2种鼠尾草对NaCl胁迫的耐受性比较及其生理机制研究
2016-05-06梁春虹赵美棠魏宇昆黄艳波
许 华,梁春虹,赵美棠,魏宇昆,黄艳波
(1 北京师范大学珠海分校 工程技术学院,广东珠海 519087;2 上海辰山植物园,中国科学院上海辰山植物科学研究中心,上海 201602)
2种鼠尾草对NaCl胁迫的耐受性比较及其生理机制研究
许华1,梁春虹1,赵美棠1,魏宇昆2*,黄艳波2
(1 北京师范大学珠海分校 工程技术学院,广东珠海 519087;2 上海辰山植物园,中国科学院上海辰山植物科学研究中心,上海 201602)
摘要:以具有较高药用和观赏价值的美丽鼠尾草和贵州鼠尾草为实验材料,分析2种鼠尾草在NaCl(0、200、300、400、500、600 mmol·L(-1))胁迫下的生长、叶绿素含量、保护酶活性和有机渗透调节物质含量的变化,以明确2种鼠尾草对NaCl胁迫的耐受性差异及其生理机制。结果显示:(1)在实验NaCl浓度范围内,美丽鼠尾草的受害程度均高于贵州鼠尾草;(2)随着NaCl浓度的提高,贵州鼠尾草叶片叶绿素含量无显著变化,而美丽鼠尾草叶绿素含量逐渐显著降低;(3)当NaCl浓度从0增加到500 mmol·L(-1)时, 2种鼠尾草叶片的POD、CAT活性以及可溶性糖、可溶性蛋白质和脯氨酸含量逐渐升高,且美丽鼠尾草叶片的SOD活性也逐渐升高;(4)当NaCl浓度达到600 mmol·L(-1)时,美丽鼠尾草叶片可溶性糖、可溶性蛋白质和脯氨酸含量继续增加,SOD、POD和CAT活性开始降低但仍显著高于对照,而贵州鼠尾草叶片的POD和CAT活性继续增加,可溶性糖、可溶性蛋白质和脯氨酸含量开始降低但仍显著高于对照。研究表明,贵州鼠尾草在NaCl胁迫下具有较高的渗透调节物质含量,而且随着NaCl浓度的增加能够维持较高的保护酶活性,因此对NaCl胁迫的耐受性强于美丽鼠尾草。
关键词:NaCl胁迫;贵州鼠尾草;美丽鼠尾草;渗透调节物质;保护酶
近年来,土壤盐渍化逐渐成了生态环境和农业发展中的严峻问题[1]。植物在盐渍化环境中,生长缓慢,代谢活动被抑制,甚至会出现盐斑、叶片萎蔫,甚至死亡等现象[2]。据最新的研究,国内盐碱地总面积达到了 37.4×106hm2,仍然呈现逐渐增长的趋势[3]。目前,对土壤盐碱地改良的生物措施主要是筛选或培育一些具有耐盐和抗盐能力的植物,以及在盐碱土壤上引种和驯化有经济价值的盐生植物和耐盐植物。随着生物技术的发展,利用传统的杂交技术和遗传工程方法培育抗盐新品种和培育转抗盐基因植物也是一种土壤盐碱地改良的生物措施[4]。
美丽鼠尾草(Salviameiliensis)和贵州鼠尾草(S.cavaleriei)同属于唇形科(Labiatae)鼠尾草属(Salvia)荔枝草亚属(Subg.Sclarea)丹参组(Sect.Drymosphace)中的植物。美丽鼠尾草主要分布在安徽西部海拔 400~1 300 m的山坡、林下及草丛;贵州鼠尾草主要分布在湖北、广东、广西、四川、贵州海拔530~1 300 m的山坡、林下、水沟边[5]。美丽鼠尾草和贵州鼠尾草均是重要的中药材。美丽鼠尾草的干燥根在安徽地区被代作丹参入药[6]。中药丹参是丹参(Salviamiltiorrhiza)的干燥根及根茎,具有祛瘀止痛,活血通经,清心除烦的功效,以丹参组成的复方制剂在治疗心脑血管疾病方面疗效显著[7]。贵州鼠尾草是全草均可入药,用于治疗吐血,咳血,血痢,血崩,刀伤出血,月经过多,胃腕痛;外用于跌打损伤,疖肿[8]。丹酚酸B是丹参活血化瘀功效的主要成分,景鹏飞[9]研究表明贵州鼠尾草的丹酚酸B含量高于丹参。同时,在我们的前期引种、栽培和驯化研究中发现,2种鼠尾草还具有一定的观赏价值和较强的生态适应性。
目前,对美丽鼠尾草的研究报道仅在于其化学成分[10],贵州鼠尾草的研究报道仅在于其化学成分[11]和药理研究[12-13],而关于美丽鼠尾草和贵州鼠尾草的抗逆性以及对逆境的适应机制的研究尚未见报道。对于植物抗逆性包括抗盐性的生理机制,国内外学者做了大量的研究工作,发现植物的渗透调节能力和保护酶活性与植物的抗逆性有重要的关系[14-17]。为此,本研究以美丽鼠尾草和贵州鼠尾草为实验材料,研究了不同浓度的NaCl胁迫下,2种鼠尾草生长、叶绿素含量、保护酶活性和有机渗透调节物质含量的变化,比较了2种鼠尾草的耐NaCl胁迫能力及耐受性差异的生理机制,旨在为其引种、栽培等提供一定的理论依据。
1材料与方法
1.1实验材料
美丽鼠尾草和贵州鼠尾草分别引种自安徽省岳西县郊外和湖南张家界市,供试材料均为一年生的实生苗,栽植于北京师范大学珠海分校苗圃。
1.2实验设计
选择生长良好、长势基本一致的美丽鼠尾草和贵州鼠尾草各30株。种植于高度为20 cm、直径为18 cm的花盆中,每盆1株。培养基质厚度为17 cm,基质为土∶鸡粪按3∶1配比。待植株生长正常后,设5个NaCl胁迫处理浓度:200、300、400、500和600 mmol·L-1NaCl和1个对照(去离子水),处理和对照各5次重复,进行一次性NaCl胁迫处理。每盆植株浇灌相应浓度的NaCl处理液150 mL,对照浇灌等量的去离子水。处理后随机排列于室外进行培养。每天观察并拍照记录每盆植株的外部形态。当发现有植株濒临死亡时将所有植株收获,清洗植株并用吸水纸吸干残留水分。取第7片叶立即测定叶绿素含量,取2种鼠尾草植株的第4片叶密封于封口袋后立即保存于4 ℃冰箱用于测定渗透调节物质的含量,取第3片叶密封于封口袋后立即保存于-70 ℃冰箱中用于测定保护酶活性。
1.3项目测定
1.3.1植株受害等级评定观察植株的生长情况,划分为5个伤害等级:A-生长正常;B-部分叶片发黄;C-叶片发黄且叶缘卷曲;D-叶片局部变暗;E-叶片萎蔫,植株濒临死亡。
1.3.2叶绿素含量测定叶绿素含量测定采用比色法[18],用乙醇∶丙酮=3∶1混合液溶解叶绿素,分别在波长663和645 nm下测定吸光度,再计算单位鲜重中叶绿素的含量(mg·g-1)。
1.3.3酶活性测定SOD活性测定采用氮蓝四唑光还原法[18],在560 nm下以抑制NBT光化还原50%所需酶含量为1个酶活力单位(U)。CAT活力测定采用高锰酸钾滴定法[19],以每克鲜重样品3 min内分解过氧化氢的毫克数为酶活力单位。POD活性测定采用愈创木酚法[18],在470 nm波长下以每分钟OD增加0.01为1个活力单位。
1.3.4有机渗透调节物质含量的测定可溶性蛋白质含量的测定采用考马斯亮蓝G-250法[18],用结晶牛血清白蛋白制作标准曲线,在波长595 nm处测定吸光度,再计算单位鲜重中可溶性蛋白质含量(mg·g-1)。可溶性糖含量的测定采用蒽酮比色法[18],在625 nm波长下测定吸光度,再计算单位鲜重中可溶性糖含量(mg·g-1)。脯氨酸含量的测定采用酸性茚三酮比色法[18],以脯氨酸制作标准曲线,在波长520 nm处测定吸光度,再计算单位鲜重中脯氨酸含量(μg·g-1)。
1.4数据分析
利用SPSS 22.0统计软件进行单因素方差分析,并对平均值进行Duncan多重比较,显著性水平为0.05。绘图采用Microsoft Excel 2010。
2结果与分析
2.1NaCl胁迫对2种鼠尾草植株外部形态的影响
由表1可知,随着NaCl浓度的增加,植株表现出不同的盐害症状。当NaCl浓度为200 mmol·L-1时,美丽鼠尾草植株叶片开始发黄;随着NaCl浓度的增加,美丽鼠尾草大部分植株叶片发黄且叶缘卷曲;当NaCl浓度升高到600 mmol·L-1时,美丽鼠尾草受害更加严重,有植株叶片萎蔫甚至濒临死亡。而在300 mmol·L-1NaCl胁迫下,贵州鼠尾草仅有2盆植株叶片发黄,即使在NaCl浓度增加到600 mmol·L-1时,贵州鼠尾草也仅有3盆植株叶片发黄且叶缘卷曲,2盆植株部分叶片发黄,并无植株枝叶枯焦甚至濒临死亡。可见,贵州鼠尾草受盐害程度较美丽鼠尾草轻。
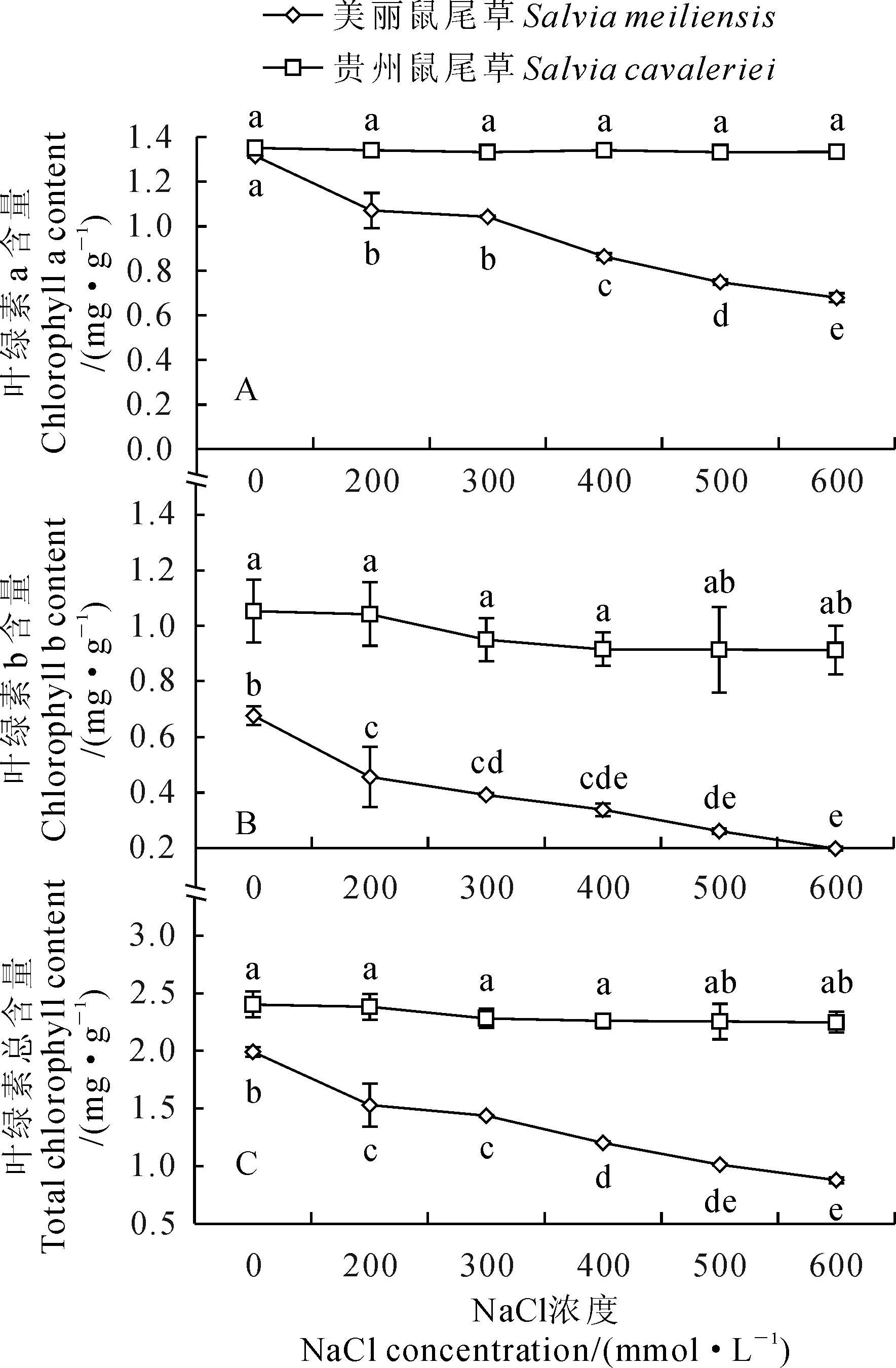
2.2NaCl胁迫对2种鼠尾草新叶叶绿素含量的影响
由图1可以看出,无NaCl胁迫时贵州鼠尾草叶片的叶绿素a、b和总含量均高于美丽鼠尾草。随NaCl浓度的增加,贵州鼠尾草叶片叶绿素a、b和总含量均无显著变化;而美丽鼠尾草叶片叶绿素a、b和总含量呈下降的趋势,三者含量在NaCl浓度为200 mmol·L-1时,较对照即已有显著降低。在所试NaCl浓度胁迫下,贵州鼠尾草叶片的叶绿素a、b和总含量均显著高于美丽鼠尾草。
2.3NaCl胁迫对2种鼠尾草保护酶活性的影响
由图2,A可知,贵州鼠尾草在不同浓度NaCl胁迫下SOD活性与对照相比无显著变化。美丽鼠尾草SOD活性随着NaCl浓度的增加呈先下降后上升再下降的趋势。在200 mmol·L-1NaCl浓度时,美丽鼠尾草SOD活性较对照降低但差异不显著,之后随NaCl浓度的增加SOD活性逐渐升高,在NaCl浓度为500 mmol·L-1时达到峰值,当NaCl浓度达到600 mmol·L-1时, SOD活性相对于500 mmol·L-1显著降低,但仍显著高于对照。由图2,B、C可知,随着NaCl浓度的增加,贵州鼠尾草的POD和CAT活性呈持续上升趋势,而美丽鼠尾草的POD和CAT活性呈先上升后下降的趋势,即贵州鼠尾草在NaCl浓度为600 mmol·L-1时POD和CAT活性仍在增加,而美丽鼠尾草POD和CAT活性却开始下降,其中CAT活性相对于500 mmol·L-1显著降低,但都显著高于对照。

表1 NaCl胁迫对2种鼠尾草的伤害等级
注:A.生长正常;B.部分叶片发黄;C.叶片发黄且叶缘卷曲;D.叶片局部变暗;E.叶片萎蔫,植株濒临死亡;Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ表示每个处理的5个重复。
Note:A.Normal growth;B.Partial leaves were yellow;C.The leaves were yellow and leaves margin were curly;D.Local leaves darkened;E.The leaves withered and the plants were on the verge of death;Ⅰ,Ⅱ,Ⅲ,Ⅳ and Ⅴ meant five replications of each treatment.
不同小写字母表示处理间差异显著(P<0.05);下同。
2.4NaCl胁迫对2种鼠尾草叶片有机渗透调节物质含量的影响
由图3可知,随着NaCl浓度的增加,美丽鼠尾草的脯氨酸、可溶性糖和可溶性蛋白3种渗透调节物质的含量均呈持续增加趋势,而贵州鼠尾草3种渗透调节物质的含量却呈先增加后减少的趋势。由图3,A可见,无NaCl胁迫时2种鼠尾草的脯氨酸含量基本相当,在NaCl浓度为200、300、500 mmol·L-1时,2种鼠尾草脯氨酸含量均无显著差异,仅在400和600 mmol·L-1时,美丽鼠尾草脯氨酸含量显著高于贵州鼠尾草;由图3,B可见,无NaCl胁迫时2种鼠尾草的可溶性糖含量基本相当,但在所试NaCl浓度胁迫下贵州鼠尾草可溶性糖含量均显著高于美丽鼠尾草;由图3,C可见,无NaCl胁迫时贵州鼠尾草可溶性蛋白质含量显著高于美丽鼠尾草,当NaCl浓度为200、300、400、500 mmol·L-1时,贵州鼠尾草可溶性蛋白质含量仍显著高于美丽鼠尾草,仅在NaCl浓度为600 mmol·L-1时,2种鼠尾草可溶性蛋白质含量基本相当,无显著差异。

图2 NaCl胁迫对美丽鼠尾草和贵州鼠尾草叶片抗氧化
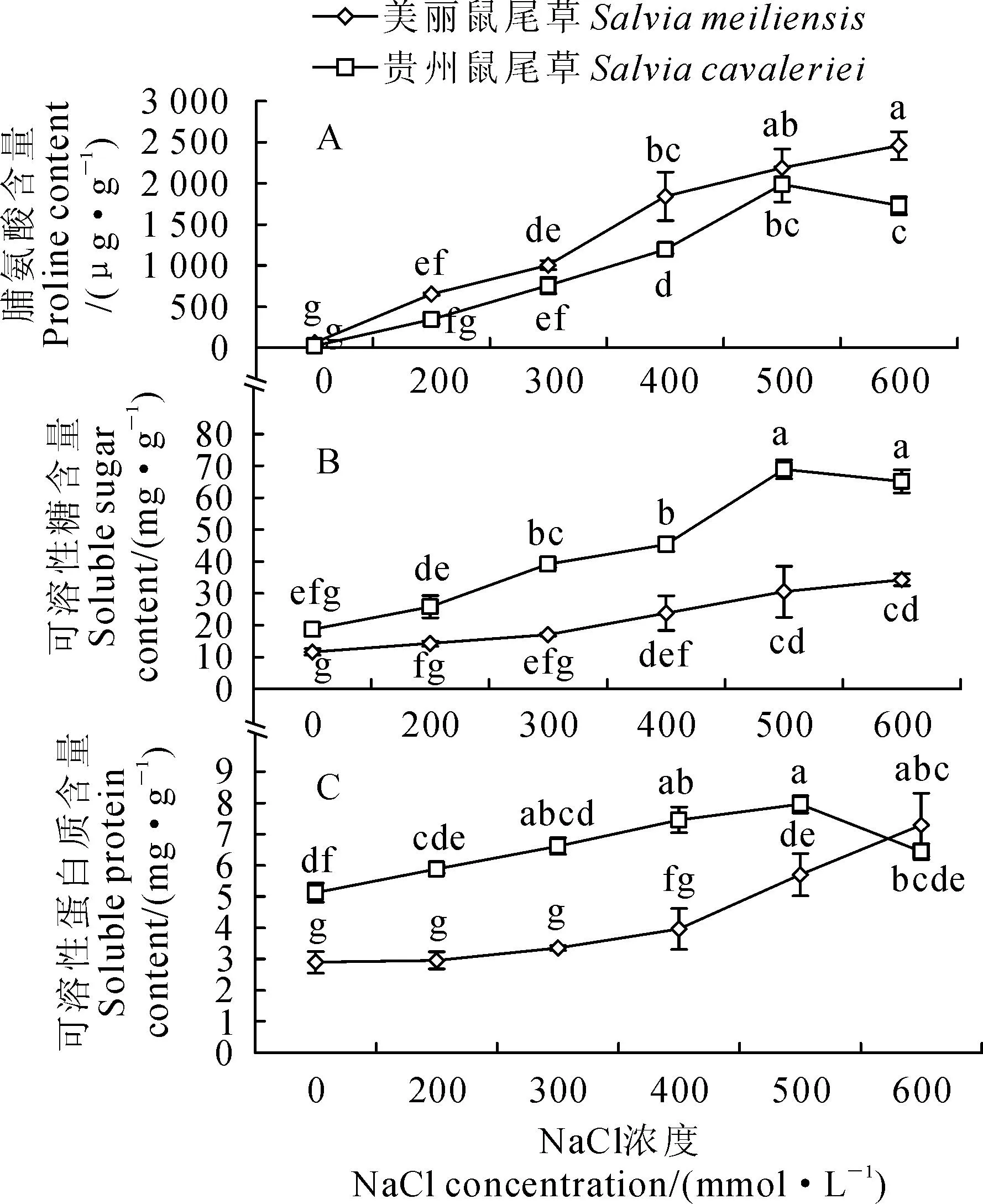
图3 NaCl胁迫对美丽鼠尾草和贵州鼠尾草叶片脯氨酸、
3讨论
叶绿素含量可以反映植物光合作用能力的强弱和植物光合产物含量的高低,从而间接反映植物的生长状况[20]。本实验以第7片新叶比较了贵州鼠尾草和美丽鼠尾草叶绿素的含量,对照中贵州鼠尾草的叶绿素a、b和总含量均高于美丽鼠尾草,说明贵州鼠尾草本身就具有比美丽鼠尾草较高的光合色素,且其叶绿素含量在各浓度的NaCl胁迫下均无显著变化,而美丽鼠尾草第7片新叶的叶绿素含量随NaCl浓度的增加逐渐降低,使其叶绿素含量在所试NaCl浓度胁迫下均显著低于贵州鼠尾草,表明贵州鼠尾草具有耐盐性,而美丽鼠尾草具有盐敏感性,这一结果与Meloni等[21]对不同棉花品种在盐胁迫下叶绿素含量的变化情况相似,即盐敏感型品种叶绿素含量降低,而耐盐型品种的叶绿素含量无显著变化。2种鼠尾草植株其它叶位的叶片,由于发生早,受盐害时间长,使得盐敏感性的美丽鼠尾草叶片黄化、变暗等程度明显高于耐盐性的贵州鼠尾草。

渗透调节机制是植物在长期进化过程中演化出的适应不利环境的策略和重要的生理基础[26-27]。在盐胁迫下,脯氨酸、可溶性糖和可溶性蛋白质等有机渗透调节物质的积累可以帮助植物细胞维持正常膨压,便于植物在盐渍环境中吸收水分,保证细胞正常生理代谢[28-29]。 本实验中,NaCl浓度从200 mmol·L-1增加到600 mmol·L-1,美丽鼠尾草和贵州鼠尾草的脯氨酸、可溶性糖和可溶性蛋白质的含量均高于各自的对照,说明2种鼠尾草对NaCl胁迫均产生适应性反应,通过提高这3种渗透调节物质的含量来降低水势,增强对土壤水分的吸收能力,一定程度上缓解盐害。这与张超强等[30]对酸浆(Physalisalkekengi)试管苗的研究结果相似。然而,在所试NaCl浓度胁迫下,贵州鼠尾草可溶性糖含量均显著高于美丽鼠尾草,NaCl浓度为200、300、400、500 mmol·L-1时,贵州鼠尾草可溶性蛋白质含量也显著高于美丽鼠尾草,使得贵州鼠尾草在NaCl胁迫下具有比美丽鼠尾草更强的渗透调节能力,这可能是其耐盐性强于美丽鼠尾草的重要原因。
综上所述,贵州鼠尾草的保护酶比美丽鼠尾草对NaCl胁迫的适应性阈值高,是贵州鼠尾草受害较美丽鼠尾草轻的原因之一。其次,在NaCl胁迫下贵州鼠尾草可溶性糖和可溶性蛋白2种渗透调节物质的含量均显著高于美丽鼠尾草,可能是贵州鼠尾草受害较美丽鼠尾草低的原因之二。2种鼠尾草对NaCl胁迫的耐受性差异与其各自的生物学特性及其原生境息息相关。本研究供试的美丽鼠尾草生长于林缘路边水分条件较好的生境,其根系为直根系,主根较粗壮,喜土层深厚且排水良好的沙壤土;而贵州鼠尾草为须根系,植株矮小,生境为溪边裸露的岩壁或土层较浅的石缝中。在长期的进化适应过程中,2种鼠尾草形成了各自迥异的生理生态特性和抗逆机制,从本研究中所表现出的对NaCl胁迫耐受性的差异得到了证实。因此,在对贵州鼠尾草和美丽鼠尾草进行引种、栽培时,应充分考虑2种鼠尾草对逆境的适应能力,才能做到因地制宜从而使经济效益最大化。
本研究仅从生理适应的角度初步阐明了2种鼠尾草对NaCl胁迫的耐受差异,显然还不能揭示2种鼠尾草对NaCl胁迫耐受性差异的深层次机制,因此,相关抗逆性基因的表达、信号通路以及相关渗透物质生物化学代谢的调节过程将是今后研究的重点。
参考文献:
[1]俞仁培.对盐渍土资源开发利用的思考[J].土壤通报,2001,32(1):138-140.
YU R P.Ideas about exploitation and utilization of saline soil resource[J].ChineseJournalofSoilScience,2001,32(1):138-140.
[2]高永生,王锁民,张承烈,等.植物盐适应性调节机制的研究进展[J].草业学报,2003,12(2):1-6.
GAO Y S,WANG S M,ZHANG C L,etal.Plant adaptive and regulation mechanism under salt stress[J].ActaPrataculturaeSinica,2003,12(2):1-6.
[3]杨劲松.中国盐渍土研究的发展历程与展望[J].土壤学报,2008,45(5):837-845.
YANG J S.Development and prospect of the research on salt-affected soils in China[J].ActaPedologicaSinica,2008,45(5):837-845.
[4]王艳青,陈雪梅,李悦,等.植物抗逆中的渗透调节物质及其转基因工程进展[J].北京林业大学学报,2001,23(4):66-70.
WANG Y Q,CHEN X M,LI Y,etal.The osmotic solute in plant resistance to adervse conditions and progress in relative genic engineering[J].JournalofBeijingForestryUnivesity,2001,23(4):66-70.
[5]中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1977:66.
[6]安徽植物志协作组.安徽植物志[M].合肥:安徽科学技术出版社,1991:272.
[7]赵宝林,刘学医,刘宏,等.安徽药用丹参资源调查研究[J].现代中药研究与实践,2010,24(5):1-32.
ZHAO B L,LIU X Y,LIU H,etal.Investigation and research of medicinalSalviamiltiorrhizasresource in Anhui Province[J].MedicinalResearchandPracticeofChineseMedicines,2010,24(5):1-32.
[8]国家药典委员会.中国药典[S].北京:化学工业出版社,2005:52.
[9]景鹏飞.丹参组药用植物资源调查和多样性评价研究[D].西安:陕西师范大学,2013.
[10]李旻辉.中国鼠尾草属药用植物亲缘学研究[D].北京:中国协和医科大学,2008.
[11]王和英,胡德禹,薛伟,等.贵州鼠尾草化学成分的研究[J].天然产物研究与开发,2011,23(1):63-65.
WANG H Y,HU D Y,XUE W,etal.Chemical constituents in herbs ofSalviacavalerieiLévl.[J].NaturalProduceResearchandDevelopment,2011,23(1):63-65.
[12]LIN L Z,WANG X M,HUANG X L,etal.Sapriolactone,a cytotoxic norditerpene fromSalviaprionitis[J].Phytochemistry,1989,28(12):3 542-3 543.
[13]乐秀芳,韩家娴,沈祖铭,等.红根草邻醌的体内外抗癌作用研究[J].肿瘤,1992,12(2):49-50.
LE X F,HAN J X,SHEN Z M,etal.Anticancer activity of saproorthoquinone[J].Tumor,1992,12(2):49-50.
[14]HOQUE M A,OKUMA E,BANU M N A,etal.Exogenous proline mitigates the detrimental effects of salt stress more than exogenous betaine by increasing antioxidant enzyme activities[J].JournalofPlantPhysiology,2007,164(5):553-561.
[15]Qiyan Jiang,Zheng Hu,Hui Zhang,etal.Overexpression of GmDREB1 improves salt tolerance in transgenic wheat and leaf protein response to high salinity[J].TheCropJournal,2014,2(3):120-131.
[16]刘爱荣,赵可夫.盐胁迫下盐芥渗透调节物质的积累及其渗透调节作用[J].植物生理与分子生物学学报,2005,31(4):389-395.
LIU A R,ZHAO K F.Osmotica accumulation and its role in osmotic adjustment inThellungiellahalophilaunder salt stress[J].JournalofPlantPhysiologyandMolecularBiology,2005,31(4):389-395.
[17]PATAKAS A,NIKOLAOU N,ZIOZIOU E,etal.The role of organic solute and ion accumulation in osmotic adjustment in drought stressed grapevines[J].PlantScience,2002,163(2):361-367.
[18]陈建勋.植物生理学实验指导[M].广州:华南理工大学出版社,2006:24-26,55-56,66-70,81-84.
[19]李合生,孙群,赵世杰,等.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:165-167,258-260.
[20]王金龙,赵念席,徐华,等.不同地理种群大针茅生理生化特性的研究[J].草业学报,2011,20(5):42-48.
WANG J L,ZHAO N X,XU H,etal.A study on the physiological and biochemical characters of differentStipagrandisgeographical populations[J].ActaPrataculturaeSinica,2011,20(5):42-48.
[21]MELONI D A,OLIVA M A,MARTINEZ C A,etal. Photosynthesis and activity of superoxide dismutase,peroxidase and glutathione reductase in cotton under salt stress[J].EnvironmentalandExperimentalBotany,2003,49(1):69-76.
[22]WANG J,LI D Q.The accumulation of plant osmoticum and active oxygen metablism under stress[J].ChineseBulletinofBotany,2001,18(4):459-465.
[23]陈娟,谷巍,段金廒,等.穿心莲生理特性及盐胁迫对其影响研究[J].中药材,2014,37(8):1 320-1 325.
CHEN J,GU W,DUAN J A,etal.Study on physiological characteristics and effects of salt stress inAndrographispaniculata[J].JournalofChineseMedicinalMaterials,2014,37(8):1 320-1 325.
[24]赵可夫,邹琦,李德全,等.盐分和水分胁迫对盐生和非盐生植物细胞膜脂过氧化作用的效应[J].植物学报,1993,35(7):519-523.
ZHAO K F,ZOU Q,LI D Q,etal.The effect of salt and water stress on membrane lipid peroxidation in leaf cells of halophyte and non-halophyte[J].ActaBotanicSinica,1993,35(7):519-523.
[25]PARIDA A K,DAS A B.Salt tolerance and salinity effects on plants:a review[J].EcotoxicologyandEnvironmentalSafety,2005,60(3):324 -349.
[26]QUARRIE S A,STOJANOVIC J,PEKIC S.Improving drought resistance in small-grained cereals:a case study,progress and prospects[J].PlantGrowthRegulation,1999,29(1-2):1-21.
[27]COSTA F M G,PHAM Thi A T,PIMENTEL C,etal.Differences in growth and water relations amongPhaseolusvulgariscultivars in response to induced drought stress[J].EnvironmentalandExperimentalBotany,2000,43(3):227-237.
[28]武香,倪建伟,张华新,等.盐胁迫下不同盐生植物渗透调节的生理响应[J].东北林业大学学报,2012,40(8):29-33.
WU X,NI J W,ZHANG H X,etal.Effects of salt stress on osmotic adjustment substances in different halophytes[J].JournalofNortheastForestryUniversity,2012,40(8):29-33.
[29]裘丽珍,黄有军,黄坚钦,等.不同耐盐性植物在盐胁迫下的生长与生理特性比较研究[J].浙江大学学报(农业与生命科学版),2006,32(4):420-427.
QIU L Z,HUANG Y J,HUANG J Q,etal.Comparative study on vegetal and physiological characteristics of different salt-tolerant plants under salt stress[J].JournalofZhejiangUniversity(Agriculture and Life Science),2006,32(4):420-427.
[30]张超强,王兴龙,杨致远,等.NaCl胁迫对酸浆试管苗生理特性的影响[J].中药材,2014,37(10):1 725-1 729.
ZHANG C Q,WANG X L,YANG Z Y,etal.Effect of NaCl stress on physiological characteristics ofPhysalisalkekengivar.franchetiitest-tube plantlet[J].JournalofChineseMedicinalMaterials,2014,37(10):1 725-1 729.
(编辑:潘新社)
Comparation of Tolerance under NaCl Stress between TwoSalviaPlants and Study of the Physiological Mechanisms
XU Hua1,LIANG Chunhong1,ZHAO Meitang1,WEI Yukun2*,HUANG Yanbo2
(1 College of Engineering Technology,Beijing Normal University Zhuhai Compus,Zhuhai,Guangdong 519087,China;2 Shanghai Chenshan Botanical Gardern,Shanghai Chenshan Plant Science Research Center,Chinese Academy of Sciences,Shanghai 201602,China)
Abstract:In order to ensure different tolerances of two Salvia plants to NaCl stress and its physiological mechanism,we used S.meiliensis and S.cavaleriei with high medicinal and ornamental value as experimental materials to analyze the variation of their growth,chlorophyll content,protective enzymes activities and their content of organic osmotic regulation substances under 0,200,300,400,500 and 600 mmol·L(-1 )NaCl.The results showed that:1)under each concentration of NaCl stress,S.meiliensis suffered from the salt damage more severely than S.cavaleriei.As NaCl concentrations increased.2)The chlorophyll content in the leaves of S.cavaleriei had no significant change,but those in the leaves of S.meiliensis decreased gradually.3) When NaCl concentration increased from 0 to 500 mmol·L(-1),S.cavaleriei and S.meiliensis plants responded adaptively to NaCl stress.The POD and CAT activities,as well as soluble sugar,soluble protein and proline contents gradually inceased in the leaves of S.cavaleriei and S.meiliensis.The SOD activity also increased in the leaves of S.meiliensis.4) When NaCl concentration reached to 600 mmol·L(-1),the soluble sugar,soluble protein and proline contents in the leaves of S.meiliensis continued to increase,while the SOD,POD and CAT activities started to drop,but were still significantly higher than those of the control.In contrast,at the same NaCl concentration,the POD and CAT activities in the leaves of S.cavaleriei continued to increase.The soluble sugar,soluble protein and proline contents started to drop,but were still significantly higher than those of the control.S.cavaleriei possesses higher contents of osmotic regulation substances under NaCl stress and can maintain higher protective enzymes activities with increase of NaCl concentration,it has stronger tolerance to NaCl than S.meiliensis.
Key words:NaCl stress;Salvia cavaleriei;Salvia meiliensis;osmotic regulation substance;protective enzyme
中图分类号:Q945.79
文献标志码:A
作者简介:许华(1981-),硕士,副教授,主要从事植物生理生态学研究。E-mail:xuhua04@mail.nankai.edu.cn*通信作者:魏宇昆,博士,高级工程师,主要从事植物生态学与保育生物学研究。E-mail:ykwei@sibs.ac.cn
基金项目:中国科学院战略生物资源科技支撑体系运行专项(CZBZX-1);广东省高等学校教育教学改革项目(GDGJ20142513);广东省大学生创新训练项目(201413177034)
收稿日期:2015-11-30;修改稿收到日期:2016-03-08
文章编号:1000-4025(2016)03-0558-07
doi:10.7606/j.issn.1000-4025.2016.03.0558
