云南松不同茎干类型群体针叶性状表型多样性比较
2016-06-04邓丽丽孙琪许玉兰周丽徐杨李德龙罗元陈诗李根前蔡年辉西南林业大学云南省高校林木遗传改良与繁育重点实验室云南昆明6504西南林业大学西南山地森林保育与利用省部共建教育部重点实验室云南昆明6504
邓丽丽孙 琪许玉兰周 丽徐 杨李德龙罗 元陈 诗李根前蔡年辉,(.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南昆明6504;.西南林业大学西南山地森林保育与利用省部共建教育部重点实验室,云南昆明6504)
云南松不同茎干类型群体针叶性状表型多样性比较
邓丽丽1孙 琪1许玉兰1周 丽1徐 杨1李德龙1罗 元1陈 诗2李根前2蔡年辉1,2
(1.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南昆明650224;2.西南林业大学西南山地森林保育与利用省部共建教育部重点实验室,云南昆明650224)
摘要:在宜良、新平和禄丰3个地点各选择1个有代表性的林分,每个林分从林冠层和更新层分别按通直和扭曲的树干表型性状采集针叶,并对7个针叶表型性状进行测量和比较。结果表明:在林冠层和更新层中,通直群体和扭曲群体间并不存在明显差异,云南松林冠层通直群体和扭曲群体的针叶性状(针叶长度、针叶粗度、针叶束粗、叶鞘长度)平均值均大于更新层,而云南松针叶的形状指数(针叶长度/针叶粗度、针叶长度/叶鞘长度、针叶束粗/针叶粗度)在林冠层和更新层间差异不大;云南松针叶性状在群体间和群体内均存在极其丰富的变异,7个针叶性状指标的变异系数按大小顺序为NL/ FSL(22.20%)> NL/ NW(19.15%)> FSL(19.04%)> NW (19.00%)> NL(16.61%)> FW(15.10%)> FW/ NW(13.20%);云南松针叶性状大部分表现为群体间变异小于群体内变异,群体间总体平均表型分化系数为20.90%,群体内的变异占79.10%,说明群体内变异是云南松针叶性状变异的主要来源。
关键词:云南松;针叶;表型多样性;干型
云南松(Pinus yunnanensis)为松科(Pinaceae)常绿针叶乔木,云南松以云南高原为起源中心和分布中心,分布于东经96°~108°、北纬23°~30°[1- 2]。云南松是重要的用材树种,也是荒山造林的先锋树种,为云南省现存面积最大的森林类型,占云南省林分总面积的19.63%、森林总蓄积量的14.28%[3-4]。云南松具有生长较快、材质较好、耐干旱瘠薄、天然更新能力强等优良特性,在我国西南地区被广泛用于人工造林,在国民经济建设和生态建设中发挥着重要作用[5-6]。
遗传多样性是种质资源保护与评价的重要指标,而用形态特征或表型性状来反映遗传多样性最为传统、简单易行,且至今仍较为有效[7-8];伴随着生物多样性保护的积极开展,遗传多样性的研究越来越受到重视,已经成为生物多样性研究的核心[9]。1个树种遗传多样性的高低决定了其对环境的适应能力,多年生林木种内分布的丰富变异,是维持森林生态系统长期稳定的基础[10-13]。松属(Pinus)植物是雌雄同株、典型的风媒传粉异交植物,具有庞大的核基因组和十分复杂的基因家族。因此,在松属植物中存在着极其丰富的变异[14]。植物表型性状的变异有其独特的遗传基础,受遗传与环境、调控基因与结构基因综合作用的影响[15]。林木遗传变异主要包括地理种源变异、林分、个体以及个体内变异4个水平,这些变异体现在表型、细胞、生化和分子等不同水平上[16]。种子和球果性状的表型变异是研究植物群体多样性的一个重要组成部分,而叶作为植物十分重要的构件结构,不但进行光合作用,而且直接与外界环境接触,因此更容易发生性状的变异以适应不同的环境条件[17-18];通常所说的表型特征是指对叶形态和其他生长性状的研究,其中叶形态是一个重要的表型特征,与植物所处环境的生态因子,以及植物的繁殖有着密切的关系,因而具有较高研究价值[19]。本研究在宜良、新平和禄丰3个地点,每个地点确定1个有代表性的天然林作为采样林分,在各林分的林冠层和更新层,按茎干表现分为通直和扭曲(包括弯曲)2个类型,分别采集针叶比较不同类型、不同林层云南松针叶性状的变异情况,以了解云南松不同类型、林层间针叶表型多样性的变化规律,以期为可持续经营策略的制定提供科学依据,为云南松表型衰退的综合评价及其遗传多样性的保护奠定基础。
1 研究方法
1.1研究材料
本试验所用针叶材料采集于云南省昆明市宜良县(YL)、玉溪市新平县(XP)和楚雄州禄丰县(LF)3个地区。每个地区确定1个采样林分,林分内按更新层和林冠层2个林层各2个类型采集,即林冠层通直类型(LS)、更新层通直类型(GS)、林冠层扭曲类型(LT)和更新层扭曲类型(GT),为便于后述分析描述,各林层每个类型所采样按1个群体来看待,即林冠层通直群体、更新层通直群体、林冠层扭曲群体和更新层扭曲群体共4个群体,在每个群体中选取50~60株生长正常、无明显病虫害的个体植株,采集树冠中上部位的2年生针叶,所采单株间距离为5倍树高以上。
1.2性状测定
参考前人的研究[20],在所采集的枝条上随机选取10束健康完整的针叶,用直尺测量针叶长度(NL)、叶鞘长度(FSL),精确到0.1 cm;用游标卡尺测量针叶的粗度(NW)、针叶束粗(FW),精确到0.01 mm;计算针叶长/针叶粗(NL/ NW)、针叶长/叶鞘长(NL/ FSL)、针叶束粗/针叶粗(FW/ NW)等7个针叶表型性状指标,其中NL/ NW、NL/ FSL、FW/ NW为云南松针叶的形状指数。
1.3数据处理
将3个地区共12个群体每个个体的7个性状作为特征量,用Excel、SPSS、SAS等软件进行处理,可以得到每个性状的平均值、平方和、均方、标准差、F值、显著水平、变异系数、方差分量以及表型分化系数等,其中用表型分化系数(Vst)来反映群体间的表型分化状况,计算公式为:

式中:δ2t/ s为群体间方差分量,δ2s为群体内方差分量[21]。
用变异系数CV来衡量表型性状的离散程度,即变异系数越大,离散程度越高,其计算公式为:

式中:S为标准差,X为平均值[12]。
2 结果与分析
2.1云南松针叶性状在群体间的变异
对3个地区各4个群体云南松针叶性状的平均值和标准差分析结果见表1。
在同一林层内,通直群体和扭曲群体间各针叶性状的差异并不明显;在不同林层间,通直群体和扭曲群体的针叶长/针叶粗、针叶长/叶鞘长和针叶束粗/针叶粗3个形状指数性状在林冠层和更新层间差异不大,而林冠层针叶长度、针叶粗度、针叶束粗和叶鞘长度的平均值均大于更新层,说明随着云南松林木的生长,云南松的针叶性状指标有增长的趋势。
2.2云南松针叶性状在群体内的变异
对云南松针叶各表型性状指标的变异系数的分析中可以看出(图1),云南松针叶表型性状存在较大的变异系数,变异范围在7.77%~27.61%之间。不同性状在不同群体内的变异系数也存在一定的波动,7个针叶性状的变异系数按大小顺序为NL/ FSL(22.20%)> NL/ NW(19.15%)> FSL (19.04%)> NW(19.00%)> NL(16.61%)> FW (15.10%)> FW/ NW(13.20%)。在同一林层中,通直群体和扭曲群体的各项表型性状的变异系数差异不大;在不同林层间,各表型性状的变异系数表现为更新层的大于林冠层的,表明更新层中,云南松针叶表型的变异较大,针叶性状不整齐,较林冠层具有更高的变化幅度。
2.3不同群体间云南松针叶性状的比较
2.3.1林冠层通直群体和扭曲群体间的变异分布
从林冠层的通直群体和扭曲群体的方差分析结果可以看出(表2),表型分化很小,3个地区分别是1.01%、2.88%和8.74%,总体平均值为4.21%。说明在林冠层,各针叶性状的变异几乎来源于群体内,群体间只存在极少数的变异。云南松7个针叶表型性状在群体内均存在极显著的差异,但在群体间表现不一样,其中新平县的叶鞘长度和针叶长度/叶鞘长度、禄丰县的针叶粗度、针叶束粗、针叶长度/针叶粗度存在显著或极显著的差异,其他性状均不存在显著的差异。
2.3.2更新层通直群体和扭曲群体间的变异分布
对更新层的通直群体和扭曲群体进行方差分析,所得结果如表3,由表3可知,云南松针叶表型性状在群体内均存在极显著的差异,而群体间的变异在不同性状、不同地区存在一定的波动,在新平县,针叶的各性状在群体间均不存在显著差异,在宜良县和禄丰县,少部分性状在群体间存在极显著差异。但不同地区不同群体表型分化系数呈现差异,在新平县,各性状的平均表型分化系数很小,仅有0.69%的变异来源于群体间,宜良县,针叶粗度、针叶束粗、针叶长度/针叶粗度3个性状的群体内方差分量小于群体间,其余性状表现为群体内的方差分量大于群体间,平均表型分化系数为22.95%;禄丰县各性状的表型分化系数均低于50%,平均为13.17%,各性状的变异主要来源于群体内。

图1 云南松针叶表型性状的变异系数Fig.1 The coefficient variation of coniferous phenotypic traits of P.yunnanensis Franch

表2 林冠层通直群体和林冠层扭曲群体间表型性状方差分析Table 2 Variance analysis of phenotypic characteristics between LS and LT
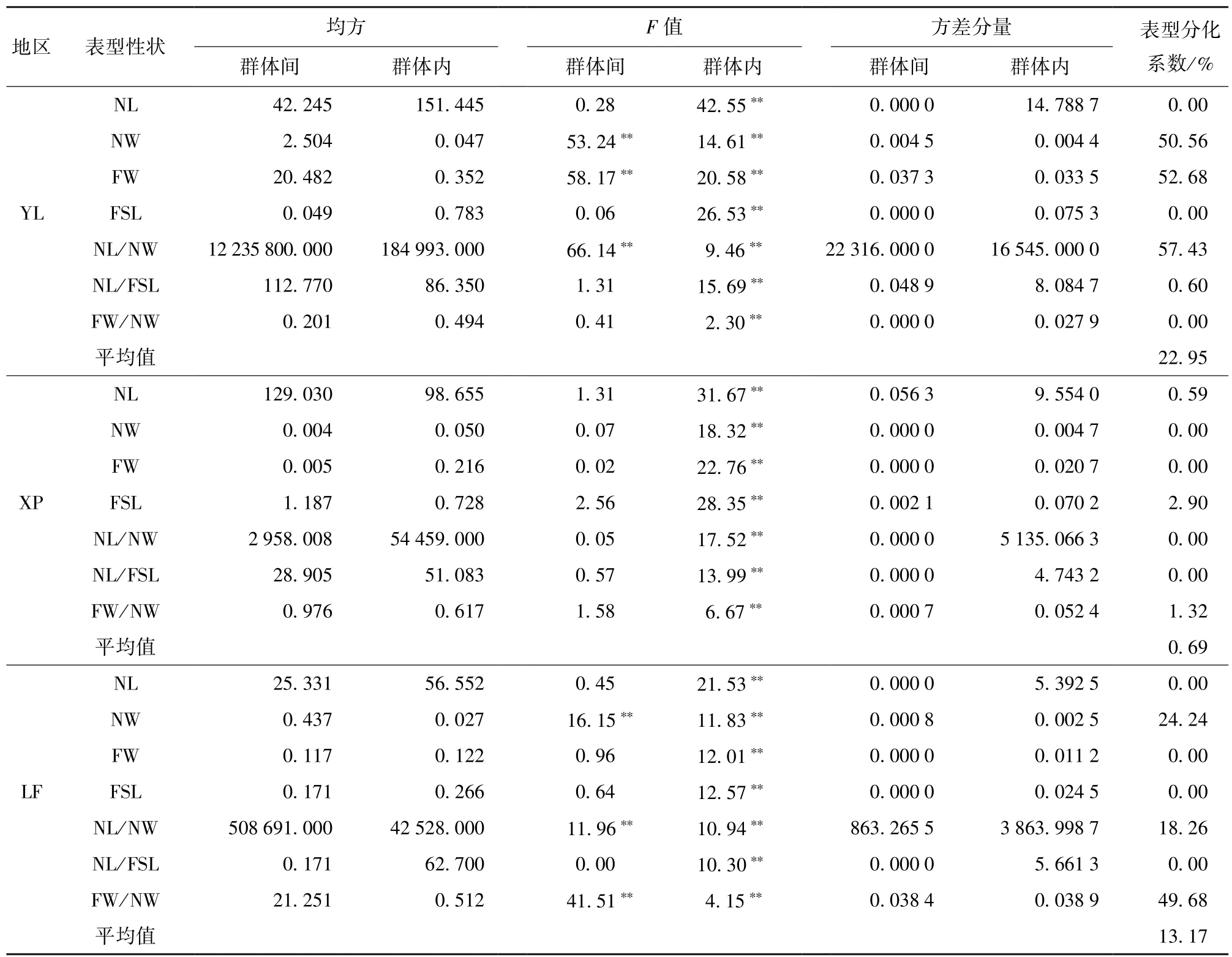
表3 更新层通直群体和更新层扭曲群体间表型性状方差分析Table 3 Variance analysis of phenotypic characteristics between LS and GS
2.3.3林冠层和更新层间通直群体的变异分布
从云南松林冠层和更新层间通直群体的方差分析结果可知(表4),云南松7个针叶性状在群体内均存在极显著的差异,在群体间,针叶长度、针叶粗度、叶鞘长度和针叶束粗4个性状存在极显著的差异,而3个形状指数性状(针叶长/针叶粗、针叶长/叶鞘长、针叶束粗/针叶粗),在不同的地区显示出不同的差异性。7个性状的平均分化系数在3个地区分别为22.24%、23.63%和49.43%,平均表型分化系数为31.77%,云南松总体的针叶性状表型变异的来源在群体内占68.23%。但不同地区各性状表型分化系数有一定的差异,在新平县,针叶束粗和叶鞘长度的表型分化系数在群体间和群体内的比例约为1∶1,说明这2个性状的变异50%来源于群体间50%来源于群体内;在禄丰县,针叶长度、针叶粗度、叶鞘长度和针叶束粗4个性状的群体间表型分化系数大于群体内,说明在这个地区表型的变异主要来源于群体间。
2.3.4扭曲群体林冠层和更新层间的变异分布
对云南松扭曲群体的林冠层和更新层进行表型性状方差分析(表5),结果显示,在3个地区绝大部分针叶性状在群体内均存在极显著的差异;在群体间,针叶长度、针叶粗度、叶鞘长度和针叶束粗4个性状(除新平地区的叶鞘长度性状)均存在极显著的差异,对于其他3个形状指数,针叶束粗/针叶粗度在群体间存在显著或极显著的差异,另外两个性状差异不显著。3个地区的表型分化系数分别为44.12%、19.07%和42.88%,平均为35.36%,小于群体内65.64%,说明针叶性状的表型变异主要来源于群体内,但不同地区不同性状的表型分化趋势存在一定的变异,如宜良和禄丰县,针叶长度、针叶粗度和针叶束粗这3个性状的群体间表型分化系数大于群体内。

表4 林冠层通直群体和更新层通直群体间表型性状方差分析Table 4 Variance analysis of phenotypic characteristics between LS and GS

表5 扭曲群体林冠层和扭曲群体更新层间表型性状方差分析Table 5 Variance analysis of phenotypic characteristics between LT and GT
在上述4种成对群体间的比较中,云南松针叶性状群体内的总体平均表型分化系数(79.10%)大于群体间(20.90%),总体上来看,云南松针叶的变异主要来源于群体内,其中以林冠层通直群体和扭曲群体间的比较最为明显,群体间的表型分化系数平均值仅为4.21%,说明变异几乎来源于群体内。但在不同的地区,会存在少数针叶性状的群体间方差分量大于群体内,如在林冠层和更新层间通直群体的比较中,禄丰地区云南松的针叶长度、针叶粗度、叶鞘长度和针叶束粗4个性状群体间方差分量大于群体内;在林冠层和更新层间扭曲群体的比较中,宜良和禄丰县的云南松针叶长度、针叶粗度和针叶束粗3个性状也表现为群体间的变异大于群体内的变异;在更新层通直群体和扭曲群体间的比较中,宜良县云南松针叶粗度、针叶束粗、针叶长度/针叶粗度3个性状表现出以群体间的变异占多数。另外,在4种成对群体间的比较来看,云南松7个针叶性状在群体内均存在极显著的差异,在群体间,云南松针叶性状也表现出显著或极显著差异,而以形状指数的差异小些。
3 结论与讨论
植物表型性状既有变异性又有稳定性,其变异在适应性和进化上均具有重要意义[22-23]。本研究中,在通直群体和扭曲群体内,云南松林冠层的针叶性状,包括针叶长度、针叶粗度、叶鞘长度和针叶束粗4个性状的平均值较更新层的高,这是由于云南松林木处于生长期,针叶性状指标均会有所增长;而其他3个形状指数,包括针叶长/针叶粗、针叶长/叶鞘长、针叶束粗/针叶粗在不同林层间差异不大,这说明了云南松针叶的形状指数变异较其他单个性状的变异稳定。在同一林层内,云南松针叶性状在通直群体和扭曲群体间的差异并不明显。因此,云南松择伐时根据茎干长势去优留劣或去劣留优对针叶表型多样性的变化影响不大,但考虑到云南松以天然更新为主,去劣留优可保留长势较好的母树作为更新母树,从而有利于云南松茎干的改良,以提高云南松林的利用价值与经济效益。
植物群体中保持大的变异对于群体是有利的,群体内多种基因型所对应的表型范围越广,可以使群体整体适应环境的变化性[24]。研究表明,云南松针叶性状变异系数波动于7.77%~27.61%之间,各性状均存在较大的变异,这个结果与川西云杉(Picea balfouriana)针叶表型变异[25]的结果相似。在不同林层间,更新层各针叶表型性状的变异系数较林冠层的变异范围更大。因此,在进行优良繁育材料的选择时,可以考虑在更新层中进行初选,以期选择出极端的变异类型,然后跟踪观察,待生长稳定后,进一步确定所选的种质材料。从结果可以发现,云南松针叶同一性状的变异在不同地区间存在着一定的差异,这说明不同环境条件下也可导致表型性状变异的不同,今后可进行深入研究,以探讨环境与遗传变异间的关系。
云南松针叶群体间性状总体平均表型分化系数为20.90%,这与前人的研究比较相似,Hamrick等[26]的研究表明,异交虫媒植物只有19.7%的遗传变异存在于群体间,绝大部分存在于群体内。与其它针叶树相比,云南松针叶性状的总体平均表型分化系数(20.90%),高于马尾松(Pinus massoniana)(6.44%)[21]和云南红豆杉(Taxus yunnanensis)(14.66%)[27],低于长白落叶松(Larix olgensis)(26.2%)[28]、青海云杉(Picea crassifolia)(27.18%)[29]、川西云杉(Picea likiangensis var.balfouriana)(36.53%)[25]、油松(Pinus tabulaeform)(38.97%)[30]和杜松(Juniperus regida)(66.63%)[31]等。云南松针叶表型性状的绝大部分变异来源于群体内,各针叶性状在群体内均存在极显著的差异。这可能是云南松的生物学特性所造成,云南松是典型的长寿命风媒传粉植物,结实率高,群体内随机交配程度相当高,群体间基因流动频率较大。因此,云南松群体内具有较高的遗传变异,遗传多样性十分丰富[20,32]。云南松针叶的表型变异虽然主要来源于群体内,但也有少数性状以群体间的变异占主导,如林冠层和更新层间扭曲群体的比较来看,云南松针叶性状表型分化系数最高达到了84.96%。尽管群体内的变异远大于群体间变异,但群体间变异反映了地理和生殖隔离上的变异,群体间的多样性变异也是种内多样性的重要组成部分[33]。由于变异主要来源于群体内,在以遗传多样性为保护目的时,可保存少数群体、且每个群体保存尽可能多的个体为宜,以更加完整地保护云南松的遗传多样性,为云南松的可持续发展与利用奠定物质基础。
[参考文献]
[1]金振洲,彭鉴.云南松[M].昆明:云南科技出版社,2004:1-66.
[2]黄瑞复.云南松的种群遗传与进化[J].云南大学学报(自然科学版),1993,11(1):50-63.
[3]中国科学院中国植物志编辑委员会.中国植物志:第7卷[M].北京:科学出版社,1978:255-259.
[4]邓喜庆,皇宝林,温庆忠,等.云南松林资源动态研究[J].自然资源学报,2014,29(8):1411-1419.
[5]吴征镒,朱彦承,姜汉侨.云南植被[M].北京:科学出版社,1987:417-419.
[6]周丽,陈诗,徐杨,等.云南松不同种源苗木光合特性比较[J].西南林业大学学报,2015,35(5):8-13.
[7]Schaal B A,Leverich W J,Rogstad S H.A comparison of methods for assessing genetic variation in plant conservation biology[M]/ / Falk D A,Holsinger K E.Genetics and Conservation of Rare Plants.New York:Oxford University Press,1991:123-124.
[8]葛颂,洪德元.遗传多样性及其检测方法[M]/ /钱迎倩,马克平.生物多样性研究的原理与方法.北京:中国科学技术出版社,1994:123-140.
[9]顾万春,王棋,游应天,等.森林遗传资源学概论[M].北京:中国科学技术出版社,1998:364-913.
[10]张泽.巢式设计的多元分析.生物数学学报[J].1996,11(4):83-87.
[11]蔡永立.中国东部亚热带青冈果实形态变异的研究[J].生态学报,1999,19(4):581-584.
[12]李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[13]曾杰,郑海水,甘四明,等.广西西南桦天然居群的表型变异[J].林业科学,2005,41(2):59-65.
[14]李斌,顾万春.松属植物遗传多样性研究进展[J].遗传,2003,25(6):740-748.
[15]周连第,兰彦平,曹庆昌,等.板栗叶片性状表型多样性研究[J].中国农学通报,2005,21(9):136-139.
[16]王明庥.林木遗传育种学[M].北京:中国林业出版社,2001:1.
[17]邹春静,卜军,徐文铎.长白松种群针叶性状及其变异的研究[J].生态学杂志,1995,14(2):18-22.
[18]Chechowitz N,Chappell D M,Guttman S I,et al.Morphologieal,electrophoretic and ecological analysis of Quercus macrocarpa population in the Black Hills of South Dakota and Wyoming[J].Canadian Journal of Botany,1990,68(10):2185-2194.
[19]顾万春.统计遗传学[M].北京:科学出版社,2004:1-422.
[20]Xu Y L,Woeste K,Cai N H,et al.Variation in needle and cone traits in natural populations of Pinus yunnanensis[J].Journal of Forestry Research,2015: DOI:10.1007/ s11676-015-0153-6.
[21]葛颂,王明庥,陈岳武.用同工酶研究马尾松群体的遗传结构[J].林业科学,1998,24(11):399-409.
[22]杨继.植物种内形态变异的机制及其研究方法[J].武汉植物学研究,1991,9(2):185-195.
[23]Baeilieri R,Ducousso A,Kremer A.Genetic morphological ecological and phonological differentiation between Quercu spetraea(Matt.)Label.And Quercu robur L.in a mixed stand of Northwest of France[J].Silvae Genetica,1995,44(1):1-10.
[24]竺利波,顾万春,李斌.紫荆群体表型性状多样性研究[J].中国农学通报,2007,23(3):138-145.[25]辜云杰,罗建勋,吴远伟,等.川西云杉天然种群表型多样性[J].植物生态学报,2009,33(2):291-301.
[26]Hamric J L,Godt M J W.Allozyme diversity in plant species[M].Brown A H D,Clegg M T,Kahler A L,et al.Plant Population Genetics,Breeding,and Genetic Recourse.Sunderland,Mass:Sinauer,1989:43-46.
[27]陈少瑜,吴丽圆,李江文,等.云南红豆杉天然种群遗传多样性研究[J].林业科学,2001,37(5):41-48.
[28]张含国,高士新,张敏莉,等.长白落叶松天然群体遗传结构的研究[J].东北林业大学学报,1995,23(6):21-31.
[29]王娅丽,李毅,祁连山.青海云杉天然群体的种实性状表型多样性[J].植物生态学报,2008,32 (2):355-362.
[30]姬明飞,张晓玮,韩瑾,等.油松天然群体的种实性状表型多样性研究[J].西北植物学报,2013,33(9):1898-1905.
[31]刘雨,李登武,秦廷松,等.宁夏贺兰山杜松天然群体的表型多样性[J].浙江农林大学学报,2011,28(4):619-627.
[32]舒筱武,郑畹,李思广,等.云南松壮苗培育与幼林生长相关性的研究[J].云南林业科技,2000 (4):1-9.
[33]宋杰,李世峰,刘丽娜,等.云南含笑天然居群的表型多样性分析[J].西北植物学报,2013,33 (2):272-279.
(责任编辑张坤)
第1作者:邓丽丽(1992—),女,硕士生。研究方向:林木遗传育种。Email:denglilimini@163.com。
Compare on the Needle Phenotypic Variations between the Different Type of Trunk Populations of Pinus yunnanensis
Deng Lili1,Sun Qi1,Xu Yulan1,Zhou Li1,Xu Yang1,Li Delong1,Luo Yuan1,Chen Shi2,Li Genqian2,Cai Nianhui1,2
(1.Key Laboratory for Forest Genetic and Tree Improvement&Propagation in Universities of Yunnan Province,Southwest Forestry University,Kunming Yunnan 650224,China;2.Key Laboratory for Forest Resources Conservation and Use in the Southwest Mountains of China,Ministry of Education,Southwest Forestry University,Kunming Yunnan 650224,China)
Abstract:The needles were collected from the different forest stand in Yiliang,Xinping and Lufeng.The plant trees were divided into the straight population and twisted or crooked population based on the morphological characteristics of trunk in canopy and regeneration layer of each forest stand.Seven needle morphological traits were measured for each population.The results showed that there was no difference for seven needle morphological traits between straight population and twisted or crooked population in canopy and regeneration layer.The average values of the needle traits(needle length,needle width,fascicle width,fascicle sheath length)for trees in canopy layer were greater than that for regeneration layer in both straight population and twisted or crooked population.But the needle shape index traits(needle length/ needle width,needle length/ fascicle sheath length,fascicle width/ needle width)had littlebook=31,ebook=34difference between canopy and regeneration layer in both straight population and twisted or crooked population.There were significant differences for each needle morphological trait both among populations and within population.The order of the coefficient variation of seven traits was:NL/ FSL(22.20%)> NL/ NW(19.15%)> FSL(19.04%)> NW (19.00%)> NL(16.61%)> FW(15.10%)> FW/ NW(13.20%).Most needle morphological traits showed the variations among populations was smaller than within population,the mean phenotypic differentiation coefficient had 20.90% among populations and 79.10%within population,which stated that the variation within group was the main source.
Key words:Pinus yunnanensis,needle,phenotypic diversity,type of trunk
通信作者:蔡年辉(1975—),男,讲师。研究方向:森林培育。Email:cainianhui@sohu.com。
基金项目:西南林业大学云南省省级重点学科(林学)资助;国家自然科学基金项目(31360189)资助;西南林业大学云南省高校林木遗传改良与繁育重点实验室开放基金资助(YNGB201504);西南地区生物多样性保育国家林业局重点实验室开放基金资助。
收稿日期:2015-12-28
doi:10.11929/ j.issn.2095-1914.2016.03.006
中图分类号:S722.3
文献标志码:A
文章编号:2095-1914(2016)03-0030-08
