草甘膦对巴西橡胶树芽接苗叶片形态和生理指标的影响
2016-05-30潘敏王萌李晓娜杨叶郑服丛张宇朱朝华
潘敏 王萌 李晓娜 杨叶 郑服丛 张宇 朱朝华


摘 要 以橡胶树品系CATAS7-33-97的芽接苗为材料,研究草甘膦对其叶片形态和生理活性的影响。结果表明:喷施草甘膦导致橡胶树叶片脱落后,引起整蓬新生叶片变长、扭曲。该蓬新生叶片稳定后,下一蓬新生叶片形状会部分恢复。叶片畸形与含水量变化无关,但会导致叶绿体结构变化、过氧化物酶和超氧化物歧化酶活性增加13%~40%、可溶性糖含量下降47%~81%和脯氨酸含量上升15%~389%。
关键词 巴西橡胶树;草甘膦;形态;生理指标
中图分类号 S794.1 文献标识码 A
Abstract Glyphosate, a nonselective herbicide, can damage plants when inappropriate used. In this study, for the first time with rubber tree clone CATAS7-33-97 grafting seedlings as materials, we studied the effects of glyphosate on the leaf morphology and physiological indices of rubber tree. Results showed that the glyphosate caused rubber tree leaf fall, new-grown leaves twist and longer leaves. After these change stabilized, the new-grown leaves would be normal. The leaf doformity had nothing to do with water content, but could result in the change of chloroplast structure, increase peroxidase and superoxide dismutase enzyme activities from 13%-40%, cut down the content of soluble sugar from 47%-81% and increase proline content from 15%-389%.
Key words Hevea brasiliensis Müll. Arg.; Glyphosate; Morphology; Physiological indices
doi 10.3969/j.issn.1000-2561.2016.01.011
草甘膦(Glyphosate)是1971年由Monsanto公司开发的广谱除草剂,抑制植物莽草酸合酶活性[1]。草甘膦是世界上应用最广、使用量最大的农药品种,大量用于农田各种杂草的防除,也可用于非农田杂草的治理,如园林、苗圃、铁路、公路、森林、湖泊等[2]。为全面了解草甘膦在环境中的毒性以便及时对可能的风险进行管理,国内外学者系统研究了草甘膦对大豆根瘤菌[3-4]、水生生物、微生物[5]、鱼类[6]、土壤[7]、蚯蚓[8]和人类[9]的危害。研究发现,草甘膦会引起棉花花药中植物激素和游离氨基酸的含量变化[10]。半致死量的草甘膦会导致油菜花变型和雄性不育[11],会影响大豆的种子成份[4]和根瘤菌的微生物构成[3]。因此,研究草甘膦药害机制和研发抗草甘膦作物品种是农业生产中的热点[12]。
橡胶树在育苗过程中,需要对苗床的杂草进行清除。在此过程中,少量的草甘膦会沾染橡胶树叶片,导致药害的产生。通常,叶片扭曲会导致叶绿体结构、活性氧合成、代谢和渗透调节物质含量的变化[13-15],并与生长素含量变化有关[16]。笔者推测,草甘膦药害会导致橡胶树叶片生理活性的变化。为了证明这一假设,笔者采用橡胶树CATAS7-33-97芽接苗为材料,对草甘膦处理后不同长度的叶片表型和生理活性進行测定,旨在阐明橡胶树草甘膦药害的生理机制,并为研究橡胶树对草甘膦的抗药机制和技术奠定基础。
1 材料与方法
1.1 材料
以中国热带农业科学院橡胶研究所培育的橡胶树品种热研7-33-97芽接苗为研究材料,芽接苗种植于中国热带农业科学院橡胶研究所实验基地草炭土与粘土比例为3 ∶ 1的育苗袋中。
1.2 方法
1.2.1 草甘膦喷施 采用浙江绿野农化生物科技有限公司41%草甘膦水剂稀释成1 ∶ 2 400,V ∶ V溶液喷施热研7-33-97芽接苗第二蓬叶后,每批处理6株,每株取叶柄中间的一片叶片进行分析。待测叶片分为正常叶(以正常叶为对照叶片)、畸形叶(按照叶片长度细分为7~10 cm、10~20 cm、20~30 cm和30 cm以上4个级别)和新生的恢复叶共6个类型。
1.2.2 生理指标测定 叶片叶绿素含量和相对含水量的测定参照文献[17]的方法。含水量、游离脯氨酸、可溶性糖含量、超氧化物歧化酶(SOD)酶活性和过氧化物酶(POD)含量的测定参照文献[18]的方法。
1.3 统计分析
实验测定为3批次重复。采用IBM-SPSS 21.0对数据进行单因素方差分析和多重比较分析,采用OriginPro 9.2软件进行绘图。
2 结果与分析
2.1 草甘膦对橡胶树芽接苗叶片形态的影响
从图1可以看出,在橡胶树幼苗苗床中喷施草甘膦除草剂之后,7~10 d后会导致橡胶树幼苗叶片脱落。叶片脱落7~10 d后,落叶的枝条会长出新的变形并扭曲的叶片,长度7 cm以上,最长可达35 cm以上,远高于正常叶片(图1-A)。第二蓬畸形叶到稳定期后,新生的第三蓬叶叶片形状会有所恢复,但仍比正常叶片长(图1-B)。这说明草甘膦会导致橡胶树幼苗叶片产生特异表型。
2.2 草甘膦对橡胶树芽接苗不同类型叶片生理活性的影响
2.2.1 草甘膦对橡胶树芽接苗叶片含水量的影响
由于叶片扭曲通常与叶片失水有关,筆者测定了6种叶片含水量变化规律。从图2可以看出,在橡胶树幼苗苗床中喷施草甘膦除草剂之后,正常叶、7~10 cm、10~20 cm、20~30 cm和30 cm以上4个级别畸形叶和恢复叶的含水量分别为69.49%、68.54%、62.28%、63.51%、63.27%、62.18%。正常叶、7~10 cm、10~20 cm、20~30 cm和30 cm以上4个级别畸形叶和恢复叶的相对含水量分别为92.68%、89.26%、91.75%、94.36%、80.24%、80.62%。这6种类型叶片含水量影响差异并不显著,但相对含水量存在显著差异,30 cm以上畸形叶和恢复叶的相对含水量较低。可见,草甘膦诱导叶片表型变化与一般性缺水引起叶片皱缩的机制不同。
2.2.2 草甘膦对橡胶树芽接苗叶片叶绿素含量的影响
为了探求叶片扭曲对叶绿体结构和功能的影响,测定了6种叶片叶绿素含量变化规律。从图3可以看出,正常叶、7~10 cm、10~20 cm、20~30 cm和30 cm以上畸形叶和恢复叶的叶绿素a含量分别为2.92、2.16、2.46、2.70、2.07、0.95 mg/g鲜重;叶绿素b含量分别为0.58、0.54、0.61、0.60、0.41、0.13 mg/g鲜重;β-胡萝卜素含量分别为0.81、0.66、0.70、0.81、0.62、0.39 mg/g鲜重。由于6种类型叶片叶绿素a和b含量存在显著差异,总叶绿素含量和叶绿素a/b比值也存在显著差异。由于Chla多位于叶片叶绿体类囊体膜PSII反应中心蛋白复合体,而Chlb多位于叶片叶绿体类囊体膜捕光色素天线蛋白复合体,二者的比值反映两类蛋白复合体的含量变化。因此,叶绿素a和b含量和比值的变化说明叶片叶绿体类囊体膜结构的变化。
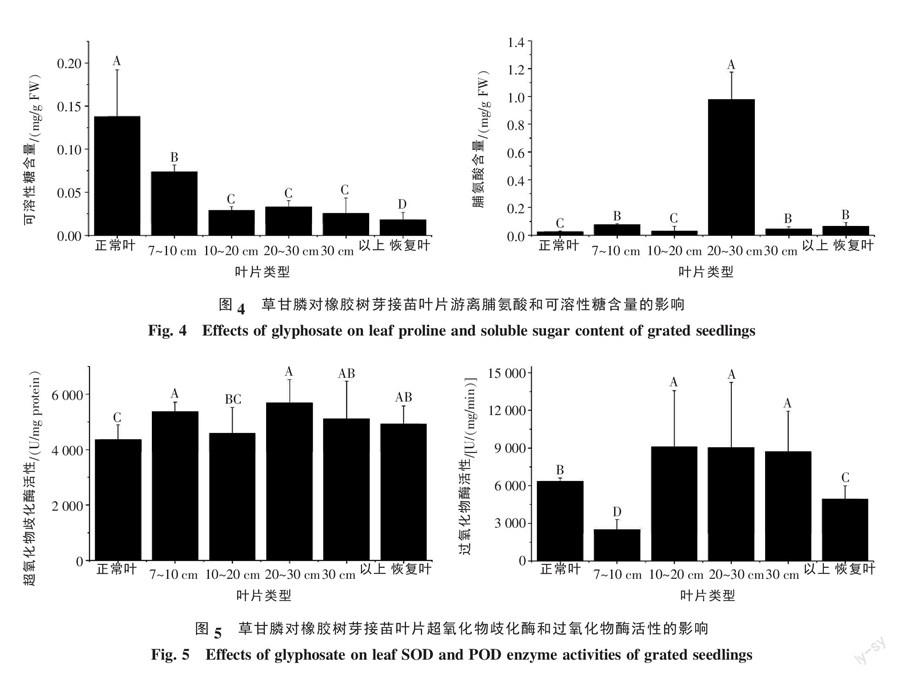
2.2.3 草甘膦对橡胶树芽接苗叶片游离脯氨酸和可溶性糖含量影响 从图4可以看出,正常叶、7~10 cm、10~20 cm、20~30 cm和30 cm以上畸形叶和恢复叶的可溶性糖含量分别为0.138、0.074、0.029、0.033、0.025、0.018 mg/g鲜重;脯氨酸含量分别为0.025、0.075、0.029、0.978、0.045、0.065 mg/g鲜重。6种类型叶片脯氨酸和可溶性糖含量均存在显著差异,畸形叶和恢复叶可溶性糖含量显著低于对照叶片,而脯氨酸含量高于对照叶片。说明草甘膦影响了不同类型叶片中渗透调节物质中脯氨酸和可溶性糖的含量。
2.2.4 草甘膦对橡胶树芽接苗叶片过氧化物酶和超氧化物歧化酶活性的影响 从图5可以看出,正常叶、7~10 cm、10~20 cm、20~30 cm和30 cm以上畸形叶和恢复叶的超氧化物歧化酶活性分别为4 359.66、5 366.80、4 582.99、5 684.86、5 108.53、4 928.34 U/mg蛋白;过氧化物酶活性分别为6 355.87、2 512.13、9 104.63、9 036.31、8 711.99、4 924.05 U/(mg·min)。6种类型叶片超氧化物歧化酶和过氧化物酶活性均存在显著差异,畸形叶和恢复叶超氧化物歧化酶活性高于正常叶片,10~20 cm、20~30 cm和30 cm以上畸形叶片过氧化物酶活性高于正常叶片。说明叶片扭曲导致活性氧的产生和活性氧淬灭酶活性的提高。
3 讨论与结论
鉴于草甘膦的广泛应用,其潜在的危害一直受到关注[2,19-21]。另一方面,通过研究草甘膦的作用机制,采用分子技术研发抗草甘膦除草剂的作物新品种是农业生产中的热点[22-23]。随着我国橡胶树种植规模的扩大、胶园更新和新品种的推广,橡胶树育苗工作大规模开展。但是,草甘膦除草剂对橡胶树幼苗的影响仍不清楚。笔者发现草甘膦诱导橡胶树幼苗叶片新生叶畸形的特点有助于阐明橡胶树叶片的草甘膦抗性机制,并培育抗草甘膦除草剂的橡胶树品种打下基础。
叶片扭曲通常认为与叶片含水量、活性氧代谢和光合作用变化有关。水稻卷叶突变体卷曲叶片会导致丙二醛含量上升和叶绿素含量降低[24]。半卷叶水稻突变体ATP合酶活性增加[25]。草甘膦药害会导致水稻穗部畸形和不育[26]。可见,植物叶片卷曲会导致生理功能发生巨大变化,这与本研究结果一致(图1)然而,笔者发现草甘膦喷施后橡胶树畸形叶片与正常叶的叶片含水量差异不显著(图2),说明叶片扭曲不是由于叶片脱水引起的,而是内部生理结构发生变化。首先,叶片扭曲导致叶绿体结构发生改变。笔者发现,畸形叶叶绿素a含量下降显著,表明光系统II反应中心的蛋白含量降低,叶绿素b含量降低较慢,表明捕光天线系统受到的影响较小(图3)。说明橡胶树畸形叶叶绿体类囊体膜捕光天线上叶绿素b吸收的过多的激发能不能通过光系统II反应中心的叶绿素a传递用于光化学淬灭,多余的激发能导致活性氧含量的上升。因此,笔者发现活性氧淬灭的关键酶过氧化物酶和超氧化物歧化酶歧化酶活性均显著高于正常叶(图5)。
由于草甘膦的靶标是莽草酸合酶,施用草甘膦会引起植物细胞内部的蛋白含量变化。例如,草甘膦会引起胡萝卜细胞中游离氨基酸含量上升,蛋白种类、功能变化[27-28],蛋白含量下降[29]。笔者亦发现草甘膦处理叶片中游离脯氨酸含量上升(图4)。畸形叶可溶性糖含量的下降与光合活性的减少有关。说明叶片扭曲影响光合作用和羧化作用。这可能是由生长素合成和运输紊乱引起的[30],也可能与乙烯生成有关[31]。笔者拟进一步采用高效液相色谱法测定畸形叶叶片激素含量,采用转录组测序技术筛选差异基因,结合本研究的结果,共同揭示草甘膦对橡胶树叶片的作用机制。
参考文献
[1] Amrhein N, Deus B, Gehrke P, et al. The site of the inhibition of the shikimate pathway by glyphosate: ii. interference of glyphosate with chorismate formation in vivo and in vitro[J]. Plant Physiology, 1980, 66(5): 830-834.
[2] 周垂帆, 李 莹, 张晓勇, 等. 草甘膦毒性研究进展[J]. 生态环境学报, 2013, 22(10): 1 737-1 743.
[3] Zobiole L H, Kremer R J, Oliveira R S, Jr., et al. Glyphosate affects micro-organisms in rhizospheres of glyphosate-resistant soybeans[J]. Journal of Applied Microbiology, 2011, 110(1): 118-127.
[4] Zobiole L H, Oliveira R S, Visentainer J V, et al. Glyphosate affects seed composition in glyphosate-resistant soybean[J]. Journal of Agricultural and Food Chemistry, 2010, 58(7): 4 517-4 522.
[5] 呼 蕾, 和文祥, 高亞军. 草甘膦对土壤微生物量及呼吸强度的影响[J]. 西北农业学报, 2010, 19(7): 168-172.
[6] Yusof S, Ismail A, Alias M S. Effect of glyphosate-based herbicide on early life stages of Java medaka(Oryzias javanicus): A potential tropical test fish[J]. Marine Pollution Bulletin, 2014, 85(2): 494-498.
[7] Khan S U, Young J C. N-nitrosamine formation in soil from the herbicide glyphosate[J]. Journal of Agricultural and Food Chemistry, 1977, 25(6): 1 430-1 432.
[8] Zhou C F, Wang Y J, Sun R J, et al. Inhibition effect of glyphosate on the acute and subacute toxicity of cadmium to earthworm Eisenia fetida[J]. Environmental Toxicology and Chemistry / SETAC, 2014, 33(10): 2 351-2 357.
[9] Zouaoui K, Dulaurent S, Gaulier J M, et al. Determination of glyphosate and AMPA in blood and urine from humans: about 13 cases of acute intoxication[J]. Forensic Science International, 2013, 226(1-3): e20-25.
[10] 刘吉焘, 狄佳春, 陈旭升. 草甘膦诱导抗草甘膦棉花花药中激素和游离氨基酸含量的变化[J]. 分子植物育种, 2014, 12(3): 530-536.
[11] Londo J P, McKinney J, Schwartz M, et al. Sub-lethal glyphosate exposure alters flowering phenology and causes transient male-sterility in Brassica spp[J]. BMC Plant Biology, 2014, 14: 70.
[12] Wang W, Xia H, Yang X, et al. A novel 5-enolpyruvoylshikimate-3-phosphate(EPSP)synthase transgene for glyphosate resistance stimulates growth and fecundity in weedy rice(Oryza sativa)without herbicide[J]. The New Phytologist, 2014, 202(2): 679-688.
[13] Zwieniecki M A, Boyce C K, Holbrook N M. Functional design space of single-veined leaves: role of tissue hydraulic properties in constraining leaf size and shape[J]. Annals of Botany, 2004, 94(4): 507-513.
[14] Shi Z, Wang J, Wan X, et al. Over-expression of rice OsAGO7 gene induces upward curling of the leaf blade that enhanced erect-leaf habit[J]. Planta, 2007, 226(1): 99-108.
[15] Wu C, Fu Y, Hu G, et al. Isolation and characterization of a rice mutant with narrow and rolled leaves[J]. Planta, 2010, 232(2): 313-324.
[16] Zgurski J M, Sharma R, Bolokoski D A, et al. Asymmetric auxin response precedes asymmetric growth and differentiation of asymmetric leaf1 and asymmetric leaf2 Arabidopsis leaves[J]. The Plant Cell, 2005, 17(1): 77-91.
[17] Wang L F, Wang M, Zhang Y. Effects of powdery mildew infection on chloroplast and mitochondrial functions in rubber tree[J]. Tropical Plant Pathology, 2014, 39(3): 242-250.
[18] 中国科学院上海植物生理研究所, 上海市植物生理学会. 现代植物生理学实验指南[M]. 北京: 科学出版社, 1999.
[19] 杨治峰, 张振玲. 草甘膦生殖发育毒性的研究进展[J]. 环境与职业医学, 2013, 30(2): 154-156.
[20] 傅建炜, 史梦竹, 李建宇, 等. 草甘膦对草鱼、 鲢鱼和鲫鱼的毒性[J]. 生物安全学报, 2013, 22(2): 119-122.
[21] 窦建瑞, 钱晓勤, 毛一扬, 等. 草甘膦对人体的毒性研究进展[J]. 江苏预防医学, 2013, 24(6): 43-45.
[22] 朱 玉, 于中连, 林 敏. 草甘膦生物抗性和生物降解及其转基因研究[J]. 分子植物育种, 2003, 1(4): 435-441.
[23] 董合忠, 代建龙. 转基因抗草甘膦棉花及其对草甘膦抗性的时空表达[J]. 中国农学通报, 2007, 23(2): 355-359.
[24] Wang L F, Fu H, Ji Y H. Photosynthetic characterization of a rolled leaf mutant of rice(Oryza sativa L.)[J]. African Journal of Biotechnology, 2012, 11(26): 6 839-6 846.
[25] Xiang J J, Zhang G H, Qian Q, et al. Semi-rolled leaf1 encodes a putative glycosylphosphatidylinositol-anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells[J]. Plant Physiology, 2012, 159(4): 1 488-1 500.
[26] 何国发. 草甘膦造成水稻药害的症状及补救措施[J]. 现代农业科技, 2011(8): 179.
[27] Killmer J, Widholm J, Slife F. Reversal of glyphosate inhibition of carrot cell culture growth by glycolytic intermediates and organic and amino acids[J]. Plant Physiology, 1981, 68(6): 1 299-1 302.
[28] Rubin J L, Gaines C G, Jensen RA. Enzymological basis for herbicidal action of glyphosate[J]. Plant Physiology, 1982, 70(3): 833-839.
[29] Haderlie L C, Widholm J M, Slife F W. Effect of glyphosate on carrot and tobacco cells[J]. Plant Physiology, 1977, 60(1): 40-43.
[30] Baur J R. Effect of glyphosate on auxin transport in corn and cotton tissues[J]. Plant Physiology, 1979, 63(5): 882-886.
[31] Lee T T, Dumas T. Effect of glyphosate on ethylene production in tobacco callus[J]. Plant Physiology, 1983, 72(3): 855-857.
責任编辑:白 净
