辽东栎林内不同小生境下幼树植冠构型分析
2016-05-06张文辉于世川邢忠利黄财智
李 罡,张文辉,于世川,邢忠利,黄财智
(西北农林科技大学 西部环境与生态教育部重点实验室,陕西杨陵 712100)
辽东栎林内不同小生境下幼树植冠构型分析
李罡,张文辉*,于世川,邢忠利,黄财智
(西北农林科技大学 西部环境与生态教育部重点实验室,陕西杨陵 712100)
摘要:以黄土高原黄龙山林区辽东栎林内3个小生境(林下、林隙、林缘)下辽东栎天然更新幼树为研究对象,采用典型抽样法对辽东栎幼树侧枝、叶片和树冠的空间分布状况以及生物量分配状况进行调查分析,探讨微生境与幼树植冠构型特征的关系,明确辽东栎幼树对不同小生境的适应策略,为栎林经营和林分结构优化提供理论依据。结果显示:(1)3种生境下辽东栎幼树构型发生了可塑性变化,林下幼树树冠层次比较单一,林隙与林缘的幼树树冠层次更加丰富。(2)由林下至林缘,幼树的树高、枝下高呈逐渐减小的趋势,而地径变化趋势与之相反;幼树的冠幅、树冠面积、树冠率呈先增加后减小的趋势,并且林下与林隙、林缘的差异显著;幼树的总体分枝率、逐步分枝率、枝径比呈先增加后减小的趋势。(3)3种生境下,幼树的一级枝的枝长、直径与倾角随着树高的增加而呈减小的趋势,但3种生境的差异不显著;林下一级枝主要分布在冠层中上部,而林隙与林缘一级枝主要分布在冠层中下、中上部。(4)由林下至林缘幼树叶长、叶宽、单叶面积和比叶面积逐渐降低,而单株叶数、叶总面积、叶面积指数呈先增大后减小趋势;与其他2种生境相比林下叶片分布趋于冠层上部。(5)幼树地上部分生物量中林下主干生物量占83%,枝和叶生物量只占17%;而林隙与林缘虽然各部位生物量有所差异但比例基本一致,其中主干占66%左右,枝和叶生物量占34%左右。研究表明,林隙生境下幼树的构型优于林缘和林下生境,在今后栎林的经营中,可以通过适当间伐来增加林隙数量,为森林更新和结构的优化提供有利条件。
关键词:辽东栎;幼树;植冠构型;生物量分配
自然界中,木本植物是由不同的构件组成的,可以认为植物是一个构件的集合体[1-2]。构型是植物内在遗传信息在一定时间表现的外部形态特征,因此,构型分析的实质是分析植物体不同构件在空间的排列方式[3-4]。植物的个体构型是由基因型决定的,但在一定程度上又受到环境条件的影响,所以可以认为植物的构型是植物基因型和环境之间相互作用的产物[5]。植物根系是固着在土壤里的,在木本植物的生长发育过程中,当外部环境和生态因子变化时,它们会通过形态可塑性来适应环境,从而形成不同的形态和构型,提高其生存适合度和竞争能力[6-7]。植物体构型不仅决定了光捕获效率以及未来的生长方向,也影响植株的生产效率[5]。幼树是即将进入林冠层的阶段,这一时期的构型变化将影响植株未来的生长方向。因此,研究生境对幼树构型差异的影响对森林结构优化和经营措施的制定具有重要的参考意义。
辽东栎(Quercusliaotungensis)是壳斗科(Fagaceae)栎属落叶乔木,主要分布于辽东半岛北部的丘陵山区山西恒山以南的山地、河北北部的土石山地、甘肃和陕西的黄土高原,南达秦岭中段北坡及其西段等地[8],辽东栎林是黄土高原一种重要的森林类型,被认为是该区域较为稳定的顶级森林群落[9]。黄土高原黄龙山林区,辽东栎林为主要地带性成林树种之一,对当地的水土保持与生态区域安全发挥着重要作用。长年的封山育林虽然对林地生态功能的恢复起了一定的作用,但是由于森林缺乏必要的抚育措施,林木干形弯曲品质下降,多年生幼苗较少使得更新潜力降低。
国外没有辽东栎,但是对其它树种的构型研究较早。主要集中在研究树木不同生境下的分枝率以及几何构型等方面[10-12]。Borchert和Slade的研究表明,Populusdeltoides分枝比受树冠大小的影响显著,树冠越大分枝比越高[13]。Steingraeber和Kascht等对不同生境下的糖槭树幼苗构型研究表明,林下生长的幼苗的叶子仅分布于几个平面层,而旷地生长的幼苗冠层更全,分枝更丰富[14]。国内对辽东栎构型的研究主要在20世纪90年代,孙书存等对不同年龄阶段辽东栎研究发现构型转变主要表现在分枝格局上,除枝、叶方位角和叶倾角外,分枝率、枝长和枝倾角都发生过显著改变[2,15]。胡晓静、余碧云等对栓皮栎构型研究发现,不同生境与径级下栓皮栎构型差异性显著[16-17]。近年来关于辽东栎的研究大多集中在种群生态、生殖生态等方面[18-20],对辽东栎构型方面研究较少。本研究以不同生境下(林下、林隙、林缘)的辽东栎天然更新幼树为研究对象,通过统计分析,阐述辽东栎幼树对不同生境的适应策略,进一步探讨生境与构型的关系,为黄龙山林区的栎林经营和林分结构优化提供依据。
1研究地区与研究方法
1.1研究区域概况
研究样地设在分布较集中的黄土高原黄龙山林区,地理位置为109°38′49″~110°12′47″E,35°28′46″~36°02′01″N,海拔 962.6~1 783.5 m,属暖温带半湿润与半干旱气候的过渡地带,年平均降水量606.2 mm,年平均气温8.6 ℃。最高气温36.7 ℃,最低气温-22.5 ℃,≥10 ℃的有效积温2 953.7 ℃。本区土壤类型主要为灰褐土。地带性植被是暖温带落叶阔叶林,辽东栎(Q.liaotungensis)、油松(Pinustabulaefomis)、白桦(Betulaplatyphylla)是当地现存最为典型的主要树种。林下灌木层主要有黄刺玫(Rosaxanthina)、卫矛(Euonymusalatus)、陕西荚蒾(Viburnumschensianum)、西北栒子(Cotoneasterzabelii)等,草本层主要有苔草(Carextristachya)、山萝花(Melampyrumroseum)、披碱草(Elymusdahuricus)等。辽东栎林是当地分布面积最大发育最好的天然林之一。
本实验以阳坡辽东栎幼树为研究对象,研究区辽东栎林没有进行过较大规模的采伐或破坏活动,林相发育较好。幼树生长环境为林下、林隙和林缘3个生境,分别在这3个生境设置样地(各生境林分与立地特征如表1所示)。林下样地设在林相整齐林木分布均匀地段(样地中心距林缘>40 m),每个样地大小30 m×30 m共3个;在林下寻找3个大小100~150 m2的林隙做为林隙样地;林缘样地设在没有人为干扰的林分边缘,设置3个大小10 m×50 m的样地。
1.2研究方法
1.2.1基本数据调查调查的辽东栎为胸径4 cm左右的天然更新幼树,采用典型抽样法在每个生境(林下、林隙、林缘)样地中选择长势良好的辽东栎幼树各12棵。选定后,用卷尺测定幼树的地径、冠幅、树高、枝下高,并计算树冠率(冠幅与树高比值);用游标卡尺测定各级枝直径,并统计枝上的叶数,测量叶长(去除叶柄的长度)和最宽处宽度。为了研究不同冠层中的侧枝和叶片分布格局,把每株幼树按树高分为4个层次:0~0.25、0.25~0.5、0.5~0.75、0.75~1.0,以每个 1 级侧枝在主干上的位置为准,按侧枝所处的高度确定所属的冠层。
1.2.2分枝格局与叶片特征调查在冠层内对一级活枝由下到上进行编号,用卷尺测定一级枝长度、枝间距、到顶距离,枝间距为一级枝在主干上的垂直距离;枝径比为二级侧枝与一级侧枝的直径比;用圆规结合半圆仪测定一级枝倾角。测定1~3级枝数,枝序的确定采用由内及外的方法,即着生在主干上的为1级枝,着生在1级枝上的为2级枝,着生在2级枝上的为3级枝,并计算树木的总体分枝率和逐步分枝率。总体分枝率Rb和逐步分枝率Ri+1:i的计算方法[21]如下:
Rb=(Nt-Ns)/(Nt-Nl)
式中:Nt为枝条总数;Ns为最高枝条数;Nl为第一枝条数。
Ri+1:i=Ni+1/Ni
式中:Ni和Ni+1分别为第i级和第i+1级的枝条总数。
叶片使用Epson Perfection 4870进行扫描,使用WinFOLIA 2004a软件分析叶面积(LA),计算相对高度上的叶面积指数(LAI)、叶重比(LMR)、比叶面积(SLA)和叶面积比(LAR) ,公式如下:
LAI=LA×N/(π×r2)
LMR=ML/M
SLA=LA/Ms
LAR=LA×N/M
式中:LA为平均单叶面积;N为相应高度上的叶片数;r为树冠冠幅的半径;ML为相对树高上的总叶片质量;M为总地上生物量;Ms为单叶质量。
1.2.3生物量分配调查将一级枝从树干上分离,摘掉叶片并统计叶片数,用电子秤称一级枝枝鲜重、叶鲜重以及树干鲜重,并取样带回实验室进行烘干(85 ℃)处理,测定枝、干、叶干重,其中叶取样随机抽取200片叶子,枝、干取样分上、中、下三段截取。
1.3数据处理
采用Excel 2013和SPSS 18.0 软件对数据进行统计分析,采用单因素方差分析(one-way ANOVA)和LSD法进行方差分析和多重比较(α=0.05)。使用Origin 9软件作图。图表中数据为平均值±标准差。

表1 不同小生境下辽东栎林分与立地特征
注:Ⅰ.林下; Ⅱ.林隙; Ⅲ.林缘;下同。
Note:Ⅰ.Under story ;Ⅱ.Forest gap ;Ⅲ.Forest edge;The same as below.
2结果与分析
2.1不同小生境下辽东栎幼树的总体构型分析
林下到林缘由于小生境的变化,导致植物以不同的构型转变来适应环境异质性,并产生相应的适应策略,以保证植株在资源获取和空间竞争中取得优势。从林下至林缘,最主要的变化就是光照环境的变化,光照环境是影响构型的重要因素。由表2可以看出,不同生境对辽东栎幼树的地径、胸径与树高影响不显著;由林下至林缘,幼树的地径呈增大趋势而树高变化趋势与之相反,胸径林隙最大。不同小生境下,辽东栎幼树的枝下高、冠长、树冠面积、树冠率差异性显著;林下至林缘,枝下高逐渐减小并且林下与林隙、林缘差异性显著;而冠长、树冠率、树冠面积呈先增大后减小趋势,林下林下与林隙、林缘差异性显著。
说明在荫蔽条件下,辽东栎幼树尽可能减少侧枝生物量投入而加大对高生长的投入,使幼树尽可能快地达到主林层,增加存活几率;而光照充足的林隙和林缘生境,辽东栎冠层的层次感更加丰富。3种小生境下,辽东栎幼树总体构型表现为林隙最佳。
2.2不同小生境下辽东栎幼树的枝系构型分析
由表3可以看出,不同小生境下辽东栎幼树的总体分枝率、逐步分枝率、枝间距和枝径比差异性均达到显著水平,说明在不同小生境下,辽东栎幼树采取不同的分枝格局来适应环境异质性变化。由林下至林缘幼树的总体分支率、逐步分枝率和枝径比均呈现先增大后减小的趋势,并且林隙与林下生境差异性显著,说明林隙下辽东栎幼树分枝更多,生命力更加旺盛。而枝间距由林下至林缘呈现逐渐增大的趋势,林下与林缘、林隙差异性显著。
辽东栎幼树侧枝的垂直分布情况,由图1可以看出,在0~0.25这一相对高度处只有枝下高相对较低的林缘有一级枝分布。3种小生境下,一级枝的长度、直径、倾角均随着树高的增加而逐渐减小;一级枝数林下与林缘呈逐渐增加趋势,林隙呈先增加后减小趋势。同一冠层下由林下至林缘,一级枝长呈先增加后减小的趋势,并且顶层枝条(>0.75)林隙与林下、林缘差异性显著,其余冠层差异不明显;一级枝直径除顶层林隙最大外,其余冠层均林缘最大;一级枝倾角除去底层树冠(<0.25)外林下倾角最大,并且在顶层与林隙、林缘差异显著;除去林缘在树冠底层有一级枝分布外,其余2个生境均没有一级枝分布,在树冠中下层(0.25~0.5)和中上层(0.5~0.75)林隙一级枝密度最大,而在顶层林下一级枝密度居3种生境之首。
2.3不同小生境下辽东栎幼树的叶片构型分析
叶片形态是指示植物适应特定环境的重要指标,植物叶片形态对时空环境变化具有极强的敏感性和可塑性,能够通过叶片形态的调整调节自身的生存适应能力[22]。本研究中(表4)可以看出,由林下至林缘这一生境的变化,引起了叶片形态的适应性变化,其中叶长、叶宽、单叶面积、比叶面积呈现逐渐减小的趋势,林下与林缘差异性显著,而叶片长宽比基本保持一致,差异性并不显著;总叶面积和单株叶数林隙达到最大,林缘次之,林下最小,其中林隙与林下差异性显著。

表2 不同小生境下辽东栎幼树总体构型
注:同列不同小写字母表示处理间差异显著(P<0.05);下同
Note:Different normal letters in the same column mean significant difference at 0.05 level among treatments;The same as below.

表3 不同小生境下辽东栎幼树分枝格局
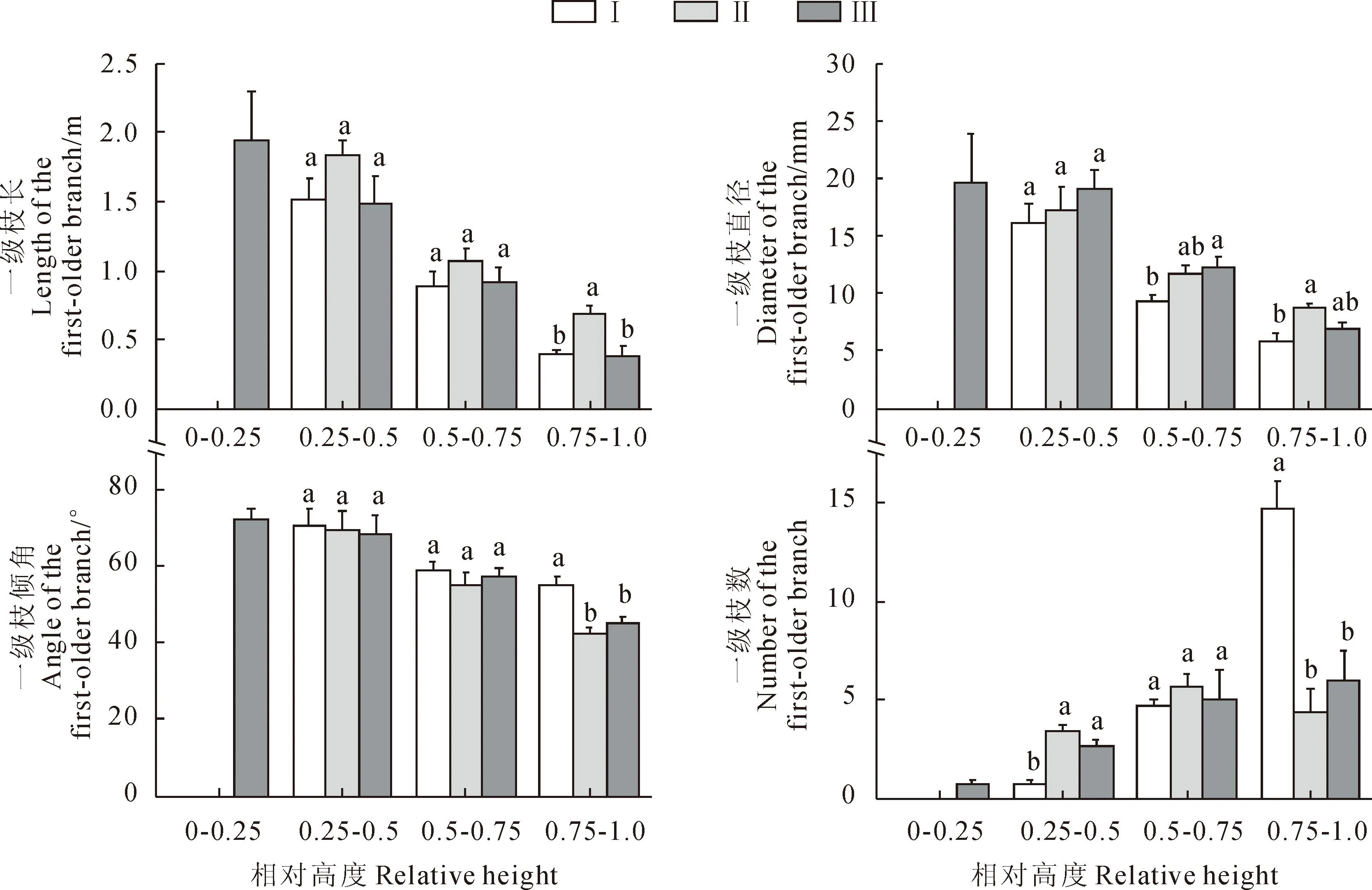
不同小写字母表示同一高度处差异显著;下同。
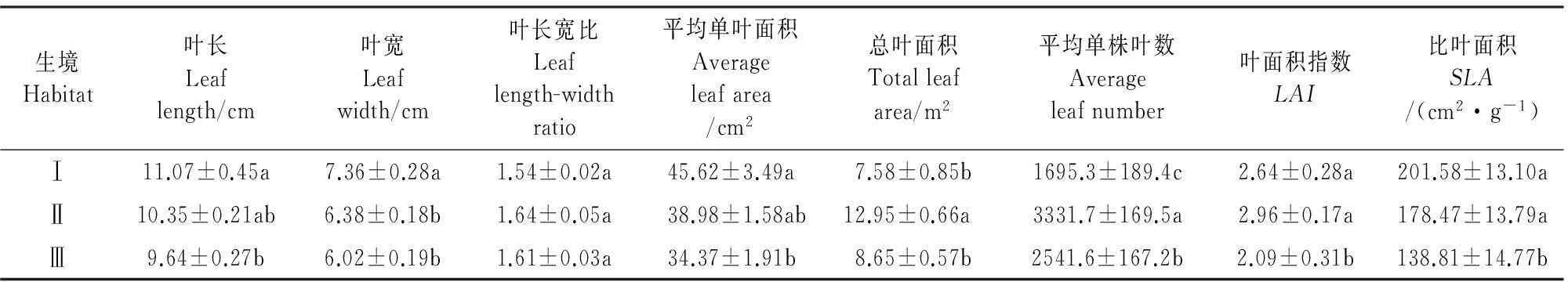
生境Habitat叶长Leaflength/cm叶宽Leafwidth/cm叶长宽比Leaflength-widthratio平均单叶面积Averageleafarea/cm2总叶面积Totalleafarea/m2平均单株叶数Averageleafnumber叶面积指数LAI比叶面积SLA/(cm2·g-1)Ⅰ11.07±0.45a7.36±0.28a1.54±0.02a45.62±3.49a7.58±0.85b1695.3±189.4c2.64±0.28a201.58±13.10aⅡ10.35±0.21ab6.38±0.18b1.64±0.05a38.98±1.58ab12.95±0.66a3331.7±169.5a2.96±0.17a178.47±13.79aⅢ9.64±0.27b6.02±0.19b1.61±0.03a34.37±1.91b8.65±0.57b2541.6±167.2b2.09±0.31b138.81±14.77b
叶片的垂直分布特征由图2可以看出,林隙与林缘生境由底层到顶层叶片随着树高的增加呈现增加后减小的趋势,而林下呈逐渐增大的趋势。除去底层只有林缘由叶片分布外,中下层与中上层林隙叶片分布最多,并与林下、林缘差异性显著,而顶层叶片分布林下最多与林隙、林缘差异性显著。其中叶面积指数的垂直变化基本与叶片数目的趋势相同,不同的是林下的叶面积指数显著增大。叶面积比在中下层林缘最大,林隙次之,2种生境与林下差异显著;中上层林隙最大,与林下林缘差异性显著;顶层林下最大并与其余2个生境差异显著。比叶重在中下层林缘最大并与林隙、林缘差异显著;在中上层林隙与林缘差异不大,与林下差异显著;在顶层林下表现最好,与其余2个小生境差异显著。
2.4不同小生境下辽东栎幼树的生物量分配分析
由图3 可以看出,主干生物量占总体生物量的比重较大,其中林下最大达到83%,林隙与林缘比重大致相同,为66%左右,而枝与叶生物量所占比重则是林隙与林缘较多,分别达到24%与9%,林下只为10%和6%。单就生物量而言,主干生物量林隙最大,林下次之,林缘最小;枝与叶的生物量均是林隙最大,林缘次之,林下最小,并且3种小生境下差异显著。
3讨论
3.1不同小生境辽东栎幼树总体构型
植物是固着在土壤里生长的,但是由于植物生存环境的异质性等原因,遗传上完全相同的植物其个体生活方式可能完全不同,其采取的生存策略也不尽相同[6]。构型是植物本身与其生长环境相互作用的结果, 一般认为在特定生境中,植物体可以通过性状的可塑性表达,形成与环境相适应的有利特征[2]。因而,对不同生境中的植物构型进行比较一定程度上可以反映植物体的不同适应对策。有学者认为,体积小、着生大量高重叠率短枝叶片的紧密型树冠通常被认为是树木对强光环境的一种适应行为[23];在没有胁迫和干扰生境中的植物具有较大的根冠比以及较大的根和叶面积[24],这与本研究的结果相同。本研究中,由林下至林缘光照逐渐加强,处于林下的辽东栎幼树由于受大树植冠的荫蔽、 压迫以及接受的光照有限,以牺牲分枝率为代价,投资高生长,从而摆脱胁迫环境,迅速达到林冠上层,因此树高、枝下高较高但冠幅、分枝率、树冠率较低;处于林隙的辽东栎幼树光照、水热条件适宜,因此幼树生长旺盛分枝率高、冠幅较大;而处在林缘的幼树树高、枝下高较低、叶面积较小,但树冠层次丰富叶片排列紧密,这是幼树对较强光照的一种适应性。

图2 不同小生境下辽东栎幼树叶片垂直分布特征

图3 不同小生境下辽东栎幼树生物量分配
3.2不同小生境下叶片形态
叶片形态是指示植物适应特定环境的重要指标[22],叶片形态不仅受基因控制,同时对不同尺度环境变化都具有极强的敏感性[25-26]。有研究认为,生长在干旱生境条件下的植物,往往通过减少单叶叶面积、降低叶面积指数来减少水分的丧失[27],这与本研究结论一致。叶面积指数在一定程度上反映了叶片截获光的能力,其值越大,光利用效率越高[28]。在本研究中,处于林隙的辽东栎幼树总叶数最多,叶面积指数最大,光能利用率较高,这也说明林隙生境更加适合辽东栎幼树生存。而处在林缘的幼树虽然平均单叶面积与叶面积指数较小,但是叶片厚度比其余2个生境要大(比叶面积较小),从而增加了光合效率,这也是植物对高光强与干旱生境的一种适应性。
3.3枝系与叶片垂直分布特征与生物量分配
一般情况下,林分中的光照是随着高度的增加而逐渐增强的,幼树枝系与叶片在垂直方向上的分布与排列与光能的获取、营养获取、空间占用密切相关[29]。通常情况下,树冠上部枝长较下部小,中部枝系和每枝叶数比上部、下部枝大[30],树木总叶片数随树高的增高而增大,叶面积指数也随树干的增长而增加[25]。叶重比、 叶面积比与植物光合能力密切相关[31]。在本研究中,处于林下的幼树枝系与叶片分布主要分布在树冠中上部,叶重比、叶面积比、叶面积指数的较大值也出现在树冠中上部,这就使得幼树在较荫蔽的环境下尽可能多的获取林下散射光,提高光合效率达到正常生长的目地,这也与前人研究结论一致[17]。处于林隙的幼树较大枝条出现在树冠中下、中上部,叶片数也是这2个层次较多分枝率高,因此处于林隙的幼树冠幅较大,并且叶重比、叶面积比、叶面积指数的最大值也分布在这2个层次,幼树光合能力较强,生长旺盛。而处于林缘的幼树的枝系和叶片的分布趋势与林隙的大致相同,但是各种指标较林隙差,但林缘的幼树树冠层次更加丰富,4个层次均有枝系分布,这也是幼树对强光干旱生境的适应性。生物量分配方面,处于林下的幼树,为了摆脱荫蔽条件以牺牲侧枝为代价投资高生长,因此生物量主要集中在主干上;而处于林隙与林缘的幼树,林隙生物量较林缘多,但分布比例相差不多。由此可见,在不同的小生境下,辽东栎幼树以不同的构型来达到资源与空间利用的最大化,以达到正常生长的目的。
因此,在辽东栎天然林抚育过程中,需确定适宜的间伐强度增加林隙数量来促进林下幼苗、幼树更新以及优良干才的形成,在保证林分防护功能的同时,增加林分质量。
参考文献:
[1]HALLE F,OLDEMAN R A A.Essai surl' Architecture etla Dynam iquede Croissance des Arbres Tropicaux[M].Parix:Masson,1970.
[2]孙书存,陈灵芝.不同生境中辽东栎的构型差异[J].生态学报,1999,19(3):359-364.
SUN S C,CHEN L Z.Architectural analysis of crown geometry inQuercusliaotungensis[J].ActaPhytoecologicaSinica,1999,23(5):433-440.
[3]White J.The plant as a metapopulation[J].AnnualReviewofEcologyandSystematics,1979,10(1):109-145.
[4]陈波,达良俊.栲树不同生长发育阶段的枝系特征分析[J].武汉植物学研究,2003,21(3):226-231.
CHEN B,DA L J.Branching pattern ofCastanopsisfargesiias affected by growth and development phases[J].JournalofWuhanBotanicalResearch,2003,21(3):226-231.
[5]ADOLFO R,ANDREA P,SILVIA C,etal.The role of tree architecture in super high density olive orchards[J].ScientiaHorticulturae,2013,161:24-29.
[6]武高林,杜国祯.植物形态生长对策研究进展[J].世界科技研究与发展,2007,29(4):47-51.
WU G L,DU G Z.Advances in plant morphological growth strategy[J].WorldScienceandTechnologyResearchandDevelopment,2007,29(4):47-51.
[7]李俊清,臧润国,蒋有绪.欧洲水青冈(FagussylvaticalL.)构筑型与形态多样性研究[J].生态学报,2001,21(1):151-155.
LI J Q,ZANG R G,JIANG Y X.Review on studies of architecture and morphological diversity forFagussylvaticaL.[J].ActaEcologicaSinica,2001,21(1):151-155.
[8]康永祥.黄土高原辽东栎林群落生态研究[D].陕西杨陵:西北农林科技大学,2012.
[9]朱志诚.关于秦岭及陕北黄土高原区辽东栎林的初步研究[J].植物生态学与地植物学丛刊,1982,6(2):95-104.
ZHU Z C.Primary study ofQuercusLiaotongensisforest in Qingling mountains and Loess Plateau [J].ActaPhytoecoloicaEtGeobotanica,1982,6(2):95-104.
[10]PRZEMYSLAW PRUSINKIEWICZ,PIERRE BARBIER DE REUILLE.Constraints of space in plant development[J].JournalofExperimentalBotany,2010,61(8):2 117-2 129.
[11]WHITNEY.The Bifurcation Ratio as an Indicator of adaptive strategy in woody plant species[J].BulletinoftheTorreyBotanicalClub,1976,103(2):67-72.
[12]Luna B.Leopold,Trees and streams:the efficiency of branching patterns[J].JournalofTheoreticalBiology,1971,31(2):339-354.
[13]BARKER S B,CUMMING G,HORSFIELD K.Quantitative morphometry of the branching structure of trees.JournalofTheoreticalBiology,1973,40(1):33-43.
[14]DAVID A,STEINGRAEBER,LAWRENCE J,etal.Variation of shoot morphology and bifurcation ratio in sugar maple (Acersaccharum) saplings[J].AmericanJournalofBotany,1979,66(4):441-445.
[15]孙书存,陈灵芝.辽东栎植冠的构型分析[J].植物生态学报,1999,23(5):433-440.
SUN S C,CHEN L Z.Architectural analysis of crown geometry in Quercus liaotungensis[J].ActaPhytoecologicaSinica,1999,23(5):433-440.
[16]胡晓静,张文辉,何景峰,等.不同生境栓皮栎天然更新幼苗植冠构型分析[J].生态学报,2015,35(3):788-795.
HU X J,ZHANG W H,HE J F,etal.Architectural analysis of crown geometry ofQuercusvariablisBL.natural regenerative seedlings in different habitats[J].ActaEcologicaSinica,2015,35(3):788-795.
[17]余碧云,张文辉,胡晓静,等.栓皮栎地上部分构型及生物量分配[J].应用生态学报,2015,26(8):2 265-2 272.
YU B Y,ZHANG W H,HU X J,etal.Aboveground architecture and biomass distribution ofQuercusvariabilis[J].ChineseJournalofAppliedEcology,2015,26(8):2 265-2 272.
[18]伊力塔,韩海荣,程小琴,等.灵空山林区辽东栎(Quercusliaotungensis)种群空间分布格局[J].生态学报,2008,28(7):3 254-3 261.
YI L T,HAN H R,CHENG X Q,etal.Spatial distribution pattern ofQuercusliaotungensispopulation in Lingkong Mountains[J].ActaEcologicaSinica,2008,28(7):3 254-3 261.
[19]李荣,张文辉,何景峰,等.不同间伐强度对辽东栎林群落稳定性的影响[J].应用生态学报,2011,22(1):14-20.
LI R,ZHANG W H,HE J F,etal.Effect of thinning intensity on community stability ofQuercusliaotungensisforest on Loess Plateau [J].ChineseJournalofAppliedEcology,2011,22(1):14-20.
[20]闫兴富,仇智虎,张嫱,等.种皮和播种深度对辽东栎种子萌发和幼苗早期生长的影响[J].应用生态学报,2014,25(1):53-60.
YAN X F,CHOU Z H,ZHANG Q,etal.Effects of coat and sowing depth on seed germination and early seedling growth ofQuercuswutaishanica[J].ChineseJournalofAppliedEcology,2014,25(1):53-60.
[21]STEINGRAEBER D A,WALLER D M.Non-stationarity of tree branching patterns and bifurcation ratios[J].Proceedings of the Royal Society of London.Series B,BiologicalSciences,1986,228(1 251):187-194.
[22]李永华,卢琦,吴波,等.干旱区叶片形态特征与植物响应和适应的关系[J].植物生态学报,2012,36(1):88-98.
LI Y H,LU Q,WU B,etal.A review of leaf morphology plasticity linked to plant response and adaptation characteristics in arid ecosystems[J].ActaPhytoecologicaSinica,2012,36(1):88-98.
[23]CORNELISSEN J H C.Aboveground morphology of shade-tolerant Castanopsis fargesii saplings in response to light environment[J].InternationalJournalofPlantSciences,1993,154(4):481-495.
[24]DONG M.Morphological responses to local light conditions in clonal herbs from contrasting habitats,and their modification due to physiological integration[J].Oecologia,1995,101(3):282-288.
[25]HOVENDEN M J,VANDER SCHOOR J K.Nature vs nurture in the leaf morphology of Southern beech,Nothofaguscunninghamii(Nothofagaceae)[J].NewPhytologist,2004,161(2):585-594.
[26]PICOTTE J J,RHODE J M,CRUZAN M B.Leaf morphological responses to variation in water availability for plants in thePiriquetacarolinianacomplex[J].PlantEcology,2009,200(2):267-275.
[27]PICOTTE J J,ROSENTHAL D M,RHODE J M,etal.Plastic responses to temporal variation in moisture availability:consequences for water use efficiency and plant performance[J].Oecologia,2007,153(4):821-832.
[28]WRIGHT I J,REICH P B,CORNELISSEN J H C,etal.Assessing the generality of global leaf trait relationships[J].NewPhytologist,2005,166(2):485-496.
[29]TAKAHASHI K, Y MIKAMI.Crown architecture and leaf traits of understory saplings ofMacarangasemiglobosain a tropical montane forest in Indonesia[J].PlantSpeciesBiology,2008,23(3):202-211.
[30]OSADA N,TAKEDA H.Branch architecture,light interception and crown development in saplings of a plagiotropically branching tropical tree,Polyalthiajenkinsii(Annonaceae)[J].AnnalsofBotany,2003,91(1):55-63.
[31]李轩然,刘琪璟,蔡哲,马泽清.千烟洲针叶林的比叶面积及叶面积指数[J].植物生态学报,2007,31(1):93-101.
LI X R,LIU Q J,CAI Z,etal.Specific leaf area and leaf area index of conifer plantations in Qianyanzhou station of subtropical China[J].ChineseJournalofPlantEcology,2007,31(1):93-101.
(编辑:潘新社)
Architectural Analysis of Crown Geometry of Saplings ofQuercusliaotungensisForest in Different Habitats
LI Gang,ZHANG Wenhui*,YU Shichuan,XING Zhongli,HUANG Caizhi
(Education of Ministry Key Laboratory of Environment and Ecology in West China,Northwest A&F University,Yangling,Shaanxi 712100,China)
Abstract:We implemented typical sampling method,in order to illustrate the crown architectural variation of Quercus liaotungensis saplings in different habitats and understand the response and adaptation mechanisms of plant to different environment conditions.The characteristics of lateral branch and leaf,as well as their spatial distribution for Q.liaotungensis saplings,which grew in three different habitats (under story,forest gap and forest edge) in Huanglong mountain of Loess Plateau,were investigated.The results showed that:(1)the crown morphological features of Q.liaotungensis saplings changed in response to three kinds of habitats,the crown of understory is simple and the crown of the other habitats are rich.(2)The tree height and the height under branch declined and the diameter increased from understory,forest gap to forest edge.The crown width,crown area,crown ratio of the saplings increased first and then decreased,and the variance under 3 habitats is significant.Overall bifurcation ratio,gradually branching ratio,branch diameter ratio of saplings firstly increased then decreased from understory,gaps to forest edge.(3)Under three kinds of habitats,the length,diameter and angle declined with the tree height increased,and the variance is not significant.The first-older branches were mainly in the top area of the canopy and in the other two habitats were mainly in the middle of the canopy.(4)Leaf length,leaf width,leaf area and leaf area ratio decreased,while the average leaf number,total leaf area and LAI firstly increased and then decreased,from understory,gaps to forest edge.Compared with the other two habitats,the leaves of understory were mainly in the top of the canopy.(5)Aboveground biomass of saplings,the trunk biomass of understory accounted for 83%,branch and leaf biomass account for only 17%.Under the gaps and forest edge habitats,the biomass of various parts was different,but the proportion was basically the same.The biomass of trunk accounted for 66%,and biomass of branch and leaf accounted for about 34%.Under three habitats,the sapling architecture of forest gap was better than that of the other two habitats.In the future,appropriate thinning techniques should be used to increase the number of the forest gap and provide favorable conditions for regeneration and structure optimization of Q.liaotungensis forests.
Key words:Quercus liaotungensis;sapling;crown architecture;biomass distribution
中图分类号:Q948.12
文献标志码:A
作者简介:李罡(1989-),男,在读硕士研究生,主要从事森林培育、天然林保护理论与实践研究。E-mail:ligang1989_1@126.com*通信作者:张文辉,教授,主要从事植物生态、生物多样性保护与利用研究。E-mail:zwhckh@163.com
基金项目:陕西省科技统筹创新工程计划(2014KTCL02-04)
收稿日期:2015-11-27;修改稿收到日期:2016-03-09
文章编号:1000-4025(2016)03-0588-08
doi:10.7606/j.issn.1000-4025.2016.03.0588
