黄淮麦区部分小麦和国外引进小麦GS2等位基因的检测及其与农艺性状的关联分析
2016-05-27宋晓朋王宇娟武炳瑾马文洁张德强周丽敏孙道杰
宋晓朋,王宇娟,武炳瑾,马文洁,张德强,周丽敏,孙道杰
(1.西北农林科技大学农学院,陕西杨凌 712100;2.杨凌区种子站,陕西杨凌 712100)
黄淮麦区部分小麦和国外引进小麦GS2等位基因的检测及其与农艺性状的关联分析
宋晓朋1,王宇娟2,武炳瑾1,马文洁1,张德强1,周丽敏1,孙道杰1
(1.西北农林科技大学农学院,陕西杨凌 712100;2.杨凌区种子站,陕西杨凌 712100)
摘要:为明确黄淮麦区 TaGS2等位基因的分布状况及其与主要农艺性状的关系,对黄淮麦区2008年之前育成的种质材料、新育成的品种(系)及国外引进材料,用 TaGS2-A1、 TaGS2-B1和 TaGS2-D1等功能标记鉴定对应的基因,并结合相关农艺性状发掘优势单倍型。结果表明,黄淮麦区2008年之前育成的种质材料和新育成品种(系)中 TaGS2等位基因分布频率存在一定的差异; TaGS2-A1b、 TaGS2-B1b和 TaGS2-D1a是优势 TaGS2等位基因, TaGS2-A1b在小麦抽穗期、株高和小穗数的改良上是优势单倍型,但在新育成的品种(系)中有下降的趋势, TaGS2-D1a能够显著增加小穗数、穗粒数和穗粒重,在各类材料中的比例都较高; TaGS2-B1b是提高千粒重的优势单倍型。因此,在黄淮麦区小麦穗部性状改良中 TaGS2-B1b和 TaGS2-D1a的作用显著,尤其是 TaGS2-D1a,同时黄淮麦区种植小麦遗传多样性在减少,一些优势单倍型未受到重视,应对地方品种和一些国外引进材料加以利用。
关键词:小麦; TaGS2;功能标记;单倍型;关联分析
氮是维持小麦高产稳产需求量的重要元素。多年来,氮肥的过量施用在满足小麦生长发育获得高产的同时也造成了严重环境污染。因此,在保障小麦高产优质的同时,如何提高氮素利用效率就显得非常重要。发掘和利用与小麦高产和氮素利用相关的基因是促进氮素高效利用的重要途径。谷氨酰胺合成酶(GS)是高等植物氮代谢的关键酶[1],对氨基酸的初步合成和来自光呼吸过程中氮的再动员起着关键作用[2]。GS有两种同工酶,分别是胞液型GS1和质体型GS2,两者具有不同的生理功能[3-4]。GS基因对小麦氮素利用效率和农艺性状的影响已经有很多学者进行了研究。在小麦中过量表达GS1基因可使小麦籽粒产量提高了20%[5];在后来的研究中Habash[6]发现,GS活性与小麦叶片可溶性蛋白和氮含量呈正相关,并且通过构建中国春与SQ1的QTL作图群体发现控制小麦叶片可溶性蛋白、氮含量和产量性状的QTLs与2AS上GS2的QTL位点重合;Gadaleta等[7]通过对硬粒小麦GS2基因的研究发现,位于2A和2B上的两个GS2基因与小麦籽粒蛋白质含量相关的QTLs处于同一区域。这些研究虽然从不同方面反映出GS2是一个与小麦氮素利用相关的基因,但是很少有学者对GS2基因在不同小麦种质资源中的分布进行探究。Li等[8]从中国春和小偃54基因组DNA序列中鉴定出3种TaGS2序列,通过序列比对分析发现了多个单倍型,并通过对一些小麦材料和性状的关联分析发现TaGS2-A1b、TaGS2-B1a、TaGS2-B1b和TaGS2-D1a对小麦产量和氮素利用效率的提高具有显著作用。但TaGS2基因在黄淮麦区小麦品种(系)中的分布如何、与TaGS2有关的功能标记能否对该区的种质材料进行检测以及与哪些农艺性状存在关联,这些问题目前都尚不清楚。基于此,本研究通过搜集黄淮麦区小麦骨干亲本和优异的种质资源对TaGS2等位基因进行鉴定,发掘优异的等位变异,以期为黄淮麦区小麦高产和氮高效育种提供检测依据和一定的技术支撑。
1材料与方法
1.1材 料
供试材料有2008之前育成154份黄淮麦区种质材料、新育成的品种(系)(包括53份2014-2015年国家黄淮南片区试品系和71份2014-2015年度陕西省区试材料)及57份国外引进材料。
1.2方 法
1.2.1田间种植和农艺性状调查
124份区试材料于2014年10月初种植于杨凌国家区试试验站,按照小麦国家区试标准进行种植;154份黄淮麦区材料和57份国外引进材料于2013-2015种植于西北农林科技大学实验基地,并按单因素随机区组设置试验,每个材料种植2行,行长2 m,行距0.25 m,株距2 cm,三个重复。154份黄淮麦区种质材料调查的主要农艺性状有抽穗期(从播种到抽穗的天数)、株高、小穗数、穗粒数、小穗密度、穗粒重、穗下节直径和厚度、不育小穗数、粒重等。
1.2.2基因组DNA提取
在小麦苗期,选取幼嫩的叶片按照改良CTAB法[9-10]提取供试材料中小麦基因组DNA。将DNA溶于TE溶液中,利用0.8%的琼脂糖凝胶电泳检测DNA的完整性,并用紫外分光光度计检测其浓度。
1.2.3TaGS2功能标记的检测
试验所用的功能标记引物参照李新鹏等[8]所公布的引物序列(表1),由上海生工生物工程有限公司合成。PCR反应体系20 μL,包括2×Mix混合液10 μL,上下游引物各1 μL(5 μmol·L-1),模板DNA 2 μL(200 ng·μL),ddH2O 6 μL。PCR反应程序:94 ℃预变性5 min,94 ℃变性40 s,退火40 s(具体标记退火温度见表1),72 ℃延伸1 min,30个循环,72 ℃延伸10 min。TaGS2-B1的两个功能标记的PCR产物均由6%的变性聚丙酰胺凝胶电泳检测,TaGS-A1和TaGS-D1功能标记扩增的产物用1.6%的琼脂糖凝胶电泳检测,并按序列间的多态性统计单倍型。
1.3数据分析
用Excel 2010对数据进行整理,用SPSS 20.0对基因型和表型性状进行t测验和关联分析。
2结果与分析
2.1TaGS2-A1基因在供试小麦材料中的分布
在供试小麦材料中,没有发现TaGS2-A1新的等位变异(图1和表2)。在335份小麦材料中,151份材料扩增出与TaGS2-A1a带型相同的片段,占供试材料的45%,而181份材料检测出TaGS2-A1b,占供试材料的55%。在2008年之前黄淮麦区育成的种质材料(154份)中,66份(43%)材料含有TaGS2-A1a带型,88份(57%)材料包含有TaGS2-A1b带型;2014-2015年度国家黄淮麦区南片区试材料中,只有23份(43%)材料检测出TaGS2-A1b,而检测出TaGS2-A1a带型的材料却有30份(57%),TaGS-A1等位变异在两个不同时期的小麦材料中的分布存在很大的差异; 但是从2014-2015年度陕西省区试材料(71份)中仍能够检测出27份(38%)包含有TaGS2-A1a,44份(62%)携带有TaGS2-A1b,这与2008前黄淮麦区种质材料中TaGS2-A1基因型分布较为一致;从国外引进的种质资源(57份)中有28份材料检测出TaGS2-A1a,29份材料检测出TaGS2-A1b,TaGS2-A1两种基因型在国外引进材料中的分布没有明显的差异。

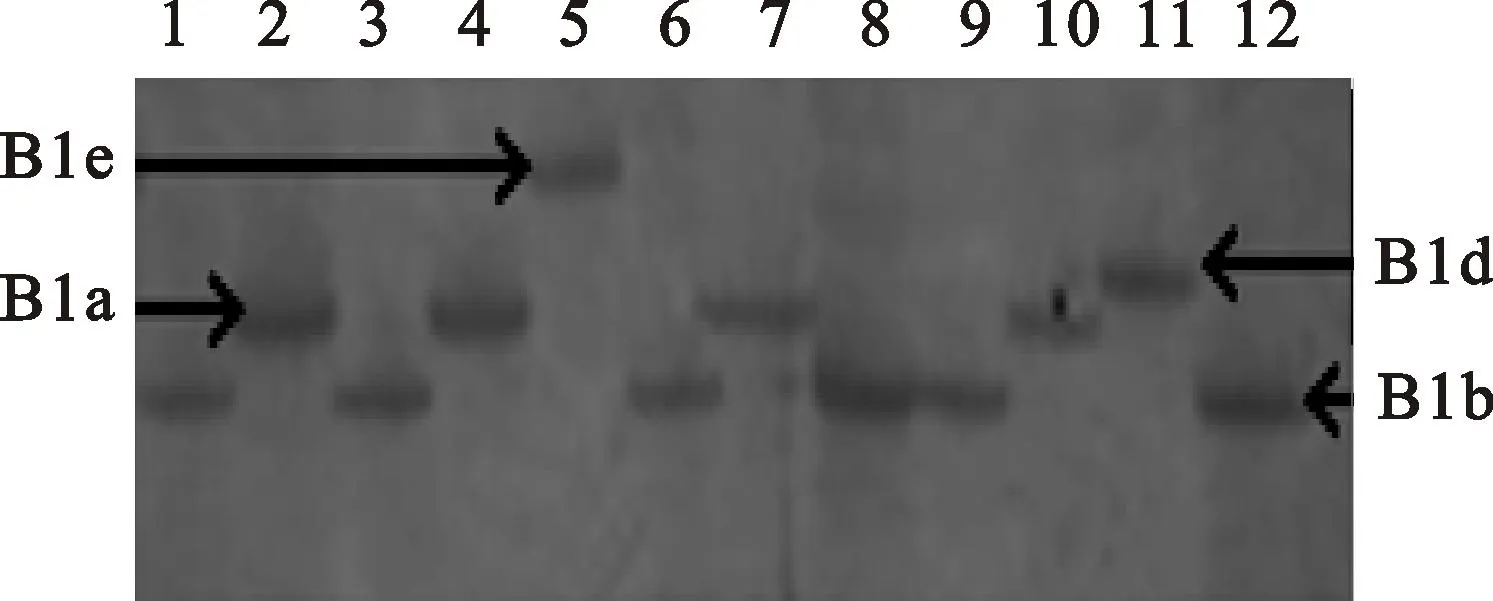
M:DL2000;1:碧蚂1号;2:偃展4110;3:周8425B;4:西农1376;5:郑麦9023;6:小偃22;7:周麦22;8:郑州3号;9:PH 82-2;10:郑引1号;11郑麦366;12:矮抗58
M:DL2000; 1:Bima 1; 2:Yanzhan 4110; 3:Zhou 8425B; 4:Xinong 1376; 5:Zhengmai 9023; 6:Xiaoyan 22; 7:Zhoumai 22; 8:Zhengzhou 3; 9:PH 82-2; 10:Zhengyin 1; 11:Zhengmai 366; 12:Aikang 58
图1功能标记TaGS2-A1扩增出的片段类型
Fig.1PCR fragments amplified with the
function markerTaGS2-A1
2.2TaGS2-B1基因在供试小麦材料中的分布
在供试材料中,有5种TaGS2-B1基因型存在,没有发现其他新的等位变异(图2和表2)。在供试材料TaGS2-B1等位变异中,TaGS2-B1a和TaGS2-B1b是主要的单倍型。TaGS2-B1a在2008年之前育成的黄淮麦区材料中检测出54份(35%),在2014-2015年陕西省区试材料和外国引进材料中分别检测出17份(24%)和16份(28%),而在2014-2015年黄淮南片区试材料中却检测出28份(53%);在2008年之前育成的黄淮麦区材料中TaGS2-B1b单倍型的比例(55%)比2014-2015年度黄淮麦区南片区试材料中的比例(47%)要高;在供试材料中区TaGS2-B1d的比例(9%)也很大,但在2008年之前黄淮麦区材料中TaGS2-B1d只有4份(3%),在2014-2015年度黄淮麦区南片区试材料中却没有检测出,而在2014-2015年度陕西省小麦区试材料中TaGS2-B1d基因型有8份(14%),在国外引进材料中则有更多的TaGS2-B1d基因型(18份,占国外引进材料的32%)。因此,在供试材料中TaGS2-B1a、TaGS2-B1b和TaGS2-B1d单倍型的分布呈现出很大变化。在TaGS2-B1等位变异中,TaGS2-B1e、TaGS2-B1f是2种稀有单倍型,在2014-2015年度黄淮麦区南片区试材料中却未被检测出。

表2 TaGS2等位基因在供试材料中的分布
Ⅰ:黄淮麦区生产中正在应用的小麦种质(2008年之育成);Ⅱ:2014-2015年度黄淮麦区南片区试材料;Ⅲ:2014-2015年度陕西省区试材料;Ⅳ:国外引进材料
I:Wheat varieties from Huang-Huai wheat region before 2008; Ⅱ:Wheat varieties from Huang-Huai southern wheat regional trial between 2014 and 2015; Ⅲ:Wheat varieties from Shaanxi province regional trial between 2014 and 2015; Ⅳ:Introduced varieties
2.3TaGS2-D1基因在供试小麦材料中的分布
在TaGS2-D1等位变异(图3和表2)中,TaGS2-D1a不仅在供试材料中占据较高的比例(234份,占供试材料的70%),而且在不同材料中的比例也较高;TaGS2-D1b在国外引进材料和2008年前黄淮麦区材料中分别检测出22份(占39%)和52份(占34%),但在2014-2015年度黄淮南片区试(12份,占23%)和陕西省区试材料(15份,占21%)的比例较低,表明TaGS2-D1a在小麦种质材料中是优势单倍型。从2014-2015年度黄淮南片区试、陕西省区试和2008年前黄淮麦区材料中TaGS2-D1a基因型分布可以看出,该基因型在黄淮麦区材料中的比例在增加。
2.4TaGS2基因等位变异与小麦农艺性状的关系
通过对154份黄淮麦区小麦材料农艺性状与TaGS2基因关联分析,TaGS2-A1a和TaGS2-A1b基因型间、TaGS2-D1a和TaGS2-D1b基因型间抽穗期差异显著或极显著;TaGS2-A1a和TaGS2-A1b基因型间株高、小穗数和穗下节直径差异显著,而小穗密度、穗粒重、穗粒数等性状不存在显著差异。对TaGS2-D1两种等位变异关联分析发现,TaGS2-D1a和TaGS2-D1b两种基因型间小穗数、小穗密度、穗粒数、穗粒重等穗部性状存在显著差异,而株高和穗下节直径差异不显著。此外,TaGS2-B1两种主要的基因型TaGS2-B1a和TaGS2-B1b间千粒重差异显著,而其他农艺性状没有显著性差异(表3)。可见,在黄淮麦区中,TaGS2-A1b和TaGS2-D1a基因型的材料能够提前抽穗,TaGS2-A1b单倍型能够降低株高和增加穗下节直径,TaGS2-D1a能够增加小穗数、穗粒数和穗粒重,在这些性状上TaGS2-A1b和TaGS2-D1a是优势的基因型,而TaGS2-B1b单倍型对千粒重的提高具有显著作用,是优势等位变异。
3讨 论
小麦是一个异源六倍体作物,基因组大,重复序列高,对单个基因位点的检测难度较大,但小麦不仅A、B和D三个基因组之间有着非常好的共线性[11],而且与水稻、短柄草等存在一定共线性[12-13],这就使得小麦基因研究结果能够得到验证,可以提高结果的可信度。本研究中,经表型性状的关联分析,TaGS2-A1和TaGS2-D1基因型间抽穗期存在显著差异,表明在两个基因附近存在着控制抽穗期的基因或QTL。在水稻中也发现GS2位点附近有叶片衰老[14]、抽穗时间[15]等性状的QTL位点与GS2连锁。株高和千粒重是小麦育种中两个重要农艺性状。本研究表明,TaGS2-A1b能够显著降低株高,尽管现在还没有相关的文献来验证这一结果,但是我们推测在TaGS2-A1基因附近可能存在与株高相关的QTL位点;仅在TaGS2-B1基因型之间检测到千粒重存在显著差异,这与Li等[15]的研究结果有所不同,这可能与试验材料不同有关;在TaGS2-D1和TaGS2-A1基因型之间也检测到了小穗数差异显著,这与Ma等[16]利用“南大2419×望水白”分离群体发现一个与控制小穗数QTL连锁的标记的研究结果较为一致。小穗数、小穗密度和穗粒重在不同TaGS2基因型之间存在显著差异,而Peng等[17]也在这一区间内发现小穗数、穗粒数、千粒重等性状相关的标记,表明这一区段在小麦遗传改良中具有重要的研究价值,应该给予重视。
1:周麦31;2:碧蚂1号;3:中麦895;4豫麦47;5:豫麦50;6:郑麦9023;7:郑麦366;8:内乡188;9:周麦30;10:偃展4110;11:豫麦34;12:小偃81
1:Zhouma 31; 2:Bima 1; 3:Zhongmai 895; 4:Yumai 47; 5:Yumai 50; 6:Zhengmai 9023; 7:Zhengmai 366; 8:Neixiang 188; 9:Zhoumai 30; 10:Yanzhan 4110; 11:Yumai 34; 12:Xiaoyan 81
图2功能标记TaGS2-B1扩增
出片段的部分类型
Fig.2PCR fragments amplified with the
function markerTaGS2-B1

M:DL2000;1:陕麦509;2:中麦895;3:小偃54;4:西农1376;5:碧蚂1号;6:豫麦34;7:周8425B;8:小偃81;9;周麦22;10郑引1号;11:矮抗58;12:PH 82-2
M:DL2000; 1:Shaanmai 509; 2:Zhongmai 895; 3:Xiaoyan 54; 4:Xinong 1376; 5:Bima 1; 6:Yumai 34; 7:Zhou 8425B; 8:Xiaoyan 81; 9:Zhoumai 22; 10:Zhengyin 1; 11:Aikang 58; 12:PH 82-2
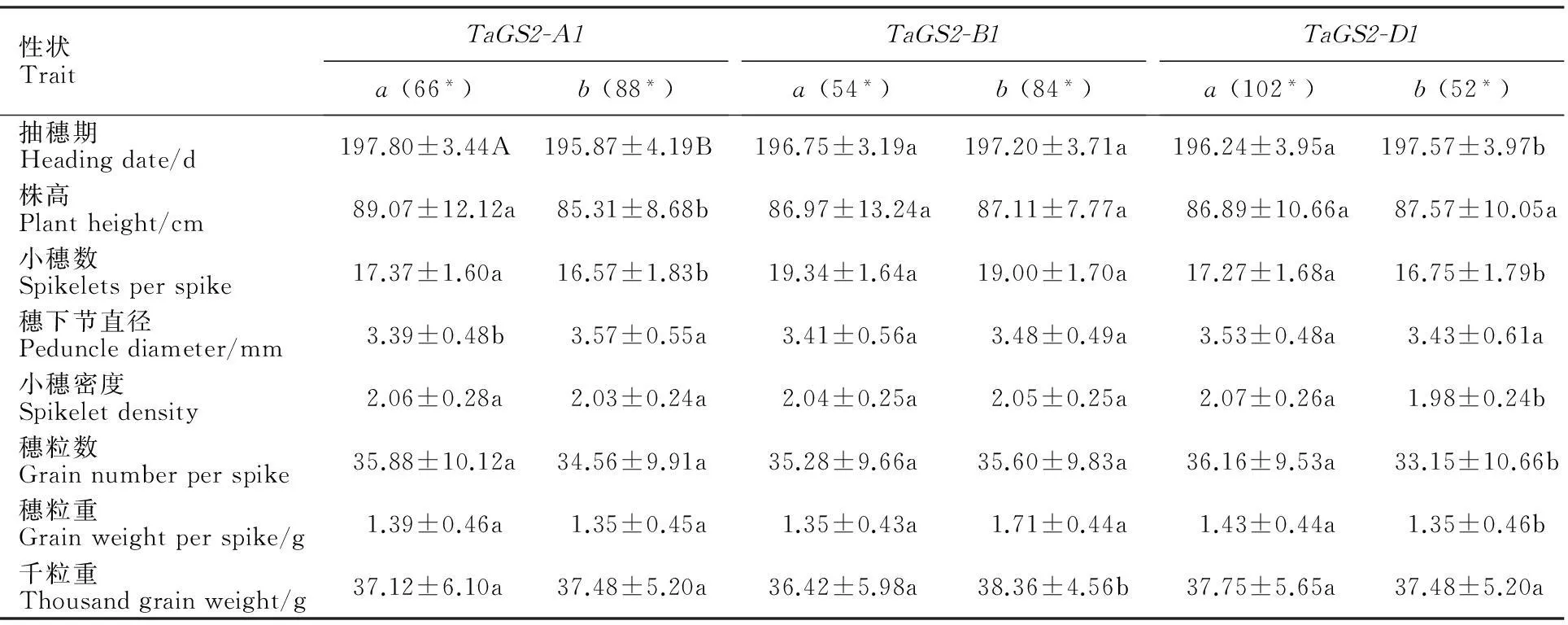
图3 功能标记 TaGS2-D1扩增出的片段类型
表型值中,不同大写和小写字母分别表示不同单倍型在0.01和0.05水平下差异显著;*:品种数目
In phenotype values,capital and small letters designate significance between different haplotypes at 0.01 and 0.05 level,respectively; *:Number of accession
从不同年份供试材料TaGS2等位变异分布看,三个TaGS2基因单倍型的频率均出现不同程度的变化。TaGS2-D1a单倍型在不同年份材料中的比例在增加,而TaGS2-A1b和TaGS2-B1b单倍型在黄淮南片区试材料中的比例却在减少,但是在陕西省区试中却呈现增长的趋势,TaGS2-B1e和TaGS2-B1f是稀有的单倍型,其比例也在降低。对于TaGS2单倍型频率的变化,推测可能有几种原因:一是在试验材料选择中,2008年之前黄淮麦区的材料154份,而2014-2015年度的黄淮南片区试材料只有53份,材料人为选择可能导致基因型比例不同的一个重要原因;二是单倍型决定了性状,在育种过程中对性状的选择直接导致了单倍型频率的改变;三是选择牵连效应[18-19],由于单倍型并不导致性状的改变,但是与决定性状的基因存在连锁关系,育种中在选择决定性状基因的同时也选择了单倍型。从2014-2015年度陕西省区试和黄淮南片区试材料中,TaGS2等位基因型的分布并没有呈现一致性趋势,因此第二和三种推测出现的可能性较小,具体什么原因造成TaGS2基因型频率的改变,需要更深入的研究。从TaGS2等位变异频率看,各单倍型在国外引进材料和国内材料中的频率几乎有很大差异,尤其是TaGS2-B1d的频率,这样很可能是在小麦种质资源改良中,只关注国外材料中的一些个别性状如丰产性、抗病性等造成的。从供试材料TaGS2等位基因变异频率变化上来看,我国黄淮麦区小麦遗传多样正在逐步减少,因此有必要对地方品种和国外引进材料进行重视和加以利用。
参考文献:
[1]Bernard S M,Habash D Z.The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling [J].NewPhytologist,2009,182:608-620.
[2]Edwards J W,Coruzzi G M.Photorespiration and light act in concert to regulate the expression of the nuclear gene for chloroplast glutamine synthetase [J].PlantCell,1989,1:241-248.
[3]Yoshie S,Noda N,Tsuneda S.Salinity decreases nitrite reductase gene diversity in denitrifying bacteria of wastewater treatment systems [J].AppliedandEnvironmentalMicrobiology,2004,70(5):3152-3157.
[4]Miflin B J.The location of nitrite reductase and other enzymes related to amino acid biosynthesis in the plastids of root and leaves [J].PlantPhysiology,1974,54(4):550-555.
[5]Habash D Z,Massiah A J,Rong H L,etal.The role of cytosolic glutamine synthetase in wheat [J].AnnalsofAppliedBiology,2001,138:83-89.
[6]Habash D Z,Bernard S,Schondelmaier J,etal.The genetics of nitrogen use in hexaploid wheat:nutilisation,development and yield [J].TheoreticalandAppliedGenetics,2007,114:403-419.
[7]Gadaleta A,Nigro D,Giancaspro A,etal.The glutamine synthetase (GS2) genes in relation to grain protein content of durum wheat [J].Functional&IntegrativeGenomics,2011,11:665-670.
[8]Li X P,Zhao X Q,He X,etal.Haplotype analysis of the genes encoding glutamine synthetase plastic isoforms and their association with nitrogen-use and yield-related traits in bread wheat [J].NewPhytologist,2011,189:449-458.
[9]王关林,方宏筠.植物基因工程[M].北京:科学出版社,2005:533-535.
Wang G L,Fang H J.Plant Genetic Engineering [M].Beijing:Science Press,2005:533-535.
[10]Wang M L,Barkley N A,Yu J K,etal.Transfer of simple sequence repeat (SSR) marker from major cereal crops to minor grass species for germplasm characterization and evaluation [J].PlantGeneticResources,2005,3(1):45-47.
[11]Dubcovsky J,Luo M C,Zhong G Y,etal.Genetic map of diploid wheat,TriticummonococcumL.,and its comparision with maps ofHordeumvulgareL.[J].Genetics,1996,143(2):983-999.
[12]Ahn S,Anderson J A,Sorrells M E,etal.Homoeologous relationships of rice,wheat and maize chromosomes [J].GeneticsandMolecularResearch,1993,241(56):483-490.
[13]Moore G,Foote T,Helentjaris T,etal.Was there a single ancestral cereal chromosome? [J].TrendsinGenetics,1995,11(3):81-82.
[14]Jiang G H,He Y Q,Xu C G,etal.The genetic basis of stay-green in rice analyzed in a population of doubled haploid lines derived from an indica by japonica cross [J].TheoreticalandAppliedGenetics,2004,108:688-698.
[15]Li J Z,He P,Zheng X W,etal.Identification and interaction analysis of six agronomic trait loci of rice based on a recombinant inbred population [J].ActaBotanySinica,1999,41:1199-1203.
[16]Ma Z,Zhao D,Zhang C,etal.Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2populations [J].GeneticsandMolecularResearch,2007,277:31-42.
[17]Peng J,Ronin Y,Fahima T,etal.Domestication quantitative trait loci inTriticumdicoccoides,the progenitor of wheat [J].ProceedingsoftheNationalAcademyofSciences,2003,100:2489-2494.
[18]Stephan W,Song Y S,Longley C H.The hitchhiking effect on linkage disequilibrium between linked neutral loci [J].Genetics,2005,172:2647-2663.
[19]张学勇,童依平,游光霞,等.选择牵连效应分析:发掘重要基因的新思路[J].中国农业科学,2006,39(8):1526-1535.
Zhang X Y,Tong Y P,You G X,etal.Hitchhiking effect mapping:a new approach for discovering agronomic important genes [J].ChinaAgricultureScience,2006,39(8):1526-1535.
Allelic Variations ofTaGS2Genes and Their Association with Agronomic Traits in Whaet Cultivars from Huang-Huai Wheat Region in China and Overseas
SONG Xiaopeng1,WANG Yujuan2,WU Bingjin1,MA Wenjie1,ZHANG Deqiang1,ZHOU Limin1,SUN Daojie1
(1.College of Agronomy,Northwest A&F University,Yangling,Shaanxi 712100,China;2.Yangling Seed Station,Yangling,Shaanxi 712100,China)
Abstract:Glutamine synthetase (GS) plays a key role in the nitrogen (N) use and yield potential of bread wheat. Investigating the haplotypes in TaGS2 genes and their associations with agronomic traits may be a useful approach for improving wheat yield and N-use efficiency. In this study,278 elite wheat varieties from Huang-Huai winter wheat region and 57 introduced varieties were used to discover the frequencies of TaGS2 haplotypes and the association between TaGS2 and agronomic traits,in addition,corresponding allelic variants were also identified using the function markers of TaGS2 genes. The results indicated that the frequency of allelic variation of TaGS2 genes detected were different between those wheat varieties,three favorable TaGS2 haplotypes ( TaGS2-A1b, TaGS2-B1b and TaGS2-D1a) were revealed,but the frequencies of TaGS2-A1b and TaGS2-B1b among varieties in different years presented a downward trend. TaGS2-A1b was considered as a potential haplotype for improving heading date,plant height and spikelet number per ear, TaGS2-D1a was associated with significantly higher values in spikelet number per ear,grain number per ear and grain weight per ear,while TaGS2-B1b was a potential superior haplotype for increasing 1 000-grain weight,nevertheless, TaGS2-D1a for improving spike traits may be more valued by many breeders. In the Huang-Huai wheat region,the genetic diversity of wheat varieties was reducing,so we should pay more attention to landrace and some introduced varieties.
Key words:Wheat; TaGS2; Function marker; Haplotype; Association analysis
中图分类号:S512.1;S330
文献标识码:A
文章编号:1009-1041(2016)03-0281-06
通讯作者:孙道杰(E-mail:chinawheat@hotmail.com)
基金项目:国家重点基础研究计划项目(2014CB138100);陕西省自然科学基金项目(2015JM3094);陕西省重点科技创新团队资助项目(2014KCT-25)
收稿日期:2015-09-14修回日期:2015-11-07
网络出版时间:2016-03-01
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160301.1338.008.html
第一作者E-mail:171302897@qq.com
