干旱胁迫对黑果枸杞幼苗光合特性的影响
2016-03-23郭有燕刘宏军孔东升张亚娟刘东花
郭有燕,刘宏军,孔东升*,闫 芳,张亚娟,刘东花
(1 河西学院,甘肃张掖 734000;2 张掖市种子管理站,甘肃张掖 734000)
干旱胁迫对黑果枸杞幼苗光合特性的影响
郭有燕1,刘宏军2,孔东升1*,闫芳1,张亚娟1,刘东花1
(1 河西学院,甘肃张掖 734000;2 张掖市种子管理站,甘肃张掖 734000)
摘要:以当年生黑果枸杞幼苗为试验材料,通过称重控水的方法设置对照(土壤含水量为32.96%~35.35%)、轻度干旱胁迫(土壤含水量为21.18%~22.32%)、中度干旱胁迫(土壤含水量为12.20%~13.82%)和重度干旱胁迫(土壤含水量为7.89%~8.73%)4个水分梯度,研究了干旱胁迫对黑果枸杞叶片光合色素、光合特性、叶绿素荧光特性的影响,以揭示黑果枸杞对干旱胁迫的适应能力和适应机制。结果显示:(1)随着干旱胁迫强度的增加,黑果枸杞幼苗叶片叶绿素含量、类胡萝卜素含量均呈显著下降趋势。(2)黑果枸杞幼苗叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)在中度和重度干旱胁迫下显著下降;其胞间CO2浓度(Ci)、水分利用效率(WUE)随干旱胁迫强度的增加而逐渐增加,而气孔限制值(Ls)随干旱胁迫强度的增加而逐渐降低。(3)随着土壤含水量的降低,黑果枸杞幼苗叶片初始荧光(F0)和非光化学猝灭系数(qspan)逐渐增加,而其最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)、实际光化学效率(ФPSⅡ)和光化学猝灭系数(qspan)均逐渐降低。研究表明,在干旱胁迫条件下,黑果枸杞叶片过多的能量以热的形式被耗散,反应中心开放程度降低,从而避免PSⅡ 反应中心受到损伤,表现出一定的耐旱性;黑果枸杞生长所允许的最大土壤水分亏缺为7.89%,维持黑果枸杞具有较高的WUE和Pn的土壤水分阈值为12.20%~13.82%。
关键词:黑果枸杞;干旱胁迫;光合色素;光合特性
干旱胁迫是限制植物生存和生长的关键因素,其在干旱和半干旱地区对植被的恢复会造成不利的影响[1]。干旱胁迫严重影响植物的形态结构、光合生长和代谢水平,植物只有适应这种干旱环境才能生存[2]。光合作用直接关系到植物的生长发育、产量形成以及次生代谢物质的合成积累,被认为是自然条件下限制植物生长,影响植物生产力的最重要因子之一[3]。光合作用对干旱胁迫更为敏感,它不仅受气孔导度下降的限制,而且受严重胁迫时叶绿体水平破坏的限制[4-5]。面对干旱胁迫,植物一般通过各种保护措施抵抗胁迫,或通过自身修复能力缓解胁迫所造成的危害[6]。因此,从光合特性方面研究植物对干旱胁迫的响应与适应特征,对深入探讨植物适应干旱的能力及对策显得越来越重要。
黑果枸杞(LyciumruthenicumMurr.)为茄科枸杞属多棘刺灌木,主要分布于中国西部地区,是中国荒漠地区地带性植被的主要建群树种[7],也是一种集药用、绿化和水土保持价值为一体的野生优良植物[8],果实富含蛋白质、枸杞多糖等多种营养成分,药用、保健价值远远高于普通红枸杞,被誉为植物“软黄金”[9]。随着该物种药用价值的进一步开发利用,野生资源破坏严重,种群数量大面积减少,部分地区甚至出现成片死亡现象,该物种当前已被列为重要保护植物[10]。在这种严酷的形势下,对黑果枸杞进行有效的管护、保护和研究是目前黑果枸杞合理经营的主要任务。迄今为止,有关黑果枸杞的研究报道较少,Chen等[11]、何芳兰等[12]、王桔红等[13]、韩多红等[14]分别从盐胁迫、干旱胁迫及贮藏方式等方面对黑果枸杞种子萌发进行了研究,但目前就黑果枸杞幼苗对干旱环境适应能力和适应机制的研究还十分缺乏。野外调查发现,黑果枸杞结实量大,实生苗数量少,干旱可能是限制黑果枸杞幼苗生长发育和定居的关键因素。在干旱环境下,黑果枸杞是否可以通过调整光合生理的变化去适应干旱环境,此类研究还未见报道,而这可以从根本上阐明黑果枸杞幼苗适应干旱环境的机理。为此,本研究通过盆栽实验,探讨干旱胁迫对当年生黑果枸杞幼苗光合生理的影响,旨在揭示黑果枸杞对干旱胁迫的适应能力和适应机制,为黑果枸杞植被恢复提供理论依据。
1材料和方法
1.1实验地概况
实验地位于甘肃省张掖市(37°28′N,97°20′E)河西学院农学实习基地,属大陆性气候,年平均气温6 ℃,年日照时数3 106 h,全年无霜期138~179 d,年太阳辐射量6 140~6 270 mJ/m2,年平均降水量113~120 mm,年蒸发量为2 291 mm。
1.2干旱胁迫处理
以当年生黑果枸杞盆栽实生苗为供试材料,其种子采自张掖市甘州区野生黑果枸杞植株。2014年3月10日播种在塑料盆内,盆高20 cm,内径23 cm,土壤为土壤、砂质土、腐殖质(1∶2∶1)的混合土,每盆装入等量的土,每盆播10粒种子,共40盆。待幼苗生长至2~3片真叶时,每盆留2株健壮的幼苗。为保证幼苗健康生长,育苗期间土壤含水量约为田间最大持水量的80%。2014年6月27日开始干旱胁迫实验,设对照(CK,土壤含水量为32.96%~35.35%)、轻度干旱胁迫(T1,土壤含水量为21.18%~22.32%)、中度干旱胁迫(T2,土壤含水量为12.20%~13.82%)和重度干旱胁迫(T3,土壤含水量为7.89%~8.73%)4各处理水平。每个处理重复3次。胁迫期间用称重法补充损失的水分,采取人工防雨措施,以保证土壤水分含量稳定。胁迫30 d后,取样测定其光合色素含量、光合作用参数和叶绿素荧光参数。
1.3测定指标及方法
1.3.1光合色素含量取幼苗冠层上部完全展开的叶片,新鲜叶立即在低温、避光条件下带回实验室进行色素含量测定。将新鲜叶片剪碎混匀,取0.1 g用80%的丙酮在低温、黑暗条件下浸泡使叶片完全变白。浸提液过滤、定容后在波长646、663和470 nm下比色,分别测定叶绿素a(Chl a)、叶绿素b(Chl b)和类胡萝卜素(Car)的吸收值,计算Chl a、Chl b、Chl(a+b)、Car的含量及Chl(a/b)、Chl(a+b)/Car 之比[15]。
1.3.2光合生理参数选择连续晴好的天气,利用CIRAS-2光合仪于9:00~11:30测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci),根据各处理数据,计算水分利用效率(WUE)和气孔限制值(Ls)。
瞬时水分利用效率:WUE=Pn/Tr
气孔限制值:Ls=(1-Ci/Co)×100%(Co为空气中CO2浓度)
1.3.3叶绿素荧光参数使用FMS-2荧光仪测定叶绿素荧光参数,选取植株中上部受光一致的功能叶3片,叶片暗适应25 min后,测定初始荧光(F0);随后给一个强闪光(6 000 μmol·m-2·s-1),脉冲时间(0.8 s)测定最大荧光(Fm);测定充分暗适应的PSⅡ最大光化学效率(Fv/Fm)。接着照射饱和脉冲光(6 000 μmol·m-2·s-1),测定光下最大荧光(Fm′)、Ft和实际光化学效率(ΦPSⅡ)。根据各处理数据,计算光化学猝灭系数(qP)和非光化学猝灭系数(qN)。
qP=(Fm′-Ft)/(Fm′-F0)
qN=(Fm-Fm′)/(Fm-F0)
1.4数据处理
采用SPSS 16.0对数据进行one-way ANOVA分析,采用LSD进行多重比较。用Origin 8.0软件作图。
2结果与分析
2.1干旱胁迫对黑果枸杞叶片光合色素含量的影响
从表1中可以看出,黑果枸杞幼苗叶片叶绿素a、叶绿素b均随干旱胁迫强度的增加逐渐降低,且各处理间差异显著(P<0.05);幼苗叶片叶绿素含量(Chl a+b)也随干旱胁迫强度的增加逐渐降低,且各处理间差异显著(P<0.05),轻度(T1)、中度(T2)、重度(T3)干旱胁迫下的叶绿素含量较CK分别显著下降了2.88%、10.17%和13.63%。同时,黑果枸杞幼苗叶片类胡萝卜素含量也随干旱胁迫强度的增加逐渐降低,且各处理间差异显著(P<0.05),轻度、中度、重度干旱胁迫下的类胡萝卜素含量较CK分别显著下降了12.77%、34.04%和42.55%。另外,光合色素含量比值Chl(a/b)、Chl(a+b)/Car均随干旱胁迫强度的增加逐渐增加,轻度、中度、重度干旱胁迫处理下Chl(a/b)较CK分别增加了0.00%、0.79%和4.72%,三者的Chl/Car较CK分别显著增加了11.25%、34.10%和49.38%。可见,干旱胁迫显著降低了黑果枸杞幼苗叶片光合色素含量,且胁迫程度越重下降幅度越大;类胡萝卜素含量比叶绿素含量降低幅度更大,叶绿素b又比叶绿素a含量降低幅度大。
2.2干旱胁迫对黑果枸杞幼苗叶片光合作用参数的影响
在不同程度干旱胁迫条件下,黑果枸杞幼苗叶片净光合速率、蒸腾速率和气孔导度随土壤含水量降低的变化趋势相同(图1)。其中,在轻度干旱胁迫下,黑果枸杞幼苗叶片净光合速率、蒸腾速率和气孔导度较CK略有增加,但差异不显著(P>0.05);在中度和重度干旱胁迫下,黑果枸杞幼苗叶片光合速率、蒸腾速率和气孔导度均较CK显著降低,在中度干旱胁迫下降幅分别为17.06%、21.64%和44.12%,在重度干旱胁迫下降幅分别为35.88%、40.07%和72.55%(P<0.05)。同时,随着土壤含水量的降低,黑果枸杞幼苗叶片胞间CO2浓度逐渐增加,但在轻度和中度干旱胁迫下与CK无显著差异(P>0.05),仅在重度干旱胁迫下较CK显著增加15.50%(P<0.05)。另外,随着土壤含水量的降低,黑果枸杞幼苗叶片水分利用效率呈先增加后降低并趋于平稳的趋势,但各处理间差异不显著(P>0.05);而此时其叶片气孔限制值随土壤含水量的降低而逐渐降低,各处理较CK降幅在16.14%~56.90%之间,但差异均未达到显著水平(P>0.05)。>可见,黑果枸杞幼苗叶片各光合生理参数在轻度干旱胁迫下均未受到显著影响,而随干旱胁迫加重,净光合速率、蒸腾速率和气孔导度显著降低,胞间CO2浓度在重度干旱胁迫下也显著降低,而其余参数未受到显著影响。
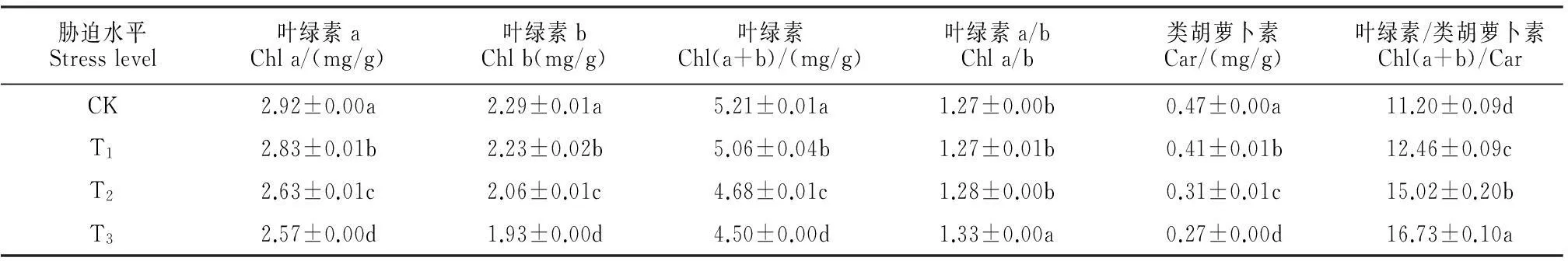
表1 干旱胁迫下黑果枸杞幼苗光合色素含量的变化
注:CK、T1、T2、T3分别表示对照及轻度、中度和重度干旱胁迫处理;同列不同字母表示处理间在0.05水平存在显著性差异;下同。
Note:CK,T1,T2and T3stand for control,mild drought stress,moderate drought stress and severe drought stress,respectively;The different normal letters indicate significant difference among treatments at 0.05 level;The same as below.
2.3干旱胁迫对黑果枸杞幼苗叶片叶绿素荧光参数的影响
图2显示,随干旱胁迫程度的加剧(土壤含水量降低),黑果枸杞幼苗叶片初始荧光(F0)逐渐增加,而同期的最大荧光(Fm)逐渐降低,且在中度和重度干旱胁迫下与CK差异均达到显著水平(P<0.05)。同时,黑果枸杞幼苗叶片PSⅡ最大光化学效率(Fv/Fm)和实际光化学效率(ΦPSⅡ)在干旱胁迫下均逐渐降低,轻度、中度和重度干旱胁迫处理的Fv/Fm分别较CK显著降低了6.80%、6.93%和13.31%,而其ΦPSⅡ则较CK分别显著降低了7.12%、20.04%和34.65%(P<0.05)。另外,黑果枸杞幼苗叶片光化学猝灭系数(qP) 随土壤含水量的降低逐渐降低,而同期非光化学猝灭系数(qN)却逐渐增加,轻度、中度和重度干旱胁迫处理的qP较CK分别降低了2.13%、6.75%和13.42%,其相应的qN值较CK分别增加了8.63%、41.69%和57.10%,且后两者变化均达到显著水平(P<0.05)。以上结果说明轻度干旱胁迫下黑果枸杞幼苗叶片叶绿素荧光参数PSⅡ最大光化学效率和实际光化学效率受显著影响外,其他参数均未受影响。而在重度干旱胁迫下最大荧光、光化学猝灭系数显著降低。
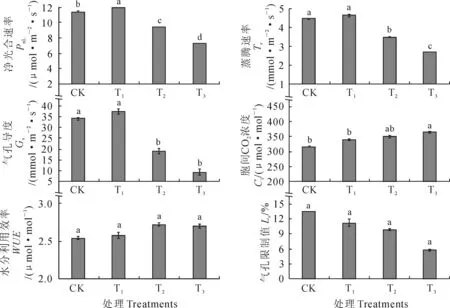
图1 不同干旱胁迫下黑果枸杞幼苗叶片光合参数的变化
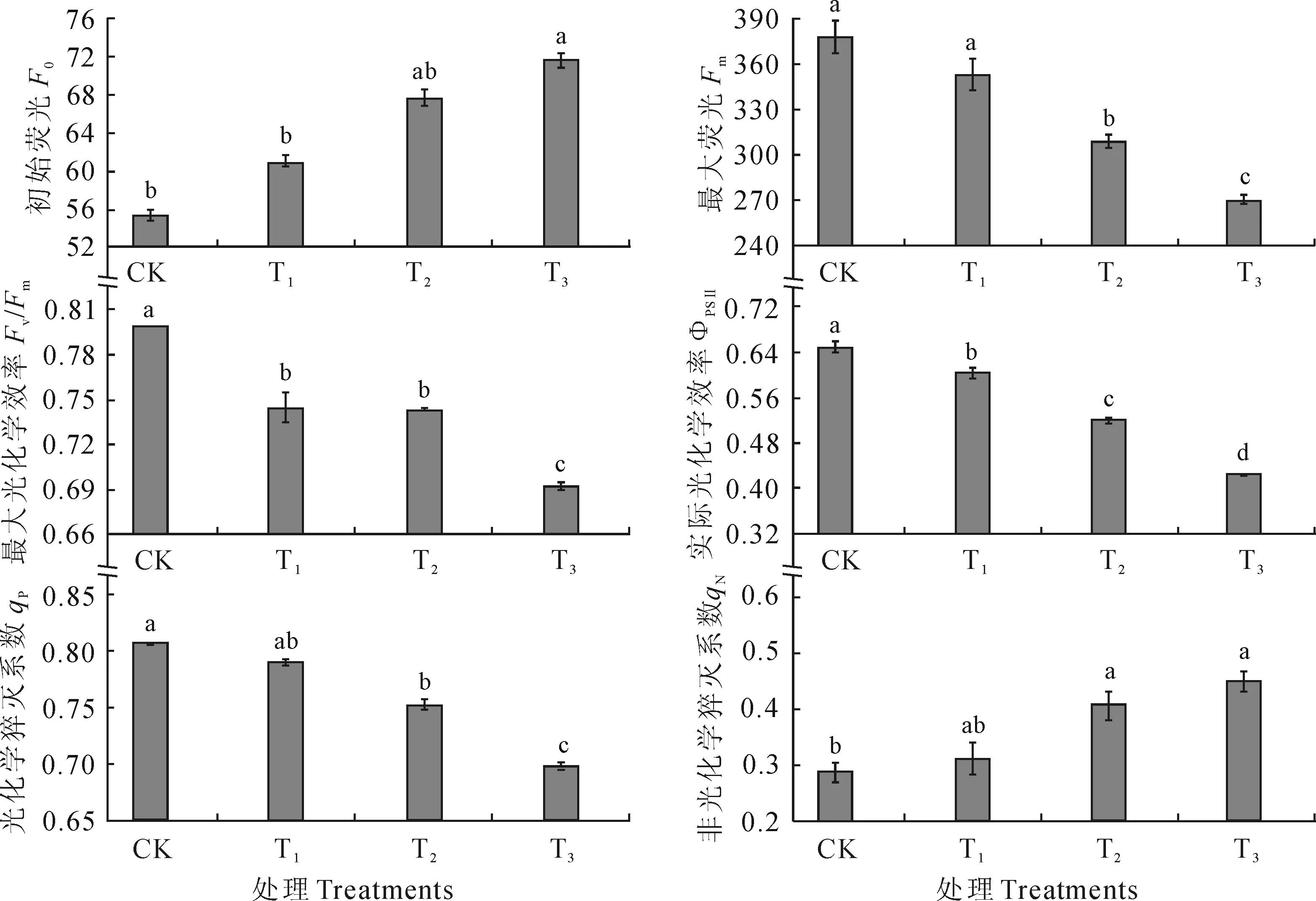
图2 不同干旱胁迫下黑果枸杞幼苗叶片叶绿素荧光参数的变化
3结论与讨论
干旱胁迫是植物的生理功能由最初失去稳定到最后趋于正常并增加抗性的一种需要。干旱胁迫影响植物许多重要的生理生态过程,其中光合生理是重要的参数[16]。本研究表明,随着干旱胁迫强度的增加,黑果枸杞幼苗叶绿素及类胡萝卜素含量均显著降低。这与前人对白刺花[17]和迷迭香[18]等植物的研究结果一致。这主要是由于干旱胁迫通过抑制叶绿素合成,并加速其分解,导致叶绿素含量直线下降,而干旱胁迫下类胡萝卜素的降低是为了清除叶绿体中活性氧,防止膜脂过氧化[19]。干旱胁迫影响黑果枸杞幼苗光合色素格局的变化,也暗示了其光合器官生理活性的变化。在干旱胁迫条件下,黑果枸杞幼苗Chl(a/b)增加,这是PSⅡ聚光复合体中LHCII含量减少的主要特征[20],说明干旱胁迫下叶片捕光蛋白色素复合物的降解程度高于反应中心的降解程度[21],这种响应可减少黑果枸杞叶片对光能的捕获,降低光合机构遭受光氧化破坏的风险,是其适应干旱胁迫的一种光保护调节机制。
同时,影响植物光合作用的因素可分为气孔因素和非气孔因素两类,气孔因素是水分胁迫导致气孔导度下降,CO2进入叶片受阻而使光合速率下降,而非气孔因素是叶肉细胞的光合活性下降[22]。Farquhar等认为,当Pn和Ci变化方向相同时,两者同时减小,Pn的下降主要是由Gs引起的气孔因素所致,否则Pn的下降要归因于叶肉细胞羧化能力的降低[23]。在本研究的轻度干旱胁迫下,Gs值相对对照没有发生明显变化,这说明轻度干旱胁迫下黑果枸杞叶片气孔能够保持一定水平的开度,利于其蒸腾和光合过程进行;而在中度和重度干旱胁迫下,黑果枸杞叶片Pn和Gs伴随着Ci的上升而下降,这表明在此干旱胁迫条件下黑果枸杞光合作用主要受非气孔因素的限制。
另外,叶绿素荧光动力学参数是干旱逆境对植物光合作用影响研究和探测的理想方法[24]。叶绿素荧光与光合作用中各种反应过程密切相关,任何环境因子对光合作用的影响都可通过叶绿素荧光动力学反映出来[25]。本研究结果表明,随着水分胁迫的加剧,黑果枸杞的Fm逐渐下降,而F0逐渐升高,这表明其光合色素吸收的光能中,以热和荧光的形式散失的能量在增加。这与前人在条墩桑[26]、小冠花[27]和丹参[28]上的相关研究结果相同。Fv/Fm反映了开放的PSⅡ 反应中心捕获激发能的效率,是研究植物胁迫的重要参数,任何影响PSⅡ效能的环境胁迫均会使Fv/Fm降低[26]。随着干旱胁迫的加剧,黑果枸杞的Fv/Fm逐渐降低,这表明干旱胁迫使黑果枸杞PSⅡ原初光能转化率下降,潜在活性中心受损,黑果枸杞叶片光合作用的原初反应受到抑制,这与在丹参[28]、草莓[29]上的研究结果相似。qP在一定程度上反映了PSⅡ 反应中心的开放程度,而qN是植物PSⅡ天线色素所吸收的光能没有被用于光化学反应,而以热的形式耗散掉的光能部分,当PSⅡ反应中心的天线色素吸收的光能过剩时,如不能及时的加以耗散,将会对光合机构造成破坏或失活[30]。随着干旱胁迫程度的增加,本研究中黑果枸杞叶片qP、ΦPSⅡ逐渐降低,但qN逐渐增加,这表明黑果枸杞叶片过多的能量以热的形式被耗散,反应中心开放程度降低,以避免PSⅡ 反应中心受到损伤。
综上所述,在干旱胁迫下,黑果枸杞幼苗通过增加Chl a/b值以减少叶片对光能的捕获,降低光合机构遭受光氧化破坏的风险。当土壤含水量低于13.82%时,黑果枸杞光合作用降低的主要原因是非气孔因素的限制;随干旱胁迫程度的增加,黑果枸杞幼苗通过降低qP、ΦPSⅡ和增加qN以降低反应中心的开放程度,避免PSⅡ 反应中心受到损伤。黑果枸杞生长所允许的最大土壤水分亏缺为7.89%,维持黑果枸杞具有较高的WUE和Pn的土壤水分阈值为12.20%~13.82%。
参考文献:
[1]LI F L,BAO W K,WU N.Effects of water stress on growth,dry matter allocation and water-use efficiency of a leguminous species,Sophoradavidii[J].AgroforestSyst,2008,77:193-201.
[2]安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干旱胁迫的生长与生理响应[J].生态学报,2011,31(3):716-725.
AN Y Y,LIANG Z S,HAO W F.Growth and physiological responses of thePeriplocasepiumBunge seedlings to drought stress[J].ActaEcologicaSinica,2011,31(3):716-725.
[3]蔡海霞,吴福忠,杨万勤.干旱胁迫对高山柳和沙棘幼苗光合生理特征的影响[J].生态学报,2011,31(9):2 430-2 436.
CAI H X,WU F Z,YANG W Q.Effects of drought stress on the photosynthesis ofSalixparaqplesiaandHippophaerhamnoidesseedlings[J].ActaEcologicaSinica,2011,31(9):2 430-2 436.
[4]BOYER T ,WONG S,FARQUHAR G.CO2and water vapour exchange across the leaf cuticle (epidermis) at various water potentials[J].PlantPhysiology,1997,114:185-189.
[5]LAWLOR D.Limitation to photosynthesis in water stressed leaves:stomata vs metabolism and the role of ATP[J].AnnalsofBotoby,2002,89:871-885.
[6]COOPER K,FARRANT J.Recovery of the resurrection plant Crate rostigma wilmsii from desiccation:protection versus repair[J].JournalofExperimentalBotany,2002,53:1 805-1 813.
[7]路兴慧.塔里木河下游五种典型荒漠植物水分生理及自维持特性研究[D].乌鲁木齐:新疆农业大学,2009.
[8]姜霞.黑果枸杞耐盐机理的相关研究[D].陕西杨陵:西北农林科技大学,2012.
[9]冯建森,刘志虎.酒泉市野生黑果枸杞资源及利用[J].林业实用技术,2013,(2):62-64.
FENG J S,LIU Z H.LyciumruthenicumMurr resources and use of Jiuquan city[J].PracticalForestryTechnology,2013,(2):62-64.
[10]LIU Z G,SHU Q Y,WANG L,etal.Genetic diversity of the endangered and medically importantLyciumruthenicumMurr.revealed by sequence-related amplified polymorphism (SRAP) markers[J].BiochemicalSystematicsandEcology,2012,45:86-97.
[11]CHEN H K,ZHAO W H.Effect of NaCl stress on seed germination ofLyciumruthenicumMurr.[J].AgricultureScienceandTechnology,2010,11(4):37-38.
[12]何芳兰,赵明,王继和,等.几种荒漠植物种子萌发对干旱胁迫的响应及其抗旱性评价研究[J].干旱区地理,2011,34(1):100-106.
HE F L,ZHAO M,WANG J H,etal.Response to droughty stresses and drought resistances evaluation of seed germination of four desert vegetation[J].AridLandGeography,2011,34(1):100-106.
[13]王桔红,马瑞君,陈文.冷层积和室温干燥贮藏对河西走廊8种荒漠植物种子萌发的影响[J].植物生态学报,2012,36(8):791-801.
WANG H H,MA R J,CHEN W.Effects of cold stratification and dry storage at room temperature on seed germination of eight desert species from the Hexi Corridor of China[J].ChineseJournalofPlantEcology,2012,36(8):791-801.
[14]韩多红,李善家,王恩军,等.外源钙对盐胁迫下黑果枸杞种子萌发和幼苗生理特性的影响[J].中国中药杂志,2014,29(1):34-39.
HAN D H,LI S J,WANG E J,etal.Effect of exogenous Calcium on seed germination and seedling physiological characteristics ofLyciumruthenium[J].ChinaJ.Chin.MaterMed.,2014,29(1):34-39.
[15]李芳兰,包维楷,吴宁.白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J].生态学报,2009,29(10):5 406-5 416.
LI F L,BAO W K,WU N.Morphological and physiological responses of currentSophoradavidiiseedlings to drought stress[J].ActaEcologicaSinica,2009,29(10):5 406-5 416.
[16]周生荟,刘玉冰,谭会娟,等.荒漠植物红砂在持续干旱胁迫下的光保护机制研究[J].中国沙漠,2010,30(1):69-73.
ZHOU S H,LIU Y B,TAN H J,etal.The photoprotective mechanism of desert plantReaumuriasoogoricaunder progressive soil drying[J].JournalofDesertResearch,2010,30(1):69-73.
[17]WU F Z,BAO W K,LI F L,etal.Effects of drought stress and N supply on the growth,biomass partitioning and water-use efficiency ofSophoradavidiiseedlings[J].EnvironExp.Bot.,2008,63:248-255.
[18]GRATANI L,VARONE L.Leaf key traits ofEricaarboreaL.,EricamultifloraL.andRosmarinusofficinalisL.co-occurring in the Mediterranean maquis[J].Flora,2004,199:58-69.
[19]WANG H M,BAO W K,LI F L.Physiological and biochemical responses of two years oldSophoradavidiiseedling leaves to different water stresses[J].ChineseJournalAppliedandEnvironmentalBiology,2008,14(6):757-762.
[20]ANDERSON J M,ARO E M.Grana staeking and proteetion of photosystem 11 in thylakoid membranes of higher plant leaves under sustained high irradianee:anhy pothesis[J].PhotosynthesisResearch,1994,41(2):315-332.
[21]LAREHER W.Physiological Plant Ecology[M].4th Ed.New York:Springer,2003.
[22]裴斌,张光灿,张淑勇,等.土壤干旱胁迫对对沙棘叶片光合作用和抗氧化物酶活性的影响[J].生态学报,2013,33(5):1 386-1 396.
PEI B,ZHANG G C,ZHANG S Y,etal.Effects of soil drought stress on photosynthetic characteristics and antioxidant enzyme activities inHippophaerhamnoidesLinn.seedings[J].ActaEcologicaSinica,2013,33(5):1 386-1 396.
[23]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].AnnualReviewofPlantPhysiology,1982,33:317-345.
[24]李娟,彭镇华,高健,等.干旱胁迫下黄条金刚竹的光合和叶绿素荧光特性[J].应用生态学报,2011,22(6):1 395-1 402.
LI J,PENG Z H,GAO J,etal.Photosynthetic parameters and chlorophyll fluorescence characteristics ofPleioblastuskongosanensisf.aureostriausunder drought stress[J].ChineseJournalofAppliedEcology,2011,22(6):1 395-1 402.
[25]惠红霞,许兴,李前荣.外源甜菜碱对盐胁迫下枸杞光合功能的改善[J].西北植物学报,2003,23(12):2 137-2 422.
HUI H X,XU X,LI Q R.Exogenous betaine improves photosynthesis ofLyciumbarbarumunder salt stress[J].ActaBot.Boreal.-Occident.Sin.,2003,23(12):2 137-2 422.
[26]闫海霞,方路斌,黄大庄.干旱胁迫对条墩桑生物量分配和光合特性的影响[J].应用生态学报,2011,22(12):3 365-3 370.
YAN H X,FANG L B,HUANG D Z.Effects of drought stress on the biomass distribution and photosynthetic characteristics of cluster mulberry[J].ChineseJournalofAppliedEcology,2011,22(12):3 365-3 370.
[27]杨文权,顾沐宇,寇建村,等.干旱及复水对小冠花光合及叶绿素荧光参数的影响[J].草地学报,2013,21(6):1 130-1 135.
YANG W Q,GU M Y,KOU J C,etal.Effect of drought and rewatering on the photosynthesis and chlorophyⅡ fluorescence ofCoronillavaria[J].ActaAgrestiaSinica,2013,21(6):1 130-1 135.
[28]罗明华,胡进耀,吴庆贵,等.干旱胁迫对丹参叶片气体交换和叶绿素荧光参数的影响[J].应用生态学报,2010,21(3):619-623.
LUO M H,HU J Y,WU Q G,etal.Effects of drought stress on leaf gas exchange and chlorophyll fluorescence ofSalviamiltiorrhiza[J].ChineseJournalofAppliedEcology,2010,21(3):619-623.
[29]吴甘霖,段仁燕,王志高,等.干旱和复水对草莓叶片叶绿素荧光特性的影响[J].生态学报,2010,30(14):3 941-3 946.
WU G L,DUAN R Y,WANG Z G,etal.Effects of drought stress and rehydration on chlorophyll fluorescence characteristics inFragaria×ananassaDuch[J].ActaEcologicaSinica,2010,30(14):3 941-3 946.
[30]KRAUSE G H,WEIS E.Chlorophyll fluorescence and photosynthesis:the basics[J].AnnualReviewofPlantPhysiologyandPlantMolecularBiology,1991,42:313-349.
(编辑:裴阿卫)
Effect of Drought Stress on Photosynthesis Characteristics ofLyciumruthenicumSeedlings
GUO Youyan1,LIU Hongjun2,KONG Dongsheng1*,YAN Fang1,ZHANG Yajuan1,LIU Donghua1
(1 Hexi College,Zhangye,Gansu 734000,China;2 Zhangye Seeding Managment Stations,Zhangye,Gansu 734000,China)
Abstract:To detect the effect of drought on one-year-old Lycium ruthenicum seedlings,we investigated the photosynthetic pigments,photosynthetic characteristics and chlorophyll fluorescence characteristics on four different soil contents:CK(21.18%-22.32% soil moiture),light stress(12.20%-13.82%),moderate stress(12.20%-13.82%) and severe stress(7.89%-8.73%).The results showed that:(1)the chlorophyll content and carotenoid content decreased with the decreasing soil water content.(2)Under the moderate stress,the net photosynthetic rate (Pn),the transpiration rate (Tr),stomatal conductance (Gs) decreased significantly.With the decreasing of soil water content,intercellular CO2concentration (Ci),the water use efficiency (WUE) increased,whereas stomatal limitation (Ls) performed the inverse trend.(3)With the increasing of the minimal fluorescence (F0) and nonphotochemical quenching (qspan) value,the maximal fluorescence (Fm),maximal photochemical efficiency (Fv/Fm),actual photochemical efficiency (ФPSⅡ),and photochemical quenching (qspan) decreased.It indicated that the excess energy of the leaves of L.ruthenicum is dissipated by heat,and the open degree of the reaction center is reduced to avoid the damage of the PSⅡ reaction center.It suggested that L.ruthenicum keep on vigor above the limitation of 7.89% soil moisture,and the largest WUE and the higher Pnarrive on 12.20%-13.82% soil moisture.
Key words:Lycium ruthenicum Murr.;drought stress;photosynthetic pigments;photosynthetic characteristics
中图分类号:Q945.79
文献标志码:A
作者简介:郭有燕(1980-),女,博士,副教授,主要从事天然林保护与利用研究。E-mail:guoyouyan_2008@163.com*通信作者:孔东升,研究员,主要从事植物生态植被恢复研究。E-mail:Kongdsh@sohu.com
基金项目:国家自然科学基金项目(31460189);甘肃省高等学校科研项目(2014A-111)
收稿日期:2015-11-12;修改稿收到日期:2015-12-28
文章编号:1000-4025(2016)01-0124-07
doi:10.7606/j.issn.1000-4025.2016.01.0124
