18种螯虾COⅠ及12S rRNA基因序列差异及其系统进化关系
2016-04-23马景微曹谨玲刘少贞
宋 晶,马景微,曹谨玲,刘少贞
(山西农业大学动物科技学院,山西太谷 030801)
螯虾属于甲壳纲、十足目、螯虾总科,螯虾科(Cambaridae)现分为12个属,分布于除美国、加拿大西部太平洋沿岸地区的整个北美洲大陆,日本以及亚洲东部的国家地区。我国分布2属4种,其中东北螯虾(Cambaroides dauricus)、朝鲜螯虾(Cambaroides similis) 和许郎螯虾(Cambaroides schrenkii)为中国的本地品种,克氏原螯虾(Procambarus clarkii)原产于北美,20世纪30年代由日本引进中国并逐渐成为优势种,是中国重要的水产资源[1-2]。中国淡水螯虾产业近20年来的进出口总量不断增加,许多淡水螯虾品类经济价值较高,是重要的水产经济动物。除了部分供给水产品市场,很多体型、体色特异的种类也当作宠物进行养殖[3]。
螯虾体征相似,种内体色差异大,依赖形态进行分类有一定的局限性,因其表型和遗传的数据有着明显差异,其中一个明显的弊端就是很难排除可能受到趋同进化的干扰[4]。在物种的进化历程中,DNA分子是遗传信息的载体,也是动物进化历史的忠实记录者。DNA分子一级序列中的所有碱基都具有较为独立的特征,极少会受到趋同适应的干扰。因此,可将DNA分子提供的一级序列的信息作为螯虾科系统分类的依据。其中,线粒体基因的结构简单、母系遗传、无内含子、进化速度快且不发生重组,作为一种分子标记技术被广泛应用[5]。
随着分子系统学的发展,使人们更加热衷于从分子水平上研究物种的系统发生关系,而系统发育树可用类似树状分支的图直接展示各种生物之间可能的亲缘关系。主要是通过DNA序列、蛋白质序列、蛋白质结构等来构建系统发育树,蛋白质结构比较利用刚体结构叠合和多结构特征比较等方法建立结构进化树。近十年来,国外已有学者相继利用分子手段研究了巨石螯虾属的系统发育和谱系地理[6]、螯虾属穴居种类的进化史[7]、侏儒螯虾亚科的进化和系统学[8],2012年,Breinholt还提出了淡水螯虾科各属的进化假说[9],但螯虾科各属的亲缘关系仍尚无定论。本研究旨在应用分子生物学技术对螯虾科进行分类及进化关系的研究,使得螯虾科在分子层面上的分类程度更加成熟,从而能够为螯虾系统分类研究提供一个有效利用资源的途径。
1 材料和方法
1.1 试验材料
从 NCBI(http://www.ncbi.nlm.nih.gov/)下载获得隶属于螯虾科的螯虾属、拟螯虾属、叉肢螯虾属、原螯虾属、侏儒螯虾属和Fallicambarus属中的18种螯虾(表1)的线粒体序列提取12S rRNA和COⅠ基因,选用亲缘关系适中的普通滨蟹(C.maenas)为外群。
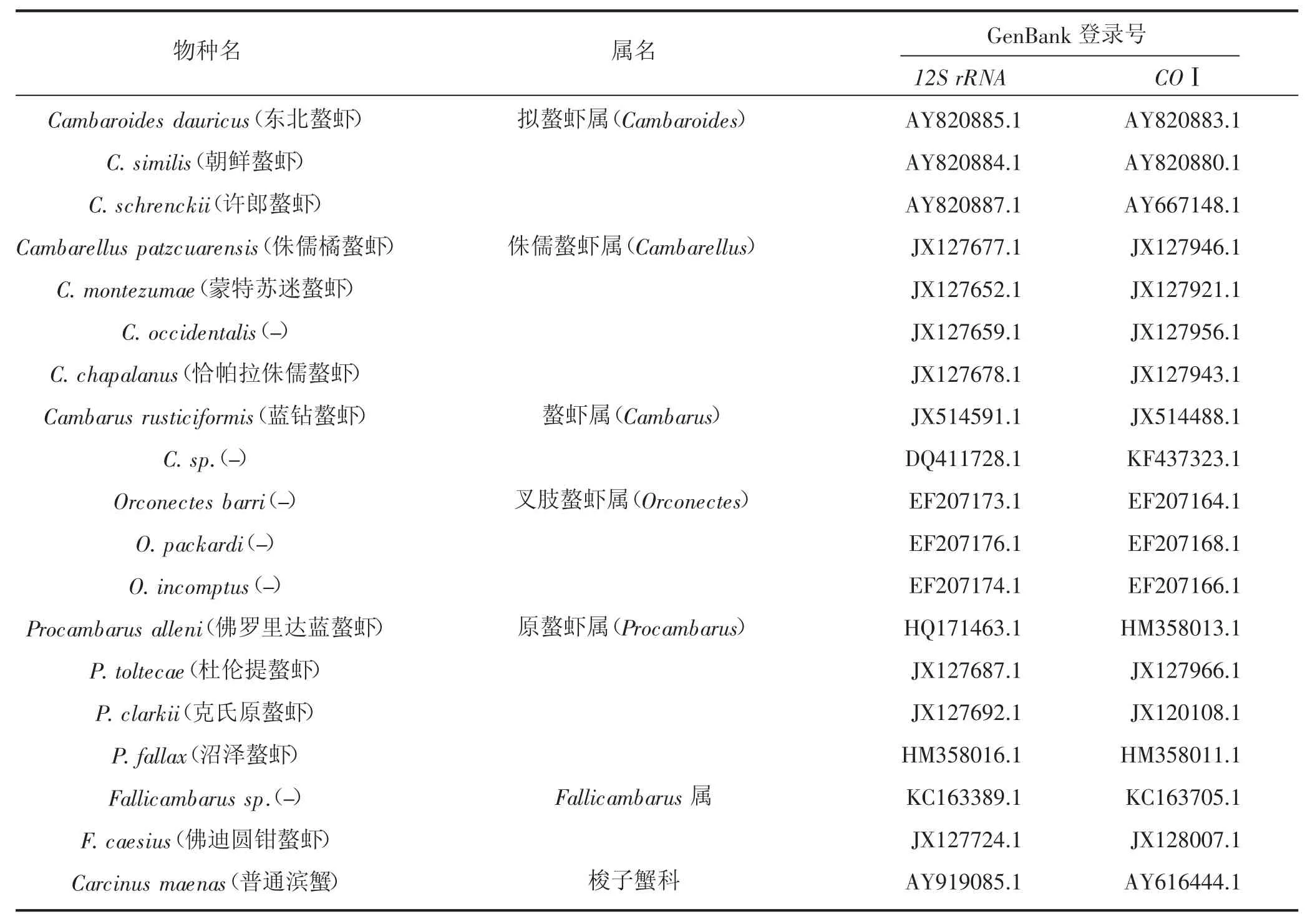
表1 物种名称和GenBank登录号
1.2 试验方法
将所有序列用NCBI网站提供的Blast分析工具包进行相似性检索,确认实验所得序列为目的片段。所有具备已知序列的物种都进行碱基序列变异统计,然后存入文本文档中,修改为“.fasta”格式,进行比对、剪切之后将前面的文件转化格式为“.meg”格式,再进行激活,运用软件MEGA 6.0计算序列的种间遗传距离,选择普通滨蟹(C.maenas)作为外群,最后构建进化树。
多种软件都可用于分析分子数据来重建系统发育树,然而没有单一的方法对所有情况都能取得很好的效果。比选择软件更重要的是生物信息数据的质量,软件不具备对输入数据进行更正的功能,所以采用不同进化速率的基因和多种构树方法来实现系统分析是较为合适的选择。本研究采用了邻接法(Neighbour-Joining,NJ) 和最大似然法(Maximum Likelihood,ML)来构建螯虾科部分属、种的系统发育树。
2 结果与分析
2.1 碱基序列变异比
从下载的线粒体全序列中选取12S rRNA、COⅠ线粒体序列,基于Kimura双参数模型,计算螯虾科种间遗传距离P值。基于12S rRNA序列的螯虾科种间遗传距离为0.002~0.127,平均P值为0.048;基于COⅠ基因的螯虾科种间遗传距离为0.031~0.351,平均遗传距离为0.161。由此可知,12S rRNA基因的进化速度远大于COⅠ基因,基因进化速率的差异很可能是导致构建的系统进化树产生不同拓扑结构的原因[10]。
2.2 分子系统进化
邻接法是基于距离的建树方法,其最大优势是运算十分简便快捷,然而对各位点的具体情况不进行考虑,因此容易丢失一些遗传信息。最大似然法是由样本估测值估计总体参数的方法,该方法在每组序列比对中考虑了每个核苷酸替换的概率,因此计算复杂,耗时较长[11]。以普通滨蟹为外群,用Bootstrap法检验,1 000次重复抽样得到结点的置信度来构建系统发生树(图1、图2)。
共用ML法和NJ法2种方法建立的系统进化树,得到的结果具有相似的拓扑结构,并且都具有较高的节点支持率。本研究的2种系统进化树中关于螯虾科的拓扑型与形态学分类结构基本一致,表明螯虾科的12S rRNA及COⅠ基因序列可用于属间的系统发生关系分析,构建的进化树结果可靠。但是在个别分区有与经典的形态分类不相符的地方,如未定名种Cambarus sp.的归属及有效性都有争议,系统发育分析的结果表示,其很可能不属于螯虾属,而与拟螯虾属有着更高的遗传相似度。而蓝钻螯虾(C.rusticiformis)也显示出与叉肢螯虾属有较近的亲缘关系。
3 讨论
形态分类的参数较多且稳定性不足,因此趋同进化有时会使传统的形态分类变得混乱。对于螯虾属(Cambarus)的种类,多以螯虾的特征作为属下的分类依据,而同科的叉肢螯虾属(Orconectes),则多以生殖肢的形态作为分类特征。在利用生殖肢形态对原螯虾属(Procambarus)和叉肢螯虾属(Orconectes)进行亚属划分时,也都受到趋同进化的影响[9]。因此,可以利用生物信息数据构建系统发育树,辅助传统的经典分类。不同的基因进化速率不同,利用单一基因构造系统发育树,很难准确地反映种属之间的亲缘关系。
分析螯虾科18个物种的12S rRNA基因的系统发育树。侏儒螯虾属、原螯虾属、叉肢螯虾属与Fallicambarus属位于同一分支(节点支持率为ML树88、NJ树95),其中克氏原螯虾先与侏儒螯虾属聚为一支,然后再与原螯虾属聚为一支,拟螯虾属与其平行进化,NJ树与ML树的节点支持率分别高达97和99。除了COⅠ基因构建的ML树,克氏原螯虾分类归属于原螯虾属,而对其他树都有不同程度的扰动,其原因可能是由于外群选择会影响基于距离的构树方法和12S rRNA序列较短(仅257 bp)所致[11]。分析得到的螯虾科18种类的COⅠ基因的系统发育树。观察得到这三种树的拓扑图总体趋势相似,其中侏儒螯虾属与原螯虾属亲缘关系较近,而螯虾属多数种类自成一支,Fallicambarus属较为特殊,总能独立分出(节点支持率在82~97),进行平行进化。
根据数据分析得到结果与形态学分类比较一致,这进一步证明了线粒体12S rRNA及COⅠ基因序列可作为推断种属间关系的有效遗传标记,而相对序列较长、进化速率也较快,COⅠ基因的分析结果与螯虾科在属这一级别上形态分类的契合度更高。在其他近缘种上也有相似的结论,如对于铠甲虾类的系统发育研究也曾采用16S rRNA及COⅠ基因进行研究,并证实COⅠ具有更好的指示效果[12],本研究对螯虾科部分种类的12S rRNA、COⅠ基因序列进行分析,探讨了该科种类的系统发育关系,为相关研究提供了基础实验数据和分子证据。但因所用物种非常有限,分析结果不一定全面反映该科的系统发育关系,有待于下一步进行更广泛和更深入的研究。
参考文献:
[1]Crandall K A,Buhay J E.Global diversity of crayfish(Astac idae,Cambaridae,and Parastacidae-Decapoda)in freshw ater[J].Hydrobiologia,2008,595(1):295-301.
[2]慕 峰,成永旭,吴旭干.世界淡水螯虾的分布与产业发展[J].上海水产大学学报,2007,16(1):64-71.
[3]Faulkes Z.Marmorkrebs (Procambarus fallax f.virginalis)are the most popular crayfish in the North American pet trade[J].Knowl Manag Aquat Ecosyst,2015,416:20-35.
[4]Brown W M,George J R,Wilson A C.Rapid evolution of animal mitchondrial DNA[J].P Natl Acad Sci USA,1979,76(4):1967-1971.
[5]Avise J C,Arnold J,Ball R N M,et al.Intraspecific phylogeography:the mitochondrial bridge between popula tion genetics and systematic[J].Annu Rev Ecol Syst,1987,18:489-522.
[6]Buhay J E,Moni G,Mann N,et al.Molecular taxonomy in the dark:evolutionary history,phylogeography,and diversity of cave crayfish in the subgenus Aviticambarus,genus Ca mbarus[J].Mol Phylogenet Evol,2007,42(2):435-448.
[7]Pedraza-Lara C,Doadrio I,Breinholt J W.Phylogeny and evolutionary patterns in the dwarf crayfish subfamily(Decapoda:Cambarellinae)[J].Plos One,2012,7(11):e48233.
[8]Trontelj P,Machino Y,Sket B.Phylogenetic and phylogeo graphic relationships in the crayfish genus Austropotamob ius inferred from mitochondrial COI gene sequences[J].Mol Phylogenet Evol,2005,34(1):212-226.
[9]Breinholt J W,Porter M L,Crandall K A.Testing phylogen etic hypotheses of the subgenera of the freshwater crayfish genus Cambarus(Decapoda:Cambaridae)[J].Plos One,2012,7(9):e46105.
[10]王利利,彭建军,于冬梅,等.Cytb和12S rRNA基因进化速度对构建系统进化树的影响[J].安徽农业科学,2011,39(9):5101-5103.
[11]张丽娜,荣昌鹤,何远,等.常用系统发育树构建算法和软件鸟瞰[J].动物学研究,2013,34(6):640-650.
[12]刘昕明.深海热液区几种铠甲虾形态学分类及分子系统发育研究[D].厦门:国家海洋局第三海洋研究所,2013:54-55.
