海南省万宁市野生荔枝资源种群特征研究
2016-05-06张萱蓉杨小波李东海林泽钦李苑菱刘子金
张萱蓉,李 丹,杨小波*,李东海,林泽钦,李苑菱,刘子金
(1 海南大学 园艺园林学院,海口 570228;2 热带作物种质资源保护与开发利用教育部重点实验室,海口 570228)
海南省万宁市野生荔枝资源种群特征研究
张萱蓉1,2,李丹1,2,杨小波1,2*,李东海1,2,林泽钦1,2,李苑菱1,2,刘子金1,2
(1 海南大学 园艺园林学院,海口 570228;2 热带作物种质资源保护与开发利用教育部重点实验室,海口 570228)
摘要:野生荔枝是荔枝(Litchi chinensis Sonn.)进行品种改良与创新的重要基因库,该研究通过对海南万宁自然保护区内、外野生荔枝资源进行实地调查,并用空间分布格局、种群径级结构、静态生命表、存活曲线等方法进行比较分析,以揭示野生荔枝种群在保护区内、外的分布规律及其种群动态趋势,为野生荔枝资源的保护提供依据。结果表明:(1) 海南万宁自然保护区内、外野生荔枝种群均为聚集分布,且非自然保护区聚集程度比自然保护区的聚集程度更为明显。(2)径级结构分析发现,自然保护区内野生荔枝种群结构为增长型,种群生命期望值整体上基本随径级增加呈递减趋势,存活曲线呈近直线型,说明自然保护区内的野生荔枝种群处于相对稳定的状态;而自然保护区外的野生荔枝种群为衰退型。(3)群动态预测结果也显示,自然保护区内外的野生荔枝种群的动态变化指数(V(pi)′)以及随机干扰风险极大值均趋于零,但自然保护区外的值均大于区内,说明保护区外受到的敏感性比自然保护区内更强,即保护区外野生荔枝种群的生存环境已受到严重的人为干扰威胁。研究认为,在自然保护区外,野生荔枝种群幼树的严重缺失不仅与野生荔枝种群自身的更新机制有关,更重要的是生存环境的破坏和人为砍伐导致野生荔枝种群结构发生变化,野生荔枝林面积下降。
关键词:野生荔枝;空间分布格局;种群径级结构;静态生命表;存活曲线
荔枝(LitchichinensisSonn.)属无患子科(Sapindaceae)荔枝属(Litchi)常绿乔木,是中国南方亚热带地区广泛栽培的特产果树[1]。中国有着2000多年的荔枝栽培历史,在不断的发展过程中,已经孕育出可以适应各种不同环境条件的荔枝品种,形成了丰富的种质资源[2]。对种质资源进一步的研究可促进荔枝产业不断的向前发展[2]。野生资源向来受到国内外育种学者的重视,因其具有改良品种、提高产量、增强抗逆性等的作用[3],在荔枝育种、荔枝种质创新及荔枝栽培生产有着不可估量的作用。野生荔枝在中国主要分布在广东省和海南省[4],曾经,野生荔枝也是森林中的优势树种之一,但由于多年林业开采,特别是20世纪80年代林业的采伐,野生荔枝资源已遭到严重的破坏,分布范围及数量越来越少。在《国家重点保护野生植物名录(第一批)》中,野生荔枝已被列为国家二级保护植物,成为渐危树种[5]。
对于野生荔枝的研究,国外鲜有报道。在国内,野生荔枝的研究不再是只停留资源调查和形态观察方面,已经逐步向分子生物学方向发展。海南省作为野生荔枝主要分布地之一,较早时期便有学者对其进行相关的研究工作,1921年,陈焕镛提出海南岛存在野生荔枝[6]。1964年,徐祥浩等[7]多次调查后确认了海南存在野生荔枝,1983年,徐祥浩[8]为分析野生和栽培种的差异,从树干、叶片、果实和生态条件四个方面着手分析,并将野生荔枝命名为新变种。1983年,傅玲娟等[4]对海南岛野生荔枝多种类型进行调查分析,从生态条件和植物学性状上列出了野生荔枝与栽培品种的不同点。此外,分子生物学技术的飞速发展,分子标记技术被更多的应用在荔枝遗传多样性及亲缘关系研究。如姚庆荣等[9]对37份海南霸王岭野生荔枝群体、7份琼山半野生群体和22份栽培品种进行SSR标记并分析其遗传多样性;邓穗生等[10]应用RAPD标记对海南霸王岭的60株野生荔枝进行了聚类分析及归类;罗海燕等[11]对海南的野生荔枝、半野生荔枝和栽培荔枝共96份样品材料进行遗传多样性及亲缘关系分析,初步鉴定了部分同物异名和同名异物品种,并提出了划定海南野生荔枝保护区的建议,取得了较大进展。
长期以来,种群结构与分布格局—直被看成是种群生态学的核心问题[12]。种群结构和空间分布结局的研究对阐明生态系统的形成与维持、群落的稳定性与演替规律、种群的动态特征、种群与环境之间的互作过程以及种群更新具有极为重要的意义[13]。目前对野生荔枝群落、种群的相关研究并不多,如刘冰浩等[14]和韦阳连等[15]分别对广西玉林市博白县与广东廉江市谢鞋山的野生荔枝种群进行了研究。但是,还没有文献对自然保护区内外的野生荔枝种群特征的比较分析进行详细的研究报道,所以本研究以万宁市为例,对分布在自然保护区内和自然保护区外的野生荔枝种群动态特征进行比较研究,揭示野生荔枝种群在保护区内外的分布规律及其种群动态趋势,旨在为海南万宁市野生荔枝资源的保护提供参考依据。
1研究区概况
万宁市位于海南岛东南部,E110°0′~110°34′,N18°35′~19°06′之间。该地区西部、西南及西北部峰峦起伏,连绵不断,海拔在500~1 200 m之间[16]。本地区属热带季风气候,积温高、雨量充足,年平均气温为24.3 ℃,最冷月的平均气温为18.7 ℃,最热月的平均气温为28.5 ℃[17]。万宁东部主要地貌是平原台地,土壤以玄武岩厚层粘质土、花岗岩中层砂砾粘壤土、砂页岩中层砂砾壤质土为主;西部主要地貌是低山丘陵,土壤以花岗岩厚层砂质粘壤土为主[18]。主要植被类型为热带山地雨林、热带次生林、灌丛和人工林等4个植被类型,种类组成较复杂,主要有壳斗科(Fagaceae)、梧桐科(Sterculiaceae)、龙脑香科(Dipterocarpaceae)、桃金娘科(Myrtaceae)、樟科(Lauraceae)、茜草科(Rubiaceae)、棕榈科(Palmae)、大戟科(Euphorbiaceae)等;除了自然保护区外,万宁境内天然林还分布在双决岭、寒门岭、黑岭、尖岭和农村周边,其植物种类较为简单,主要以无患子科(Sapindaceae)、棕榈科、桃金娘科、大戟科和桑科(Moraceae)等为主。
2研究方法
2.1野外调查方法
对海南省万宁市境内7个自然保护区(海南大洲岛国家级自然保护区、海南礼纪青皮林省级自然保护区、海南六连岭省级自然保护区、海南南林省级自然保护区、海南尖岭省级自然保护区、海南上溪省级自然保护区以及海南加新省级自然保护区)和非自然保护区的野生荔枝资源进行全面调查。
调查期间,当野生荔枝散生或团状分布,且连片分布,面积较大时采用“样方法”;当分布区域狭窄,分布面积小,种群数量稀少而可以直接计数且有相关资料,其分布点、范围和资源都较清楚时采用“实测法”。设置野生荔枝样地时,每个样地为20 m×20 m,并记录样地地理坐标、海拔、坡度、坡向等生境指标(表1),对所设立样方内的所有野生荔枝进行调查,测定其胸径、树高、冠幅、坐标等。实测即对野生荔枝进行定位,调查内容与“样方法”相同。本次调查中野生荔枝有17个实测点,设置样方23个,总面积为9 200 m2。
2.2数据分析
2.2.1空间分布格局分析方法本研究参考张金屯[19]、闫淑君等[20]的种群空间分布格局方法,利用样方数据(20 m×20 m),采用扩散系数法(C)、负二项式参数(K)、Cassie指标(Ca)、格林指数(GI)、平均拥挤度(m*)和聚块性指标(PAI)等指标分别测定种群聚集强度,用Poisson分布、负二项分布检验种群分布格局的拟合[21-24]。
2.2.2径级划分划分龄级是研究种群生命表、存活曲线等的关键[25],虽然种群的龄级和径级有所不同,但同一环境下,同一树种的龄级和径级对环境的反应规律具有一致性[26]。对于数量较少的濒危树种,在很难获取其年龄的情况下,大多数学者采用径级结构代替年龄级结构[27-28],本研究参考其他学者的划分方法[29-32],用立木级结构代替年龄结构分析野生荔枝种群,并结合野生荔枝生物学特性和生长情况,将野生荔枝进行以下划分:DBH<5 cm,按其树高划分为<33 cm的为Ⅰ级幼苗阶段,≥33 cm的为Ⅱ级幼树阶段;DBH≥5 cm以上,按其DBH大小分级:5 cm≤DBH<10 cm为Ⅲ级小树阶段,10 cm≤DBH<25 cm为Ⅳ级中树阶段,25 cm≤DBH<40 cm为Ⅴ级大树阶段,DBH≥40 cm为Ⅵ级老树阶段。

表1 各样地野生荔枝种群基本概况
2.2.3编制种群静态生命表生命表是判定种群趋势的重要指标,可反映种群现实状况,种群与环境的竞争关系[33-34]。按照静态生命表的编制方法,综合野外调查数据编制野生荔枝种群静态生命表。统计各径级株数,依据野外调查各径级存活数(Ax),计算出标准化存活数(Lx)、死亡数(dx)、死亡率(qx)、从第x径级起超过x径级的有活个体总数(Tx)、生命期望(ex)、消失率(Kx)等编制种群生命表[35-36],并绘制种群的存活曲线,Deevey将个体存活概率随相对年龄的变化分为3个基本模式,即Ⅰ型、Ⅱ型和Ⅲ型。Ⅰ型:凸形曲线,显示年轻个体存活率较高;Ⅱ型:对角线,显示各年龄阶段死亡率相等;Ⅲ型:凹形曲线,表明幼年期死亡率很高[37-39]。
2.2.4种群动态数量化方法种群动态数量化方法采用文献[40]的方法,定量描述种群动态:
(1)
(2)
式中,Vn为种群从n到n+1级的个体数量变化;Vpi为整个种群结构的数量变化动态指数;Sn、Sn+1分别为第n和n+1级种群个体数;k为种群大小级数量。式(2)仅适用于不考虑未来外部环境干扰的种群结构动态的比较,当考虑未来的外部干扰时,则种群结构动态还与大小级数量(k)及各大小级个体数(S)两因素相关,因此将式(2)修正为:
(3)
(4)
式中,Vn、Vpi和Vpi′分别反映种群个体数量的增长、衰退和稳定的动态关系。
2.2.5种群数量动态的时间序列分析时间序列分析是时间数列预测和回归预测方法的综合形式,该方法不用于分析因果关系,侧重于根据时间序列过去的变化和变动规律推测今后的趋势[41],本研究采用时间序列分析的一次移动平均法对野生荔枝的种群龄级结构进行模拟和预测[42-43]。
3结果与分析
3.1野生荔枝种群分布格局
根据所调查的自然保护区17个样方,非自然保护区6个样方的资料,计算出万宁野生荔枝种群的格局指数和各聚集强度指标(表2)。从表2可知,自然保护区与非自然保护区的野生荔枝与格局指数(C)值分别为1.852 3、4.097 7,均大于1,t值分别为2.410 7、4.897 8,表明无论是自然保护区还是非自然保护区,野生荔枝种群都趋于聚集分布;从各聚集强度指标来看,而负二项参数K值分别为3.243 8、1.560 3,数值小于8且大于0,种群为聚集分布,且非自然保护区K值小于自然保护区K值,说明非自然保护区野生荔枝种群的聚集程度更大;Cassie指标(Ca)值和丛生指数(GI)值均大于零,聚块性指标(PAI)值分别为0.638 2、1.206 9,自然保护区的野生荔枝种群聚块性指标小于1,呈现出均匀分布,而非自然保护区大于1,为聚集分布。综合格局指数和各聚集强度指标值,万宁野生荔枝群落总体为聚集分布,且非自然保护区聚集程度比自然保护区聚集程度更为明显。
3.2野生荔枝种群结构特征
以野生荔枝的各个年龄级植株数为纵坐标,各个年龄级为横坐标,绘制万宁野生荔枝年龄级结构图。从图1可知,万宁的野生荔枝种群年龄结构呈现不规则型,各个年龄级均有分布。其中,自然保护区内的野生荔枝第Ⅱ径级个体数量最多,占总体数量的39.77%,其次是第Ⅰ径级、第Ⅲ径级。第Ⅵ径级个体数最少,占总体数量的2.27%;而非自然保护区的野生荔枝第Ⅰ径级个体数量最多,占总体数量的34.48%,其次是第Ⅵ径级,第Ⅴ径级和第Ⅵ径级的个体数一样,第Ⅲ径级的个体数量最少。从总体来看,自然保护区内的野生荔枝种群幼龄个体较多,属于增长型种群;而非自然保护区的野生荔枝虽然在幼苗时期的个体数量最多,但到了幼树时期数量急剧减少,到了中年和老年时期的种群数量增加且相对稳定,可见非自然保护区的野生荔枝种群呈现出衰退趋势。
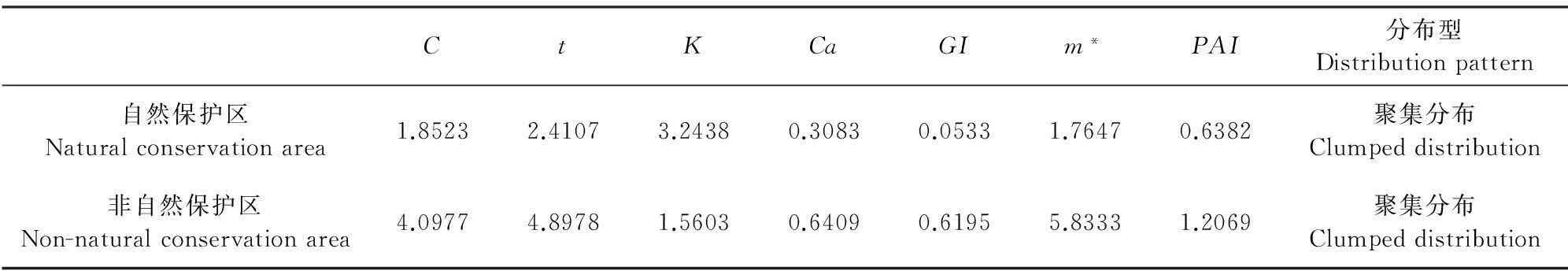
表2 自然保护区与非自然保护区的野生荔枝种群分布格局
根据种群动态量化方法分析野生荔枝种群相邻年龄级个体变化,以对种群动态进行更加客观精确的评价。结果显示,自然保护区的野生荔枝种群相邻各级间个体数量变化动态指数V1小于0,说明Ⅰ龄级个体数目少于Ⅱ龄级个体数目;而非保护区内中,野生荔枝种群相邻各级间个体数量变化动态指数V3小于0,说明Ⅲ龄级个体数目少于Ⅳ龄级个体数目;进一步计算不同保护情况下野生荔枝种群的总体数量动态数量变化指数Vpi(在不考虑外界的环境干扰时)和Vpi′(受外界随机干扰),结果显示,2种类型下野生荔枝种群的总体数量动态变化指数Vpi′均小于Vpi但都大于0,其中Vpi′更趋于0(表3),随机干扰风险极大值(P极大)分别为0.083(自然保护区)、0.167(非自然保护区),说明2种保护状况下的野生荔枝种群总体均表现为增长型,但增长缓慢,对外界干扰具有较高的敏感性且非保护区更为敏感。

Ⅰ级(幼苗).DBH<5 cm,树高<33 cm;Ⅱ级(幼树).DBH<5 cm,
3.3野生荔枝种群静态生命表及存活曲线
从静态生命表(表4)可以看出,自然保护区内野生荔枝种群第Ⅰ龄级的个体数少于第Ⅱ龄级的个体数,导致其死亡率为负值;但是从整体来看,野生荔枝种群生命期望随着年龄级的增加而呈递减趋势,符合种群生物学特性,但在第Ⅱ年龄级存在波动现象;野生荔枝生命期望值在第Ⅰ年龄级达到最大,明显高于其它年龄级的生命期望,而生命期望值(ex)反映x年龄级内个体数的平均生长状况良好,种群呈增长趋势。而在非自然保护区,由于第Ⅲ年龄级个体数比第Ⅳ年龄级少,出现了死亡率为负的现象,但从其生命期望值可看出,非自然保护区野生荔枝种群的生命期望值先升高达到—个较高的峰值后再逐渐降低,较高的生命期望值出现在年龄级Ⅱ和年龄级Ⅲ,之后逐渐减小,说明非保护区野生荔枝种群第Ⅱ年龄级和第Ⅲ年龄级这2个阶段野生荔枝的生存质量较高,而到了较高年龄级时野生荔枝的生存质量下降。
本研究以存活量为纵坐标,龄级为横坐标绘制野生荔枝种群存活曲线。从图2可知,自然保护区内的野生荔枝种群的存活曲线存在波动,第Ⅰ径级的存活数少于第Ⅱ径级,但之后随着径级的递增,单调递减。其中,第Ⅰ径级与第Ⅱ径级幼龄阶段的存活率最高,表现出丰富的后备资源,从第Ⅱ径级到第Ⅲ径级存活率有较大幅度的下降;而非保护区野生荔枝种群的存活曲线呈现出“V”型,先下降后升高的趋势;通常检验估算的存活状况是符合Deevey-Ⅱ型曲线还是符合Deevey-Ⅲ型曲线时,采用两种数学模型进行检验,即指数方程式Nx=N0e-bx用以描述Deevey-Ⅱ型存活曲线,幂函数式Nx=N0x-bx描述Deevey-Ⅲ型存活曲线[44]。这里采用上述两种模型对植被类型为季雨林和热带雨林的野生荔枝种群进行存活曲线类型的检验,经建立其相应模型得到,自然保护区:Nx=2 450.1e-0.499x(R2=0.857 9),Nx=1 729x-1.275(R2=0.702 6);非自然保护区:Nx=341.07e0.030 7x(R2=0.004 7),Nx=450.27x-0.155(R2=0.015 2)。自然保护区中,指数模型的R2值大于幂函数模型的R2值,因此可以认为自然保护区的野生荔枝种群存活曲线曲线更加符合Deevey-Ⅱ型,呈对角线型,各年龄级种群有相近的死亡率;而非自然保护区的幂函数模型的R2值大于指数模型的R2值,存活曲线更加符合Deevey-Ⅲ型,呈凹形曲线,表明幼年期死亡率很高。
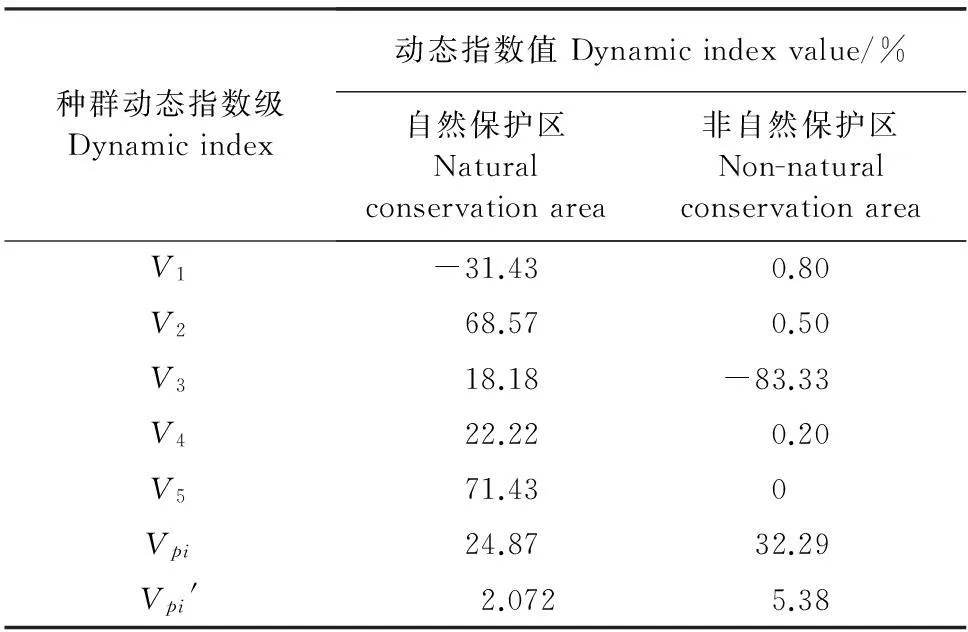
表3 自然保护区与非自然保护区的野生荔枝

表4 自然保护区与非自然保护区的野生荔枝种群静态生命表
注:x.龄级Age class;Ax.x龄级开始时的实际存活数;lx.x龄级开始时的标准化存活数;dx.从x到x+1龄级的标准化死亡数;qx.x龄级的个体死亡率;Lx.从x到x+1龄级的平均存活个体数;Tx.x龄级及以上各龄级的存活个体数;ex.进入x龄级个体的平均生命期望;Kx.种群消失率。
Note:x.Age class;Ax.Real survival individual number at beginning ofxage class;lx.xStandardized survival number at beginning of age class;dx.Standardized death number fromxtox+1 age class;qx.Individual mortality rate atxage class;Lx.Average survival individual number fromxtox+1 age class;Tx.Survival individual number fromxto higher age class;ex.Average expectative life of individuals intoxage class;Kx.Disappearance rate of population.
3.4时间序列预测
野生荔枝种群数量动态时间序列预测以野生荔枝种群各龄级株数为原始数据,按照一次平均推移法预测出各龄级在未来2、4个龄级时间后的个体数。野生荔枝的时间序列分析(表5)表明:在未来的2、4个龄级后,自然保护区的野生荔枝种群从Ⅳ龄级之后的个体数呈现增加的趋势,表明野生荔枝幼龄个体占的比例高,使得种群生长中后期的个体数量得到一定的补充。但也存在一定的波动,Ⅱ龄级在经历了2级时间后,个体数由原来的35株下降到30株,说明幼苗的不足会使幼树的数量有所缩减,但幅度不大;而非自然保护区的野生荔枝种群在Ⅳ级之后;种群的数量有所下降,虽然Ⅱ、Ⅲ龄级在经历2级时间后数量有小幅度的提升,但因受外界环境干扰较大,幼龄级的植株无法转换到下一阶段,使得种群呈现出衰退的趋势。因此,从种群动态预测结果来看,自然保护区的野生荔枝种群由于幼年个体数相对丰富,种群维持有较充足的后备资源,中树、大树阶段的个体数能够在现有的基础上得到补充,如果能保持现有野生荔枝种群分布的植株和生境斑块,野生荔枝种群具备较好的恢复潜能;但非自然保护区的野生荔枝种群因为外界的影响,如果不采取适当的保护措施,种群未来必然趋于衰退。

图2 自然保护区与非自然保护区的野生荔枝种群存活曲线
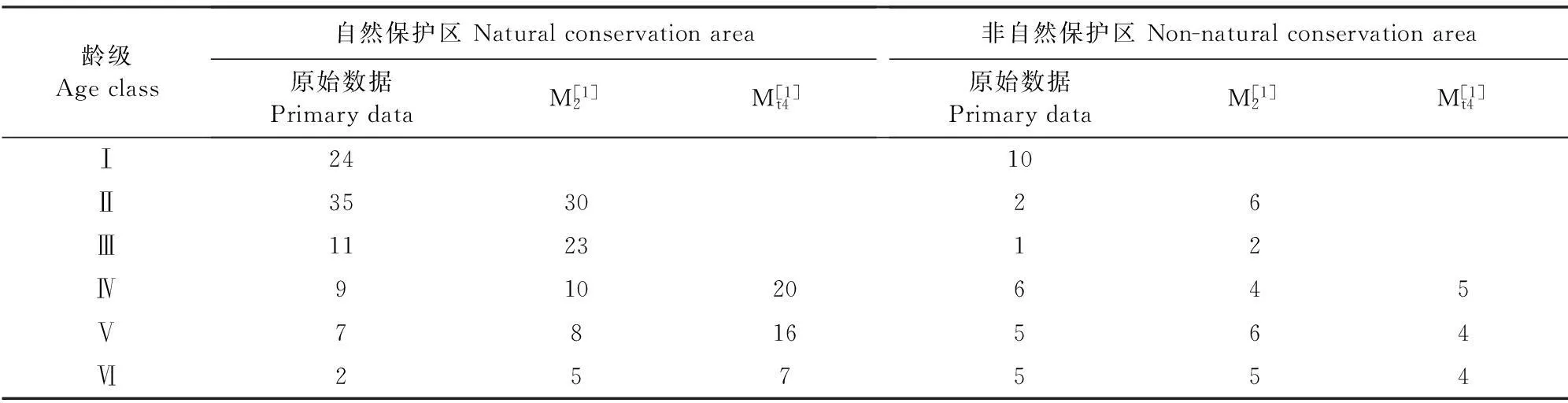
表5 自然保护区内外的野生荔枝种群龄级结构动态变化的时间序列预测
4讨论
4.1野生荔枝种群空间分布格局
野生荔枝生活环境复杂,基因资源丰富多样,对栽培荔枝的起源演化研究中具有十分重要的作用。种群格局形成,除了自身的特性,还与群落环境相关,群落环境包含生物因子和非生物因子。种群分布格局及其动态,是由于种群生物学特性和生境条件相互作用而形成的[45]。这次研究调查结果显示,无论在自然保护区或非自然保护区,野生荔枝种群都呈聚集分布,但是,对比两种不同的生境,自然保护区格局指数(C)值1.852 3,聚集强度指标中,负二项参数K值3.243 8,Cassie指标(Ca)0.308 3,Green指数(GI)0.053 3,平均拥挤度指数1.764 7,聚块性指标0.638 2而非自然保护区格局指数(C)值4.097 7,聚集强度指标中,负二项参数K值1.560 3,Cassie指标(Ca) 0.640 9,Green指数(GI)0.619 5,平均拥挤度指数5.833 3,聚块性指标1.206 9。综合格局指数和各聚集强度指标值,万宁自然保护区内外的野生荔枝种群均为聚集分布,但是非自然保护区聚集程度比自然保护区聚集程度更为明显。从野生荔枝自身特性分析,在自然保护区内,由于林下和小林窗的光强度低,且野生荔枝种子大,能为幼苗的存活提供—定的营养,有利于叶子的形成以及其他的生理特性,也有利于低光下生长,从而能够忍耐不良光环境条件[46]。光强度会影响苗木叶绿素含量和RuBP羧化酶活性,不同光环境中的同—物种的叶片总蛋白含量基本—致,但是单位酶活性有很大的变化,如果长时间的暴晒,幼苗的叶绿素会被漂白[47],从而使幼苗无法存活。在非自然保护区,见光度高,野生荔枝的幼苗长时间的接受光照,从而出现“饿死”的现象,再加上人为干扰,野生荔枝种群更新能力下降,限制了野生荔枝种群的分布范围。
4.2野生荔枝种群动态特征
植物种群表现结构是种群发育过程中每个个体实现其增长机会的一种表达,也是对立地条件优劣及植物对环境适应性的反映[48]。本次调查研究发现自然保护区内野生荔枝种群径级结构显示野生荔枝种群结构为增长型,种群生命期望值整体上基本随径级增加呈递减趋势,存活曲线呈近直线型,说明自然保护区内的野生荔枝种群处于相对稳定的状态。这与海南昌江县野生荔枝种群结构的结果一致。周文嵩等[49]在对海南昌江县野生荔枝种群结构分析结果显示,野生荔枝种群的存活曲线呈对角线型,接近于Deevey-Ⅱ型;而非自然保护区的野生荔枝种群径级结构显示野生荔枝种群结构呈衰退型,存活曲线接近Deevey-Ⅲ型,幼龄个体具有较高的死亡率,这与刘冰浩等[14]对广西野荔枝种群的研究结果—致,刘冰浩等研究发现,广西地区野荔枝种群幼苗个体较少,种群处于衰退状态;韦阳连等[15]对广东廉江野生荔枝种群的研究也得到类似的结果。另外,自然保护区内的野生荔枝种群的动态变化指数以及随机干扰风险极大值均趋于零,说明种群表现为增长型,但对外界有较高的敏感性,种群动态预测结果表明,自然保护区的野生荔枝种群由于幼年个体数相对丰富,种群维持有较充足的后备资源,中树、大树阶段的个体数能够在现有的基础上得到补充,如果能保持现有野生荔枝种群分布的植株和生境斑块,野生荔枝种群具备较好的恢复潜能;而非自然保护区的野生荔枝种群的动态变化指数以及随机干扰风险极大值虽趋于零,但大于自然保护区的值,说明其受到的敏感性比自然保护区的更强且种群动态预测结果也显示如果不采取适当的保护措施,非自然保护区的种群未来必然趋于衰退。分析其两者不同的原因可能是:生境破坏和人为砍伐导致野生荔枝种群结构发生变化,在自然保护区内的野生荔枝由于受到的人为干扰较低,种群结构为稳定型,有丰富的幼苗储备,种群更新能力强,整个种群处于稳定发展阶段,幼苗的补充对群落起着决定性作用,使得野生荔枝种群得以恢复和发展;而在非自然保护区,幼树的严重缺失不仅与野生荔枝种群自身的更新机制有关,更重要的是生存环境的破坏,非自然保护区没有得到应有的保护,人为干扰较为严重,特别是近年来,当地居民大量毁林种植经济作物,生境破坏和人为砍伐导致野生荔枝种群结构发生变化,野生荔枝林面积迅速下降,野生荔枝的生存受到极大的威胁,导致荔枝种群更新能力不足,受损群体难以复壮和扩展。
分析结果表明,野生荔枝种群的数量不断在减少,目前,野生荔枝已被列为国家二级保护植物,作为荔枝育种的重要种质资源,需保护和利用野生荔枝中丰富的基因资源。针对野生荔枝种群的生存现状,提出以下的建议措施:(1)应进一步全面普查海南现有的野生荔枝资源,摸清野生荔枝资源在海南的分布数量、类型、生态。同时建立有代表性的野生荔枝原位保护区。(2)提高群众的保护意识,大力宣传野生荔枝的科研价值和开发利用的重要意义,使更多的群众加入到保护队伍中来。(3)重点加强对野生荔枝种质创新利用的研究,使优良种质资源在推广应用中得到有效保护和发展。
参考文献:
[1]庄光辉,胡福初,何舒,等.海南原生荔枝种质资源收集保存及主要性状评价[J].热带农业科学,2011,31(11):28-35.
ZUANG G H,HU F C,HE S,etal,Collection and conservation of nativeLitchiGermplasmsin Hainan and evaluation of their main characters[J].ChineseJournalofTropicalAgriculture,2011,31(11):28-35.
[2]陈业渊,海南荔枝种质资源考察收集、鉴定评价及分析[D].海南:海南大学,2011.
[3]诸葛莹,黄吉森,梁冠陵.百色地区甘蔗野生资源植物考察简报[J].广西农业科学,1986,5:18-19.
ZHUGE Y,HUANG J S,LIANG G L.Wild plant resources ofSugarcaneinvestigation briefing in Baise region[J].JSouthAgric,1986,5:18-19.
[4]傅玲娟,袁沛元.广东海南野生荔枝多种类型的发现[J].中国果树,1983,4:21-25.
FU L J,YUAN P Y.The discovery of variety types of wild litchi in Guangdong,Hainan[J].ChinaFruits,1983,4:21-25.
[5]姜成东,蔡胜忠,肖翔,等.海南两个自然保护区野生荔枝遗传多样性研究[J].中国农学通报,2009,25(9):282-286.
JIANG C D,CAI S Z,XIAO X,etal.Studies of allozyme heredity diversity of Wild Litchi in two Hainan Nature Protection Areas[J].ChineseAgriculturalScienceBulletin,2009,25(9):282-286.
[6]吴淑娴.中国果树志.荔枝卷[M].北京:中国林业出版社,1998:1-68.
[7]徐详浩,彭浙,薛日辉,等.关于野生荔枝的发现[J].园艺学报,1964,3(4):415-416.
XU X H,PENG Z,XUE R H,etal.About the discovery ofwildLitchi[J].ActaHorticulturaeSinica,1964,3(4):415-416.
[8]徐详浩.荔枝的新变种——野生荔枝[J].华南农学院学报,1983,4(2):32-34.
XU X H.A new variety ofLitchichinensissonn.-Wildlichee[J].JournalofSouthChinaAgriculturalUniversity,1983,4(2):32-34.
[9]姚庆荣.用SSR标记对中国荔枝(LitchichinensisSonn)野生种质资源的遗传多样性分析[D].兰州:甘肃农业大学,2004.
[10]邓穗生,陈业渊,张欣.应用RAPD标记研究野生荔枝种质资源[J].植物遗传资源学报,2006,7(3):288-291.
DENG S S,CHEN Y Y,ZHANG X.Identification of wild Litchi (LitchichinensisSonn.var.euspontaneaHsue) germplasm resources using RAPD markers[J].JournalofPlantGeneticResources,2006,7(3):288-291.
[11]罗海燕.海南野生荔枝种质资源遗传多样性及与半野生、栽培荔枝亲缘关系的ISSR分析[D].海南:华南热带农业大学,2007.
[12]吴晓莆,郑豫,马克平.北京东灵山地区辽东栎、大叶白蜡和五角枫种群分布格局与动态[J].植物学报,2002,44(2):212-223.
WU X P,ZHENG Y,MA K.Population Distribution and Dynamics ofQuercusliaotungensis,FraxinusrhynchophyllaandAcermonoin Dongling Mountain,Beijing[J].JournalofIntegrativePlantBiology,2002,44(2):212-223.
[13]徐学红,于明坚,胡正华,等.浙江古田山自然保护区甜槠种群结构[J].生态学报,2005,25(3):645-653.
XU X H,YU M J,HU Z H,etal.The structure and dynamics ofCastanopsiseyreipopulation in Gutian Mountain Natural Reserve in Zhejiang,East China[J].ActaEcologicaSinica,2005,25(3):645-653.
[14]刘浩冰,朱建华,潘丽梅,等.广西野生荔枝博白种群生命表分析[J].果树学报,2010,27(3):445-448.
LIU H B,ZHU J H,PAN L M,etal.Analysis on life table ofwildlitchichinensispopulation in Bobai of Guangxi Province[J].JournalofFruitScience,2010,27(3):445-448.
[15]韦阳连,欧阳勤森,钟卫东,等.广东廉江野生荔枝种群结构和分布格局[J].亚热带植物科学,2014,43(1):59-62.
WEI Y L,OUYANG Q S,ZHONG W D,etal.Population Structure and Spatial Pattern ofWildLitchichinensisin Lianjiang,Guangdong Province[J].SubtropicalPlantScience,2014,43(1):59-62.
[16]郑希龙,戴好富,刘寿柏,等.海南黎族药用植物资源调查研究——以万宁市黎族为例[J].中国民族医药杂志,2013,4(4):20-23.
ZHEN X L,DAI H F,LIU S B,etal.A case study on folk medicinal plant resources used by Li Minority in Wanning City,Hainan[J].JournalofMedicine&PharmacyofChineseMinorities,2013,4(4):20-23.
[17]张永夏,张荣京,刑福武,等.海南万宁青皮林群落多样性特征研究[J].2007,27(7):1 454-1 460.
ZHANG Y X,ZHANG R J,XING F W,etal.Community Diversity of theVaticamangachapoiForests in Wanning,Hainan Islands[J].ActaBot.Boreal.-Occident.Sin,2007,27(7):1 454-1 460.
[18]李福燕,李许明,陈柳燕,等.海南万宁市土壤养分状况分析[J].土壤通报,2008,39(6):1 284-1 287.
LI F Y,LI X M,CHEN L Y,etal.The analysis of soil nutrient situations in Wanning of Hainan Province[J].ChineseJournalofSoilScience,2008,39(6):1 284-1 287.
[19]张金屯,孟东平.芦芽山油松-辽东栎林优势树种空间分布格局研究[J].西北植物学报,2006,26(8):1 682-1 685.
ZHANG J T,MENG D P.Spatial distribution patterns of dominant tree species inPinustabulaeformisQuercusliaotungensisForest in Luyashan Mountain China[J].Acta.Bot.Boreal.-Occident.Sin,2006,26(8):1 682-1 685.
[20]闫淑君,洪伟,吴承祯.闽江口琅岐岛朴树种群结构与空间分布格局[J].生态学杂志,2011,30(2):221-226.
YAN S J,HONG W,WU C Z.Population structure and spatial pattern ofCeltissinensison Langqi Island of Minjiang Estuary[J].ChineseJournalofEcology,2011,30(2):221-226.
[21]李晓景,何东进,洪伟,等.天宝岩自然保护区不同海拔天然柳杉种群空间分布格局研究[J].江西农业大学学报,2009,31(3):422-426.
LI X J,HE D J,HONG W,etal.A study on the spatial distribution pattern of NaturalCryptromeriafortune populations at different altitudes in the Tianbaoyan Nature Reserve[J].ActaAgriculturaeUniversitatisJiangxiensis,2009,31(3):422-426.
[22]曹晏宁,史利莎,韩烁,等.科尔沁沙地植被恢复中差不嘎蒿种群不同龄级个体的点格局分析[J].植物学报,2008,25(4):437-442.
CAO Y N,SHI L S,HAN S,etal.Point pattern analysis for a population ofArtemisiahalodendronin a Kerqin Sandlot[J].ChineseBulletinofBotany,2008,25(4):437-442.
[23]上官铁梁,张峰.山西绵山植被优势种群的分布格局与种间联结的研究[J].植物科学学报,1988,6(4):357-364.
SHANGGUAN T L,ZHANG F.Research on the pattern and assocoations between dominants of the vegetation in Mian mountain,Shanxi Province[J].PlantScienceJournal,1988,6(4):357-364.
[24]GIRTIN’S R.Canonical analysis,a review with applications in ecology [M].Berlin:Sprinter Overflag,1985.
[25]何亚平,费世民,蒋俊明,等.不同龄级划分方法对种群存活分析的影响——以水灾迹地油松和华山松种群生存分析为例[J].植物生态学报,2008,32(2):448-455.
HE Y P,FEI S M,JIANG J M,etal.Influences of substituting size variables for age on population survival analysis:a case study ofPinustabulaeformisandP.Armandiiin Qinling Mountain[J].ChineseJournalofPlantEcology,2008,32(2):448-455.
[26]FROST I,RYDIN H.Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests[J].Ecoscience,2000,7(1):38-44.
[27]MA K,LI G Y,ZHU C X,etal.Population structure and distribution patterns of the rare and endangeredArdisiaviolacea(Myrsinaceae)[J].ActaEcologicaSinica,2013,33(2):72-79.
[28]宋超,余琦殷,于梦凡,等.北京地区黄檗种群数量结构及空间分布特征[J].生态杂志,2015,34(11):3 040-3 049.
SONG C,YU Q Y,YU M F,etal,Population structure and spatial distribution characteristics ofPhellodendronamurensein Beijing[J].ChineseJournalofEcology,2015,34(11):3 040-3 049.
[29]Cassie R M.Frequency distribution medal in ecology plant and other organism[J].Anim.Ecol.,1962,31:65-95.
[30]JOHNSON J B.Stand structure and vegetation dynamics of a subalpine tree fen in Rocky Mountain National Park,Colorado[J].JournalofVegetationScience,1997,8(3):337-342.
[31]张维,焦子伟,任艳利,等.新疆野核桃自然保护区不同坡向野核桃种群生命表与波动周期[J].西北植物学报,2015,35(6):1 229-1 237.
ZHANG W,JIAO Z W,REN Y L,etal.Life Tables and Fluctuation Cycle ofJuglansCathayensispopulations on different slope aspects in Wild Walnut Natural Conservation Area of Xinjiang,China[J].ActaBot.Boreal.-OccidentSin,2015,35(6):1 229-1 237.
[32]解婷婷,苏培玺,周紫鹃,等.荒漠绿洲过渡带沙拐枣种群结构及动态特征[J].生态学报,2014,34(15):4 273-4 279.
XIE T T,SU P X,ZHOU Z J,etal.Structure and dynamic characteristics ofCalligonummongolicumpopulation in the desert-oasis ecotone[J].ActaEcologicaSinica,2014,34(15):4 273-4 279.
[33]MOLLES MC.Concept and Applications[M].2nd edition New York:McGraw-Hill Companies.Ecology,2002.
[34]王道亮,李卫忠,曹铸,等.黄龙山天然次生林辽东栎种群空间格局[J].生态学报,2016,36(9):1-10.
WANG D L,LI W Z,CAO Z,etal.Spatial pattern ofQuercuswutaisshanicain natural secondary forest of Huanglong Mountain[J].ActaEcologicaSinica,2016,36(9):1-10.
[35]张兴旺,张小平,郭传友,等.安徽皇藏峪自然保护区青檀种群数量动态[J].植物生态学报,2012,31(2):282-291.
ZHANG X W,ZHANG X P,GUO C Y,etal.Quantitative dynamics ofPteroceltistatarinowiipopulations in Huangcangyu Nature Reserve of Anhui Province,East China[J].ChineseJournalofEcology,2012,31(2):282-291.
[36]杨斌,巨天珍,曹春,等.甘肃小陇山国家级自然保护区红豆杉种群动态分析[J].甘肃农业大学学报,2015,12(6):88-93.
YANG B,JU T Z,CAO C,etal.Dynamic analysis ofTaxuswallichianavar.chinensispopulation in Xiaolong Mountain National Nature Reserve,Gansu Province[J].JournalofGansuAgriculturalUniversity,2015,12(6):88-93.
[37]缪绅裕,曾庆昌,陈志明,等.南岭大东山长柄双花木群落物种组成与种群结构特征分析[J].植物资源与环境学报,2014,23(1):51-57.
MIAO S Y,ZENG Q C,CHEN Z M,etal.Analyses on community species composition and population structure characteristics ofDisanthuscercidifoliusvar.longipesin Dadongshan of Nanling Mountain[J].JournalofPlantResourcesandEnvironment,2014,23(1):51-57.
[38]黄雅昆,王得祥,张洪武,等.秦岭佛坪自然保护区短柄枹栎种群的数量特征[J].西北植物学报,2015,35(3):594-600.
HUANG Y K,WANG D X,ZHANG H W,etal.Quantitative characteristic ofQuercusglanduliferavar.brevipetiolatapopulation in Foping National Reserve of Qinling Mountains[J].Acta.Bot.Boreal.-Occident.Sin,2015,35(3):594-600.
[39]刘海洋,金晓玲,沈守云,等.湖南珍稀濒危植物--珙桐种群数量动态[J].生态学报,2012,32(24):7 738-7 746.
LIU H Y,JIN X L,SHEN S Y,etal.Population quantitative characteristics and dynamics of rare and endangered plantDavidiainvolucratain Hunan Province[J].ActaEcologicaSinica,2012,32(24):7 738-7 746.
[40]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
CHEN X D.A study on the method of quantitative analysis for plant population and community structural dynamics[J].ActaEcologicaSinica,1998,18(2):214-217.
[41]韩路,王家强,王海珍,等.塔里木河上游胡杨种群结构与动态[J].生态学报,2014,34(16):4 640-4 651.
HAN L,WANG J Q,WANG H Z,etal.The population structure and dynamics ofPopuluseuphraticaat the upper reaches of the Tarim River[J].ActaEcologicaSinica,2014,34(16):4 640-4 651.
[42]申仕康,马海英,王跃华,等.濒危植物猪血木(EuryodendronexcelsumH.T.Chang)自然种群结构及动态[J].生态学报,2008,28(5):2 404-2 412.
SHEN S K,MA H Y,WANG Y H,etal.The structure and dynamics of natural population of the endangered plantEuryodendronexcelsumH.T.Chang[J].ActaEcologicaSinica,2008,28(5):2 404-2 412.
[43]郭其强,卢杰,罗大庆,等.西藏色季拉山阳坡林线方枝柏种群结构特征研究[J].西北林学院学报,2010,25(2):15-18.
GUO Q Q,LU J,LUO D Q,etal.Ecological characteristics ofSabinasaltuariapopulation at the timber line ecotone on the sunny slope of the Sejila Mountain[J].JournalNorthwestForestryUniversity,2010,25(2):15-18.
[44]SILVERTOWN J W.Introduction to plant population ecology[M].London:Longman Group United Kingdom,1982.
[45]宋萍,洪伟,吴承祯,等.珍稀濒危植物桫椤种群结构与动态研究[J].应用生态学报,2005,16(3):413-418.
SONG P,HONG W,WU C Z,etal.Population structure and its dynamics of rare and endangered plantAlsophilaspinulosa[J].ChineseJournalofAppliedEcology,2005,16(3):413-418.
[46]杨小波.不同演替阶段森林群落优势种苗木叶形态解剖结构比较研究[J].海南大学学报(自然科学版),1997,15(3):212-217.
YANG X B.The comparative study on leaf anatomic characteristics of dominant species Seedling in different stages of forest secondary succession[J].NaturalScienceJournalofHainanUniversity,1997,15(3):212-217.
[47]杨小波,王伯荪.森林次生演替优势种苗木的光可塑性比较研究[J].植物学报,1999,16(3):113-118.
YANG X B,WANG B S.The comparative study on photo-plasticity of the dominant species seedlings of secondary forest succession[J].ChineseBulletinofBotany,1999,16(3):113-118.
[48]张军,黄永梅,焦会景,等.毛乌素沙地油蒿群落演替的生理生态学机制[J].中国沙漠,2007,27(6):977-983.
ZHANG J,HUANG Y M,JIAO H J,etal.Ecophysiological characteristics ofArtemisiaordosicacommunity succession in Mu Us Sandy Grassland[J].JournalofDesertResearch,2007,27(6):977-983.
[49]周文嵩,冯丹丹,李东海,等.海南昌江县濒危药用植物的种群动态[J].热带生物学报,2014,5(4):392-399.
ZHOU W S,FENG D D,LI D H,etal.Population dynamics of endangered medicinal plants in Changjiang County,Hainan[J].JournalofTropicalBiology,2014,5(4):392-399.
(编辑:潘新社)
Population Characteristics of Wild Litchi Resources in Wanning,Hainan Province
ZHANG Xuanrong1,2,LI Dan1,2,YANG Xiaobo1,2*,LI Donghai1,2,LIN Zeqin1,2,LI Yuanling1,2,LIU Zijin1,2
(1 College of Horticulture and Landscape Agriculture,Hainan University,Haikou 570228,China;2 Key Laboratory of Protection and Development Utilization of Tropical Crop Germplasm Resources,Haikou 570228,China)
Abstract:Wild litchi,the wild population of litchi,is an important gene bank for the betterment and innovation of litchi variety.The spatial distribution pattern,population diameter structure,static life table,and survival curve of litchi populations are studied base on the field survey of both inside and outside the natural conservation area.We revealed the population distribution and the trend of population dynamics of wild litchi,in order to provid the basis for the protection of wild litchi resources.The results show that:(1) wild litchi populations both inside and outside the natural reserve are aggregated distribution,and the aggregation degree of the non-natural conservation area is more obvious than that of the natural conservation area.(2) According to the age structure analysis,the population structure of wild litchi is growth-oriented,and its life expectancy is decremented with the age class increasing,and its survival curve is nearly linear type,which indicates that the population structure of wild litchi in the natural reserve is relatively stable;while those outside the natural reserve is of declining type.(3) The maximum of population dynamic index and random disturbance risky value of wild litchi population in the natural reserve is close to zero,but the value of the non-natural conservation area is higher that than that of the natural conservation area.It indicates that the sensitivity of the protection zone outside is stronger than inside.That is to say,the living environment of wild litchi outside the protection zone has been seriously disturbed and the threatened by human beings.The study suggests that,in the non-natural conservation area,the serious lack of saplings of wild litchi population is not only related to its own update mechanism,but more important is the destruction of the environment and human deforestation leading to structure change of wild litchi population and the decrease of wild litchi forest.
Key words:wild litchi;spatial distribution pattern analysis;population diameter structure;static life table;survival curve
中图分类号:Q948.12
文献标志码:A
作者简介:张萱蓉(1989-),女,硕士,从事植物生态学与植物资源学研究。E-mail:1160043639@qq.com*通信作者:杨小波,教授,博士生导师,从事植物生态学与植物资源学研究。E-mail:yanfengxb@163.com
基金项目:国家自然科学基金(31460120)
收稿日期:2015-09-09;修改稿收到日期:2016-03-08
文章编号:1000-4025(2016)03-0596-10
doi:10.7606/j.issn.1000-4025.2016.03.0596
