科技,2013,43(1):65.
[21]郭春芳,孙云.干旱胁迫下植物的渗透调节及脯氨酸代谢研究进展[J].福建教育学院学报,2015,16(1):114-118.
[22]李州,彭燕,苏星源.不同叶型白三叶抗氧化保护及渗透调节生理对干旱胁迫的响应[J].草业学报,2013,22(2):257-263.
[23]蒋宣斌,耿养会,彭秀,等.常绿岩垂草抗旱生理特性分析[J].西南林业大学学报,2012,32(4):17-21.
[24]刘建兵.马尾松苗木渗透调节物质与耐旱性关系研究[D].南京:南京林业大学,2008.
[25]安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干旱胁迫的生长与生理响应[J].生态学报,2011,31(3):716-725.
[26]刘艳,陈贵林,蔡贵芳,等.干旱胁迫对甘草幼苗生长和渗透调节物质含量的影响[J].西北植物学报,2011,31(11):2259-2264.
[27]王海珍,徐雅丽,张翠丽,等.干旱胁迫对胡杨和灰胡杨幼苗渗透调节物质及抗氧化酶活性的影响[J].干旱区资源与环境,2015,29(12):125-130.
[28]胡俊靖,陈卫军,郭子武,等.水分胁迫对竹子生理特性影响的研究进展[J].西南林业大学学报,2015,35(1):91-95.
[29]张智猛,宋文武,丁红,等.不同生育期花生渗透调节物质含量和抗氧化酶活性对土壤水分的响应[J].生态学报,2013,33(14):4257-4265.
(责任编辑张坤)
第1作者:孙琪(1992—),女,硕士生。研究方向:林木遗传育种。Email:camugn@sina.com。
Drought Stress on Physiological Characteristics of Pinus yunnanensis Seedlings
Sun Qi1,Cai Nianhui1,2,Chen Shi2,Wang Dawei1,Duan Anan1,Xu Yulan1
(1.Key Laboratory for Forest Genetic and Tree Improvement&Propagation in Universities of Yunnan Province,Southwest Forestry University,Kunming Yunnan 650224,China;2.Key laboratory for Forest Resources Conservation and Use in the Southwest Mountains of China,Ministry of Education,Southwest Forestry University,Kunming Yunnan 650224,China)
Abstract:Effects of drought stress on osmotic adjustment substance and protective enzyme activity of Pinus yunnanensis seedlings were studied by the design of the different drought interval days.The results showed that within a certain range,membrane relative conductivity,malondialdehyd,proline,soluble sugar content and superoxide dismutase activity of P.yunnanensis seedlings increased with the extension of the drought interval days.But ranges of the increases were different due to the difference of the drought interval days and the type of osmotic adjustment substance.There were significantly different among treatments(drought interval days)in soluble sugar and superoxide dismutase,malondialdehyd content had great differences and no obvious difference among treatments for proline and plasma membrane relative conductivity.Osmotic adjustment ability of soluble sugar was higher than proline.Malondialdehyd has higher correlation with the drought tolerance compared with plasma membrane relative conductivity.The response of drought stress was weak for proline and plasma membrane relative conductivity.The damage of plasma membrane,osmotic adjustment and superoxide dismutase protection were carried out simultaneously,which resulted in the drought tolerance of P.yunnanensis seedlings under drought stress.
Key words:Pinus yunnanensis,drought stress,osmotic adjustment,enzyme activity,drought tolerance
通信作者:许玉兰(1979—),女,副教授,硕士生导师。研究方向:林木遗传育种。Email:xvyulan@163.com。
基金项目:西南林业大学林学学位授权学科经费资助青年后备人才研究项目资助(5009750101-1);西南地区生物多样性保育国家林业局重点实验室开放基金资助。
收稿日期:2015-12-25
doi:10.11929/ j.issn.2095-1914.2016.03.004
中图分类号:S718.43
文献标志码:A
文章编号:2095-1914(2016)03-0018-05
