8个大粒裸燕麦品种核型研究
2016-04-27耿帆周青平梁国玲贾志锋刘文辉丁成翔刘勇颜红波
耿帆,周青平,梁国玲,贾志锋,刘文辉,丁成翔,刘勇,颜红波
(1.青海大学农牧学院,青海 西宁 810016;2.青海省畜牧兽医科学院,青海 西宁 810016;3.西南民族大学青藏高原研究所,
四川 成都 610041;4.青藏高原优良牧草种质资源研究利用省级重点实验室,青海 西宁 810016)
8个大粒裸燕麦品种核型研究
耿帆1,2,4,周青平3*,梁国玲2,4,贾志锋2,4,刘文辉2,4,丁成翔2,4,刘勇2,4,颜红波2,4
(1.青海大学农牧学院,青海 西宁 810016;2.青海省畜牧兽医科学院,青海 西宁 810016;3.西南民族大学青藏高原研究所,
四川 成都 610041;4.青藏高原优良牧草种质资源研究利用省级重点实验室,青海 西宁 810016)
摘要:采用常规压片法对来源于我国不同省份的8个大粒裸燕麦品种进行了核型分析及进化趋势研究,旨在为燕麦种质资源的收集、鉴定、开发利用和良种培育提供细胞学依据。研究结果表明,8份大粒裸燕麦材料的核型公式分别为:青莜3号:2n=6x=42=22m+20sm (4SAT);平安莜麦:2n=6x=42=18m+24sm (4SAT);白燕2号:2n=6x=42=18m+24sm (2SAT);花早2号:2n=6x=42=20m+22sm (2SAT);花晚6号:2n=6x=42=14m+28sm (6SAT);晋燕8号:2n=6x=42=22m+20sm (4SAT);宁莜1号:2n=6x=42=16m+26sm (2SAT);定莜6号:2n=6x=42=16m+26sm (4SAT),8个燕麦品种的核型均属于2B型。进行聚类及进化趋势分析发现,白燕2号为最进化的品种,青莜3号为最原始的品种。
关键词:核型分析;裸燕麦;聚类分析
燕麦隶属于禾本科(Gramineae)燕麦属(Avena)1年生草本植物,一般分为裸燕麦(Avenanuda)和皮燕麦(Avenasterieis)两种,世界各国以种植皮燕麦为主,而我国是大粒裸燕麦的起源中心和主产国[1-2]。大粒裸燕麦以其较高的营养价值、医疗保健价值以及特有的加工优势受到了广泛欢迎[3-7]。目前我国主产大粒裸燕麦的地区主要有内蒙古、河北、吉林、山西、宁夏和甘肃等省份,由于各地区间气候条件等的差异,各品种皆形成了各自的特性,丰富了我国大粒裸燕麦种质资源的遗传多样性[8]。目前有关大粒裸燕麦的研究只对内蒙古和山西等少部分地区的种质进行了核型分析[9-11],有关其他地区及不同地区间种质核型的系统研究鲜有报道。本研究以来自于我国不同省份的8个大粒裸燕麦品种作为实验材料,对其核型及进化趋势进行研究,通过聚类分析了解其间的亲缘关系,旨在为我国燕麦种质资源的收集、开发利用和良种培育提供必要的细胞学依据。
1材料与方法
1.1材料
供试材料选择来自青海、吉林、河北、山西、宁夏和甘肃的8份燕麦材料,供试材料详情见表1。
1.2方法
该实验于2014年3月-2015年1月在青海省青藏高原优良牧草种质资源利用重点实验室进行。
1.2.1染色体制片方法每份材料各选取30粒饱满的种子,放入干净垫有多层浸水滤纸的培养皿中,置于23℃恒温培养箱中培养至露白,将露白种子放在洁净的垫有已灭菌浸水滤纸的培养皿中,置于4℃冰箱内培养48 h,然后置于23℃恒温培养箱中培养,待根长至1.0~2.0 cm时, 剪取新生粗壮根尖于冰水中预处理48 h,接着将根尖转入卡诺氏固定液(无水乙醇∶冰乙酸=3∶1,V/V)中在4℃下固定24 h,然后用1 mol/L盐酸在60℃下解离12~13 min,最后用改良的苯酚品红染色15 min后常规压片。制好的临时装片在UB100i series光学显微镜下观察并拍照。

表1 实验材料
1.2.2染色体核型分析方法分别选取30个分裂中期染色体分散良好的细胞进行染色体计数,将大多数(85%以上)细胞共有的染色体数目记作所研究燕麦材料的标准染色体数目。依据李懋学和陈瑞阳[12]的植物核型分析标准,每个品种分别选取5个染色体分散良好、形态清晰完整的高质量细胞中期分裂相照片用于核型分析。参照杨大翔[13]及乔永刚和宋芸[14]所述方法,利用Photoshop和Excel软件对染色体照片进行处理,完成核型图及核型模式图的制作。染色体分类按照Levan[15]的命名方法进行,核型分类按照Stebbins[16]的分类系统进行。
1.3数据分析
参照周娅[17]的方法绘制核型不对称性程度散点图。以染色体长度比为纵坐标、平均臂比为横坐标制图,坐标点的分布即可表示各材料核型不对称性及进化的情况,坐标点越靠近右上方说明其核型不对称性较大和进化程度较高,越靠近左下方说明其核型不对称性较小和进化程度较原始。参照戴小红[18]的方法对核型参数(核型不对称系数、平均臂比和染色体长度比)进行系统聚类分析。利用SPSS 19.0软件,对原始数据作标准变换,然后计算欧氏距离,应用最远邻元素法进行聚类分析。
2结果与分析
2.1核型特征
显微镜下观察结果显示,8份大粒裸燕麦材料均为六倍体,体细胞染色体数目均为2n=6x=42,染色体照片、核型图及核型模式图见图1,主要核型参数见表2。

图1 供试大粒裸燕麦材料的染色体照片、核型图及核型模式图Fig.1 Chromosomes shape, karyotype and karyotype pattern of the selected oat cultivars
由表2可知,8份大粒裸燕麦材料的染色体均由m和sm两种类型组成,但在组成数目上存在差异,宁莜1号和定莜6号、平安莜麦和白燕2号、青莜3号和晋燕8号两两之间2种类型染色体的组成数目相同,其他染色体之间均不相同。由图1易见不同材料间随体的数目和位置也存在差异,其中白燕2号、花早2号和宁莜2号有1对随体染色体;花晚6号有3对随体染色体;其余4份材料均有2对随体染色体。8份大粒裸燕麦材料的染色体长度比、平均臂比及核不对称系数差异不大,变幅分别为2.02~2.72、1.75~1.92及63.06%~64.97%,核型均属于2B型,是Stebbins核型分类标准中较为进化的核型。
2.2核型参数的散点图分析
平均臂比和染色体长度比可以表示不同种源间核型的不对称性,值越大,其核型越不对称。由8份大粒裸燕麦材料的核型不对称性程度散点图(图2)可知,白燕2号在平均臂比方向进化趋快;青莜3号和宁莜1号在平均臂比方向和染色体长度比方向进化趋一致;花早2号、花晚6号和定莜6号在染色体长度比方向进化趋快。

表2 供试大粒裸燕麦材料的核型参数
注:n表示物种配子体的染色体个数;x表示一个染色体组中染色体数目;m表示中部着丝点区染色体;sm表示近中部着丝点区染色体;SAT表示随体染色体。
Note:nwas the chromosome number of species’ gametophyte;xwas the chromosome number of a genome; m was median region chromosome; sm was submedian region chromosome; SAT was satellite chromosome.
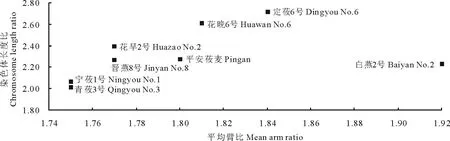
图2 供试材料的核型不对称性程度散点图Fig.2 Scatter diagram of the selected cultivars based on the degree of karyotype asymmetry
2.38份大粒裸燕麦材料的聚类分析
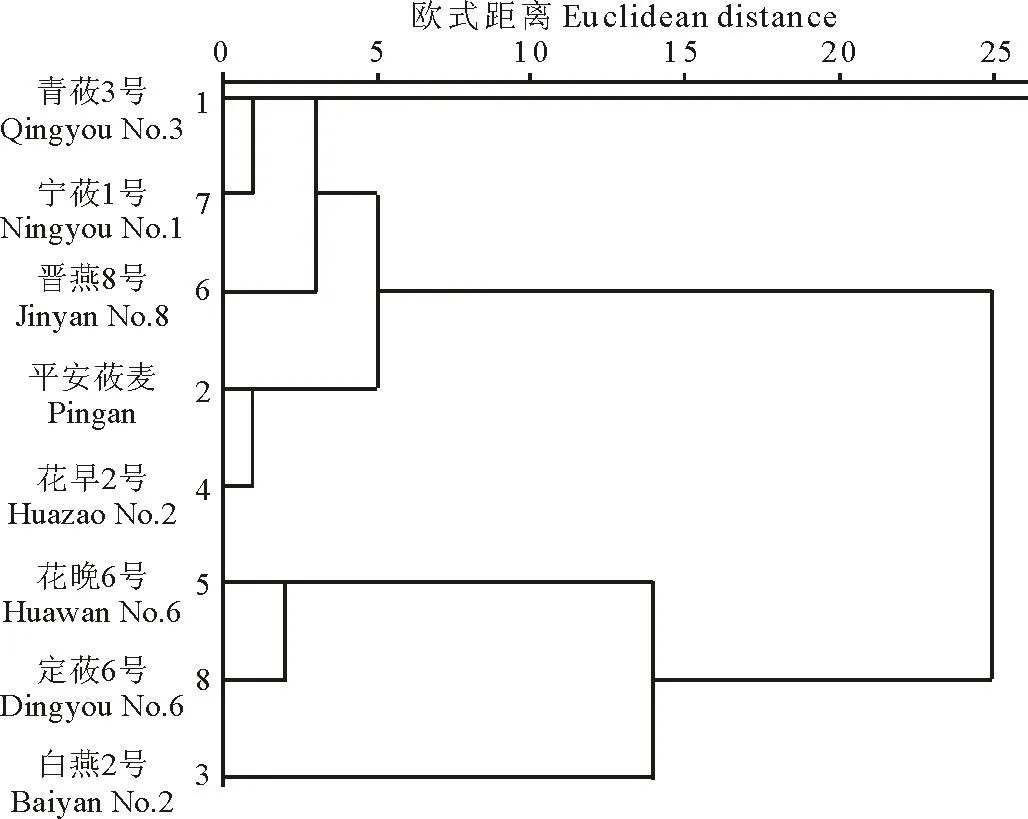
图3 8份大粒裸燕麦材料的聚类分析图Fig.3 Dendrogram of cluster analysis of eight oat cultivars
8份大粒裸燕麦材料核型参数的聚类结果见图3,当欧式距离为10时可将8份材料分为3类:第Ⅲ类只有白燕2号;第Ⅱ类包括花晚6号和定莜6号;其余5份材料归为第Ⅰ类。划分为同一类的材料染色体特征相似性大。由各分类的核型参数均值(表3)可知,第Ⅲ类的核不对称系数和平均臂比最大,分别为64.97%和1.92;第Ⅱ类的染色体长度比最大,为2.67;第Ⅰ类的各指标值居中。
3讨论
3.1供试材料的核型特征
本试验中选取的8份大粒裸燕麦材料的染色体类型与核型类型与刘充治等[9]、武生辉等[10-11]及刘伟[19]的研究结果一致,均为m、sm 2种染色体类型和2B核型类型;但武生辉等[20]对内蒙古农家品种五寨莜麦(A.nudacv. wuzhai)进行了核型分析,结果表明五寨莜麦的染色体类型除了有m和sm 2种类型之外,还有3对st(近端部着丝点区染色体)型,这与本研究的结果差异较大,可能原因是材料的来源存在较大差异。本试验的结果表明,所选8份大粒裸燕麦材料间的m和sm 2种类型染色体的组成数目及随体染色体数目均存在差异,前人的研究也表现出了这种差异。 武生辉等[10-11]得出蒙燕7312莜麦和内燕5号莜麦的染色体组成为22m+20sm,均具有1对随体染色体;刘伟[19]对国外引进的大粒裸燕麦的研究结果表明国外大粒裸燕麦的染色体组成为24m+18sm,具有1对随体染色体;刘充治等[9]的研究结果表明加拿大大粒裸燕麦Abvgtou、山西大粒裸燕麦三分三及玉麦L13的染色体组成为10m+11sm,均具有3对随体染色体,内蒙古的左37裸燕麦的染色体组成为12m+9sm,具有3对随体染色体,燕麦1079的染色体组成为13m+8sm,具有2对随体染色体。这些差异表明我国的大粒裸燕麦种质资源丰富,在以后的研究当中应该加强对我国各地区大粒裸燕麦种质资源的综合研究。

表3 8份大粒裸燕麦材料各分类的核型参数均值
3.28份大粒裸燕麦材料的进化分析
Stebbins[16]认为核型愈不对称一般表明愈进化,核型不对称性的增加是指染色体两臂长度不等,或是同一核内染色体大小不等,两者可分别用平均臂比和染色体长度比表述。王米力和石大兴[21]在分析云杉属内种质的核型进化以及邢世岩等[22]对银杏特异种质核型进化趋势的研究中,发现所研究对象的核型呈“双向进化趋势”,即一些种沿染色体长度比方向进化快,一些种沿平均臂比方向进化快,本研究在大粒裸燕麦中也发现了类似的双向进化趋势。
Arano[23]研究发现,核型不对称系数越大,染色体不对称性越大,其进化程度越高。刘慧民等[24]曾报道染色体核型参数的重要性排序为:核型不对称系数>平均臂比>臂比大于2的比率>染色体长度比>核型类型。通过染色体相关核型参数的分析得知8份大粒裸燕麦材料核型不对称程度依次为:白燕2号>定莜6号>花晚6号>花早2号>晋燕8号>平安莜麦>宁莜1号>青莜3号。其中白燕2号在8份大粒裸燕麦材料中核型不对称程度最高,相对进化程度愈大,其次是定莜6号;青莜3号的核不对称程度最低,相对较原始;青海省地方品种平安莜麦的核不对称程度位于中间位置。
3.38份大粒裸燕麦材料间的亲缘关系的比较
在聚类分析过程中,被聚在一起的往往具有较近的亲缘关系。当欧式距离为10时,8份大粒裸燕麦材料被分成了3大类,表明这3大类材料之间亲缘关系相对较远。本试验所选的8份大粒裸燕麦材料皆是有其各自特性的优良品种,如花晚6号具有很好的粮草兼用性,晋燕8号具有较高含量的蛋白质、脂肪和亚油酸,主要用于燕麦食品的加工。在育种中,可选择具有良好目标品质且亲缘关系较远的材料进行杂交,以期培育出综合生产性能较好的燕麦新品种。
References:
[1]Yang H P, Sun Z M. China Oat[M]. Beijing: Beijing Agricultural Press,1989.
[2]Wang M Y, Qi X L, Zhang F Y. Progress in the research of oat molecular biology abroad. Journal of Inner Mongolia Institute of Agricultural and Animal Husbandry, 2001, 22(4): 139-144.
[3]Dong Y C, Liu X. Crops and Their Wild Relatives in China: Food Crops to Volume[M]. Beijing: China Agriculture Press, 2006.
[4]Liu H. Health care function and its development ofAvenanudaL. Journal of Cereals & Oils, 1999, (2): 11-12.
[5]Li T, Zuo F L, Li Z G,etal. Present situation and development prospect ofAvenaL. Farm Machinery, 2012, (24): 88-92.
[6]Beer M U, Arrigoni E, Amado R. Effect of oat gum on blood cholesterol levels in healthy young men. European Journal of Clinical Nutrition, 1995, 49(7): 517-522.
[7]Peterson D M. Oat antioxidants. Journal of Eal Science, 2001, 33: 115-129.
[8]Zhao S F, Tian C Y, Wang Z G,etal. Oat production and research present situation and future development direction in China. Rain Fed Crops, 2007, 27(6): 428-431.
[9]Liu C Z, Pei C E, Li G Q,etal. Karyotype analysis of hexaploid oat. Journal of Shanxi Agricultural University, 1986, 6(2): 169-171.
[10]Wu S H, Li X X, Zhao X Y,etal. A comparison of karyotypes between naked oat and oat bran. Journal of Inner Mongolia Academy of Agricultural and Animal Husbandry, 1997, 18(4): 12-15.
[11]Wu S H, Li X X, Li M Z. Study on the karyotype ofAvenanuda. Journal of Inner Mongolia Academy of Agricultural and Animal Husbandry, 1988, 9(2): 23-27.
[12]Li M X, Chen R Y. Standardization of plant nuclear analysis. Botanical Research of Wuhan, 1985, 3(4): 297-302.
[13]Yang D X. Karyotype analysis by Adobe Photoshop. Agricultural Network Information, 2005, (3): 45-46.
[14]Qiao Y G, Song Y. Making use of EXCLE to produce the karyotype patterns. Agricultural Network Information, 2006, (10): 97-98.
[15]Levan A. Nomenclature for centromere position in chromosome. Hereditas, 1964, 52: 201-220.
[16]Stebbins G L. Chromosome Evolution in High Plants[M]. London: Edward Arnold, 1971: 87-90.
[17]Zhou Y. Study on the Karyotype of Nine Species of Pinus[D]. Yaan: Agricultural University of Sichuan, 2012.
[18]Dai X H. Analysis on the Genetic Diversity and Phylogenetic Relationships of the Genus Plants in the Genus of the Qin Ba Mountain Area[D]. Yangling: Northwest Agriculture and Forestry University, 2006.
[19]Liu W. Chromosome Identification and SSR Fingerprinting of Different Ploidy Oat Germplasm[D]. Beijing: Chinese Academy of Agricultural Sciences, 2012.
[20]Wu S H, Li X X, Li M Z,etal. Study on the karyotype of Wuzhai naked oats. Journal of Inner Mongolia Academy of Agricultural and Animal Husbandry, 1996, 17(1): 41-44.
[21]Wang M L, Shi D X. Karyotype analysis of six species of spruce. Plant Research of Yunnan, 1994, 16(2): 157-164.
[22]Xing S Y, Gao J H, Jiang Y Z,etal. The evolution trend of specific germplasm ofGinkgobiloba. Forestry Science, 2007, 43(1): 21-27.
[23]Arano H. Cytological studies in subfamily Carduoideae (Compositae) of Japan VIII. The karyotype analysis in tribe Senecioneae. The Botanical Magazine, 1963, 76: 32-39.
[24]Liu H M, Chen Y J, Lv G E,etal. Karyotype characteristics and parameters analysis of 17 kinds of embroidery thread. Journal of Horticulture, 2010, 37(9): 1456-1462.
参考文献:
[1]杨海鹏, 孙泽民. 中国燕麦[M]. 北京: 农业出版社, 1989.
[2]王茅雁, 齐秀丽, 张凤英. 国外燕麦分子生物学研究进展. 内蒙古农业大学学报, 2001, 22(4): 139-144.
[3]董玉琛, 刘旭. 中国作物及其野生近缘植物: 粮食作物卷[M]. 北京: 中国农业出版社, 2006.
[4]刘河. 裸燕麦保健功能及其开发. 粮食与油脂, 1999, (2): 11-12.
[5]李婷, 左芳雷, 李再贵, 等. 燕麦的研究现状及其发展前景. 农业机械, 2012, (24): 88-92.
[8]赵世锋, 田长叶, 王志刚, 等.我国燕麦生产和科研现状及未来发展方向. 杂粮作物, 2007, 27(6): 428-431.
[9]刘充治, 裴成娥, 李贵全, 等. 六倍体裸燕麦的核型研究. 山西农业大学学报, 1986, 6(2): 169-171.
[10]武生辉, 李秀娴, 赵晓英, 等. 裸燕麦和皮燕麦的核型比较. 内蒙古农牧学院学报, 1997, 18(4): 12-15.
[11]武生辉, 李秀娴, 李明哲. 裸燕麦(Avenanuda)的核型研究. 内蒙古农牧学院学报, 1988, 9(2): 23-27.
[12]李懋学, 陈瑞阳. 关于植物核型分析的标准化问题. 武汉植物学研究, 1985, 3(4): 297-302.
[13]杨大翔. 用Adobe Photoshop进行核型分析. 农业网络信息, 2005, (3): 45-46.
[14]乔永刚, 宋芸. 利用EXCLE制作核型模式图. 农业网络信息, 2006, (10): 97-98.
[17]周娅. 九种松属植物的核型研究[D]. 雅安: 四川农业大学, 2012.
[18]戴小红. 秦巴山区百合属植物的核型多样性与亲缘关系分析[D]. 杨凌: 西北农林科技大学, 2006.
[19]刘伟. 不同倍性燕麦种质核型鉴定和SSR指纹图谱构建[D]. 北京: 中国农业科学院, 2012.
[20]武生辉, 李秀娴, 李明哲, 等. 五寨筱麦的核型研究. 内蒙古农牧学院学报, 1996, 17(1): 41-44.
[21]王米力, 石大兴. 六种云杉的核型分析. 云南植物研究, 1994, 16(2): 157-164.
[22]邢世岩, 髙进红, 姜岳忠, 等. 银杏特异种质核型进化趋势. 林业科学, 2007, 43(1): 21-27.
[24]刘慧民, 陈雅君, 吕贵娥, 等. 17种绣线菊核型特征及核型参数分析. 园艺学报, 2010, 37(9): 1456-1462.
Karyotype study of 8 kinds ofAvenanuda
GENG Fan1,2,4, ZHOU Qing-Ping3*, LIANG Guo-Ling2,4, JIA Zhi-Feng2,4, LIU Wen-Hui2,4, DING Cheng-Xiang2,4, LIU Yong2,4, YAN Hong-Bo2,4
1.CollegeofAgricultureandAnimalHusbandry,QinghaiUniversity,Xining810016,China; 2.AcademyofAnimalScienceandVeterinaryMedicineofQinghaiProvince,Xining810016,China; 3.CollegeofQing-TibetanPlateauofSouthwestUniversityforNationalities,Chengdu610041,China; 4.KeyLaboratoryofSuperiorForageGermplasmintheQinghai-Tibetan,Xining810016,China
Abstract:In order to provide cytological information for the collection, identification, utilization and breeding of Avena nuda germplasm, karyotype analysis and an evolution trend investigation have been undertaken of 8 A. nuda cultivars from different provinces of China using the conventional tableting method. The karyotype formulas of the 8 cultivars as follows: Qingyou No. 3: 2n=6x=42=22m+20sm (4SAT); Pingan: 2n=6x=42=18m+24sm (4SAT); Baiyan No. 2: 2n=6x=42=18m+24sm (2SAT); Huazao No. 2: 2n=6x=42=20m+22sm (2SAT); Huawan No. 6: 2n=6x=42=14m+28sm (6SAT); Jinyan No. 8: 2n=6x=42=22m+20sm (4SAT); Ningyou No. 1: 2n=6x=42=16m+26sm (2SAT); Dingyou No. 6: 2n=6x=42=16m+26sm (4SAT). The karyotype of the 8 cultivars belongs to the 2B type. According to clustering and evolutionary trend analysis, Baiyan No. 2 is a relatively new cultivar and Qingyou No. 3 is the most original cultivar.
Key words:karyotype analysis; Avena nuda; cluster analysis
*通信作者
Corresponding author. E-mail:qpingzh@aliyun.com
作者简介:耿帆(1988-),女,陕西泾阳人,在读硕士。E-mail:892492927@qq.com
基金项目:国家科技部农转资金项目“莜麦新品种青引3号试验与示范(2013GB2G200503)”,青海省农牧科技领域人才“小高地”,国家科技部科技支撑项目“青海地区优质牧草选育及生产利用技术集成与示范(2011BAD17B05-5)”,“现代农业产业技术体系建设专项资金资助”(CARS-35-41)和青藏高原牧草种质资源保护利用(070401)资助。
收稿日期:2015-02-25;改回日期:2015-11-03
DOI:10.11686/cyxb2015101
http://cyxb.lzu.edu.cn
耿帆,周青平,梁国玲,贾志锋,刘文辉,丁成翔,刘勇,颜红波. 8个大粒裸燕麦品种核型研究.草业学报, 2016, 25(3): 120-125.
GENG Fan, ZHOU Qing-Ping, LIANG Guo-Ling, JIA Zhi-Feng, LIU Wen-Hui, DING Cheng-Xiang, LIU Yong, YAN Hong-Bo. Karyotype study of 8 kinds ofAvenanuda. Acta Prataculturae Sinica, 2016, 25(3): 120-125.
