牛角瓜大小孢子发生及雌雄配子体发育研究
2016-03-23张太奎王连春罗明灿刘惠民
刘 鹏,张太奎,李 璐,刘 婷,王连春,罗明灿,刘惠民
(西南林业大学 林学院,昆明 650224)
牛角瓜大小孢子发生及雌雄配子体发育研究
刘鹏,张太奎,李璐,刘婷,王连春,罗明灿,刘惠民*
(西南林业大学 林学院,昆明 650224)
摘要:为从胚胎学方面解释牛角瓜坐果率低的原因,该实验以牛角瓜(Calotropis gigantea L.)不同时期的新鲜花朵为试材,采用石蜡切片法,观察牛角瓜大小孢子发生及雌雄配子体发育过程。结果显示:(1)牛角瓜小孢子孢原细胞起源于表皮细胞下面的分生组织并在两个地方分化;花药壁由外向内由1层表皮、1层药室内壁、2层中层和1层绒毡层组成,花药壁发育属于基本型,绒毡层属于分泌型;小孢子形成时胞质分裂方式为连续型,成熟花粉粒为3-细胞型复合花粉,有2个较小的精细胞和1个较大的营养细胞,精细胞呈凸透镜型。(2)牛角瓜的雌蕊具有2心皮,子房1室,边缘胎座,胚珠倒生,单珠被,薄珠心;大孢子母细胞由1个孢原细胞直接发育而成,合点端大孢子分化成为功能大孢子,由功能大孢子发育为7细胞8核的蓼型胚囊。(3)牛角瓜雌蕊发育晚于雄蕊,花蕾开放前雌雄蕊发育趋于同步。研究表明,虽然在雌雄配子体发育过程中存在败育现象,但大部分雌雄配子体发育正常。
关键词:牛角瓜;大小孢子发生;雌雄配子体发育
牛角瓜(CalotropisgiganteaL.)属于萝藦科(Asclepiadaceae),为直立灌木或小乔木,主要分布于东南亚国家和地区,在中国见于云南、海南、广东、四川等南方省区。牛角瓜在棉纺原料、药用、能源等方面用途广泛。国内外关于牛角瓜的研究较少,主要集中于牛角瓜的组培苗的无性繁殖技术[1]、种子萌发[2]、纤维素开发[3-4]、药用[5-6]、固体生物燃料[7]以及化合物的提取[8]等方面。
萝藦科植物因其独特的花部结构而被关注[9],具有副花冠、匙状花药、花粉块、合蕊柱等特殊花部结构,在自然条件下,一般萝藦科植物花量大,但坐果率很低,仅在1.33%~5.0%之间[10]。造成坐果率低的因素有很多,如:传粉生物特征,或胚胎发育过程中的败育现象等。关于萝藦科植物的胚胎学研究鲜有报道,牛角瓜的胚胎学研究也非常少见。牛角瓜周年开花结果,但因其独特的花部特征,授粉位置不在柱头表面,而在柱头腔中,花粉块很难到达柱头腔完成授粉,导致“花多果少”,制约了以利用其种毛纤维为主的牛角瓜栽培业的发展[11]。本实验对牛角瓜大、小孢子发生及雌雄配子体的发育进行观察研究,从胚胎学方面探讨其坐果率低的原因,为牛角瓜的胚胎学以及传粉生物学提供基础资料,并为解决牛角瓜的繁殖问题提供理论依据。
1材料和方法
牛角瓜采自云南元江县种苗站。于2013年10月份和2014年4月份分别采集牛角瓜不同发育阶段的花蕾和花,现场用FPA固定液固定并带回实验室保存。2014年5月~10月,采用常规石蜡切片法,将固定材料经选材、修材后进行逐级Johannsen Gem脱水液脱水、浸蜡、包埋、切片(切片厚度为8 μm)、脱蜡、爱氏苏木精染色、加拿大树胶封片制成永久切片。观察牛角瓜大小孢子发生与雌雄配子体发育过程。然后选择典型制片在Axiovert 200M显微镜下观察各个发育时期的具体特征,并照相。
2结果与分析
2.1小孢子发生和雄配子体发育
2.1.1花药壁的发育牛角瓜是两性花,每朵花内有雄蕊5枚,每枚雄蕊具2个花粉块。花药原基是由雄蕊原基顶端发育而来,它的最外层为1层表皮细胞,里面主要为一团分生组织,随后分化出孢原细胞,孢原细胞经一次平周分裂,形成初生壁细胞和初生造孢细胞(图版Ⅰ,A)。初生壁细胞继续进行平周分裂(图版Ⅰ,B、C)和多次垂周分裂(图版Ⅰ,D、E),产生呈同心排列的数层细胞,自外向内依次为1层药室内壁、2层中层和1层绒毡层,它们与花药表皮共同构成牛角瓜的花药囊壁。由于牛角瓜的次生壁细胞外层发育为药室内壁和中层,内层则发育为中层和绒毡层(图版Ⅰ,F、G),具有2层中层,根据Davis[12]对被子植物花药壁发育标准的划分,牛角瓜的花药壁发育属于基本型。
(1)表皮表皮层细胞随着花药的不断扩大,而进行切向加长生长,变为扁平(图版Ⅰ,H);小孢子母细胞减数分裂期,表皮层细胞扁长形,并出现角质层(图版Ⅰ,I);到花药成熟,表皮层细胞的角质层进一步加厚(图版Ⅰ,O、P)。
(2)药室内壁小孢子母细胞时期,药室内壁细胞呈扁长方形,沿圆周方向排列,细胞液泡化,细胞核位于细胞中间(图版Ⅰ,F、G);小孢子母细胞减数分裂时期,药室内壁开始径向生长(图版Ⅰ,H);二核花粉时期,药室内壁进一步径向延长(图版Ⅰ,M、N),到花粉成熟时转变为纤维层(图版Ⅰ,O、P)。
(3)中层在花药的整个发育过程中,中层存在的时间较为短暂。小孢子母细胞减数分裂时期,中层开始解体(图版Ⅰ,H),逐渐被周围细胞吸收(图版Ⅰ,I);小孢子收缩期,中层只留下残迹(图版Ⅰ,J)。
(4)绒毡层绒毡层细胞垂直长轴沿圆周方向排列,绒毡层细胞初期,体积较小,排列紧密,呈长方形,单核;小孢子母细胞形成过程中,绒毡层细胞体积和细胞核逐渐增大(图版Ⅰ,F、G);小孢子减数分裂时期,绒毡层细胞继续径向生长,发育达到顶峰,细胞较大,细胞质浓,绒毡层细胞具有多核(图版Ⅰ,H);在小孢子减数分裂后期,绒毡层开始原位解体(图版Ⅰ,I),随着花药的发育,绒毡层进一步解体(图版Ⅰ,J~M);至花粉成熟时,绒毡层完全溶解消失,只剩下表皮细胞和药室内壁(图版Ⅰ,O、P)。
绒毡层细胞长轴沿径向规则地排列于花粉囊内侧,直接与花粉囊内的细胞毗连,是整个花药发育时期最为活跃的一层。绒毡层解体的过程,可为牛角瓜花粉粒的发育提供营养物质和结构物质,且具有分泌功能,分泌物包围花粉粒,因此牛角瓜的绒毡层是分泌型绒毡层[10]。
2.1.2小孢子发生花药在未分化时期,雄蕊原基分裂伸长,它的最外层为1层表皮细胞,里面主要为一团分生组织(图版 Ⅱ,A),随后分化出孢原细胞,其体积较大,染色深,径向延长并细胞核显著(图版 Ⅱ,B);孢原细胞经一次平周分裂(图版 Ⅱ,C),形成初生壁细胞和初生造孢细胞,初生造孢细胞体积大,核仁大而显著,多边形,稍后渐近圆形,细胞质浓厚,没有明显的液泡(图版 Ⅱ,D、E)。牛角瓜的初生造孢细胞不经分裂直接发育成小孢子母细胞,并行使小孢子母细胞的功能(图版Ⅱ,F~H),小孢子母细胞体积较大,细胞质浓厚,细胞核大,无明显的液泡,染色质和核仁形成染色较深的圆球体(图版Ⅱ,I~K)。
与大多数植物不同的是,牛角瓜的小孢子母细胞进一步发育,但并未彼此分离,依然紧密相连在一起,直接进入减数分裂时期。分别经过前期Ⅰ的细线期(图版Ⅲ,A)、偶线期(图版Ⅲ,B、C)、双线期(图版Ⅲ,D)和终变期(图版Ⅲ,E、F),以及中期Ⅰ(图版Ⅲ,G、H)、中后期Ⅰ(图版Ⅲ,I、J)、后期(图版Ⅲ,K、L)和末期(图版Ⅳ,A、B),形成二分体(图版Ⅳ,C)。紧接着细胞进入减数分裂Ⅱ期,依次经过分裂前期Ⅱ、中期Ⅱ、后期Ⅱ(图版Ⅳ,D),最终形成四分体(图版Ⅳ,E),但花粉粒没有互相分开,而是紧密的联系在一起。牛角瓜小孢子的孢质分裂方式为连续型。在分裂的过程中,不同药室间和同一药室内的小孢子母细胞发育基本上能保持一致。
2.1.3雄配子体发育牛角瓜小孢子呈不规则圆球形。经过短暂的收缩期之后(图版Ⅳ,F),细胞核位于中央,体积大,核仁明显,孢质浓厚,成为单核花粉(图版Ⅳ,G)。随后孢质液泡化,成为单核靠边期(图版Ⅳ,H)。小孢子核在贴近花粉壁的位置进行有丝分裂(图版Ⅳ,I),先形成两个细胞核,贴近花粉壁的为生殖核,靠着大液泡的为营养核,随后发生不均等的孢质分裂,形成2个大小悬殊的细胞,其中靠近花粉壁一侧的呈凸透镜状的小细胞,为生殖细胞,另一侧为营养细胞(图版Ⅳ,J)。生殖细胞进行有丝分裂,形成2个精细胞(精子)(图版Ⅳ,K),牛角瓜是以含有1个营养细胞和2个精细胞进行传粉的,为3-细胞型花粉(图版Ⅳ,L)。
2.2大孢子发生与雌配子体发育
2.2.1大孢子发生牛角瓜的大孢子发生晚于小孢子。牛角瓜雌蕊具2心皮,子房1室,具多枚倒生胚珠,边缘胎座。由胎座表皮下层的局部细胞进行分裂(图版Ⅴ,A),产生突起(图版Ⅴ,B),形成胚珠原基,胚珠原基的前端成为珠心,基部分化为株柄,珠心基部的表皮细胞快速分裂,产生环状突起,逐渐生长扩展为珠被,将珠心包围(图版Ⅴ,C)。
随着胚珠原基的发育,由薄壁细胞组成的珠心内部发生了变化,在近珠孔端的珠心表皮下分化出一个体积较大、细胞质浓厚、具有显著的细胞核的孢原细胞(图版Ⅴ,D)。孢原细胞不分裂,继续发育形成大孢子母细胞,直接行使大孢子母细胞的功能,此时的大孢子母细胞体积明显较大,核大,细胞质较浓,位于1层珠心表皮下,因此胚珠发育属于薄珠心型(图版Ⅴ,E)。随后大孢子母细胞进入减数分裂阶段,经第一次减数分裂形成二分体(图版Ⅴ,F、G),继而进行第二次减数分裂(图版Ⅴ,H)后形成的大孢子四分体呈线形排列(图版Ⅴ,I)。其四分体中位于珠孔端的3个大孢子相继退化(图版Ⅴ,J、K),仅合点端的1个发育为功能大孢子,以后发育为胚囊(图版Ⅴ,L)。
在大孢子发生过程中,珠被细胞分裂并逐渐向上生长,牛角瓜发展成为具6~8层细胞的单层珠被。在珠被形成的过程中,在珠心最前端的地方留下一条未愈合的孔道,形成了一个狭长的珠孔。
2.2.2雌配子体发育功能大孢子成为雌配子体的第一个细胞。功能大孢子纵向体积延伸并液泡化,进而发育为单核胚囊(图版Ⅵ,A)。单核胚囊经过一次核的有丝分裂(图版Ⅵ,B),形成二核胚囊,初始形成的2个核彼此相互靠近(图版Ⅵ,C),而后二核分别位于胚囊的珠孔端与合点端(图版Ⅵ,D、E)。二核胚囊体积增大的同时,核分别进行有丝分裂形成四核胚囊,两端分别有2个核(图版Ⅵ,F、G)。4个核分别进行有丝分裂形成8核胚囊(图版Ⅵ,I、J)。珠孔端4个核,合点端4个核,随后珠孔端1个核向下移动,合点端的1个核向上移动,并相互靠近,形成极核(图版Ⅵ,H)。成熟胚囊的完整形态,它由1个卵细胞、2个助细胞、3个反足细胞和1个中央细胞构成。
在显微镜下可观察到牛角瓜胚囊的珠孔端由1个卵细胞和2个助细胞组成的卵器。卵器3个细胞均较大,各具1核,原生质浓。2个助细胞并列位于珠孔端(图版Ⅵ,K),卵细胞较大呈卵圆形,细胞核靠近合点端,位于两助细胞之间,三者一起构成卵器。成熟的卵细胞接近洋梨形,核位于合点端,呈现明显的极性。胚囊合点端有体积较小的3个反足细胞,呈“品”字型排列(图版Ⅵ,L)。由2个并排的极核组成的中央细胞较其它细胞大,胚囊的中央大部分空间为中央细胞所占据,中央细胞高度液泡化。上述胚囊形成过程表明,牛角瓜的雌配子体是由单孢子发生的7细胞8核胚囊,胚囊的发育类型属典型的蓼型胚囊[13]。
3结论与讨论
3.1小孢子发生和雄配子体发育特征
花药是雄蕊的主要部分,通常由4个花粉囊组成,少数植物为2个。萝藦科植物的花药有4室的和2室的。本实验观察到牛角瓜的花药为2室。萝藦科从夹竹桃科分离出来,主要由于夹竹桃科植物的花药多为4室,花粉的转运机制和萝藦科植物不一样。萝藦科植物的花粉粒常以花粉块的形式存在,而夹竹桃科的花粉粒不形成花粉块,常以四分体的形式存在[14]。由于牛角瓜花药为2室且花粉黏合在一起形成花粉块,因此,从形态学上证明了牛角瓜从属于萝藦科。
牛角瓜的绒毡层由初生壁层细胞分裂形成,是花药壁的最内层细胞,包围着小孢子母细胞。牛角瓜小孢子母细胞形成过程以及减数分裂时期,绒毡层逐渐发育并达到高峰,细胞核分裂呈多核现象,这与八角莲[15]绒毡层的发育相似。而在此后牛角瓜花粉粒发育过程中,绒毡层逐渐解体,释放内含物,为小孢子和雄细胞的发育形成提供营养,并分泌物质,形成包围花粉块的膜,这与王定康等[16]观察青阳参的结果相一致。至花粉成熟时,绒毡层完全退化。由此可见,绒毡层活动状态与小孢子、雄配子体发育以及包围花粉粒膜的形成对营养物质的要求相一致。且牛角瓜的花药壁绒毡层具有分泌功能,包围花粉块的膜由绒毡层分泌形成。这在萝藦科植物中比较常见[16]。
在花药的形成过程中,被子植物小孢子的发生大多经历孢原细胞时期、初生造孢时期以及次生造孢时期,然后形成小孢子母细胞,如西畴含笑[17]、楸树[18]、烟草[19]等,与大多数植物先由初生造孢细胞分裂发育形成次生造孢细胞进而发育成小孢子母细胞不同,牛角瓜初生造孢细胞直接行使小孢子母细胞的功能,初生造孢细胞不分化形成次生造孢细胞。牛角瓜的小孢子,并不像其它植物[20-21]一样从四分体中游离出来,依然挤在一起,所以牛角瓜的花粉属于复合型花粉[13],复合花粉的形式有很多,目前已经发现13种不同形式的复合花粉[22],在兰科、杜鹃科、萝藦科、夹竹桃科以及灯心草科中较为常见。
目前,有关复合花粉的的研究报道较少。每花粉散落单元中的花粉粒数越多,同一果实中的种子拥有相同父本的可能性越大;与柱头粘着的花粉量越大,雄配子体之间的竞争越激烈;每子房中的胚珠数越多,雌配子体之间的竞争越激烈[23]。复合花粉的结合方式主要有3种[13]:(1)黏性的液体,通常是从绒毡层的退化中产生;(2)花粉外壁的孢粉素;(3)普通的壁层。本研究表明,牛角瓜的花粉主要通过黏性液体相结合,且由绒毡层分泌物形成的膜包裹,形成一个大的花粉单位,牛角瓜花粉以复合花粉的形式散落。由于牛角瓜传粉及受精过程较为特殊,需将由花粉粒聚集在一起的花粉块插入柱头腔中才能完成授粉[11],因此同一果实中拥有的父本是一样的。牛角瓜花粉块中花粉粒众多,每个子房有上百个胚珠,它们之间在授粉受精过程中是否存在竞争关系及其强度大小都值得进行进一步的研究。
植物雌雄配子体的发育特点在一定程度上影响着植物的授粉方式,如雌雄异熟避免了自花授粉,保证了异花授粉;雌雄蕊在空间上的分离使植物更倾向于异花授粉等[24]。牛角瓜雄配子体发育较雌配子体发育早,但在发育过程中,雌雄配子体发育趋于同步,在开花时雌雄配子体同时发育成熟,不存在雌雄异熟;且牛角瓜具有合蕊柱,但雄蕊低于柱头表面,存在空间隔离,且牛角瓜是虫媒花,不能通过风等进行传粉,保证了异花传粉;牛角瓜的传粉单位是由1个着粉腺、2个转运臂和2个花粉块组成,传粉昆虫若要顺利完成传粉,需要将传粉单位从花药囊中拔出[11]。切片发现牛角瓜药室内壁细胞发育在后期发生纤维性加厚,致使花药囊变大,逐渐与花粉块脱离,这种现象还未在其它文献中见报道过,这种特征更易使花粉块被传粉昆虫拔出并带走,是适应虫媒传粉的特征。
牛角瓜不同药室间和同一药室内的小孢子母细胞发育基本上能保持一致。关于萝藦科小孢子母细胞减数分裂过程的研究报道中,大多数报道小孢子在花粉块内的发育过程就像在本实验对牛角瓜观察的结果一样是同步进行的[25],但Deshpande等[26]在Leptadenia中发现小孢子母细胞形成营养细胞和生殖细胞的过程并不同时进行。
3.2大孢子发生和雌配子体发育特征
萝藦科植物胚胎的特征:倒生型、薄珠心和单珠被[25,27-28],这与本实验观察牛角瓜的结果一致。牛角瓜为2心皮1室子房,边缘胎座,胚珠倒生,单珠被,薄珠心。
牛角瓜的孢原细胞位于表皮下,直接行使大孢子母细胞的功能,这与王定康观察青阳参的实验结果相一致[29]。珠被原基像Biswas[25]在Daeminextensa中和Frye[26]在一些萝藦科植物中报道的一样,从珠心顶部附近长出并包围大孢子母细胞。珠心组织大约在4核胚囊阶段解体,这与Biswas[25]在D.extensa中报道的结果一致,而Younis[28]在Calotropisprocera中发现珠心组织是在2~4核阶段解体。在牛角瓜中只有合点端的大孢子经过3次连续的分裂形成8核胚囊,而Frye[27]在Asclepias中还发现其它大孢子发育成胚囊的情况。
通过对大量不同时期的花朵材料进行切片观察,发现牛角瓜的大小孢子发育过程中都存在不同程度的败育,但大多数仍能够发育成熟。可见牛角瓜的低坐果率并不是因为雌雄配子体发育不良造成的,而与后期的传粉受精过程有着密切的关系。因此有必要对牛角瓜传粉过程以及受精过程进行进一步研究,将有助于深入解答其“花多果少”的原因,同时对生产实践也具有一定的指导意义。
参考文献:
[1]李克烈,罗联忠,陈伟,等.牛角瓜的组织培养[J].广西农业生物科学,2007,26(3):247-249.
LI K L,LUO L Z,CHEN W,etal.Tissue culture ofCalortropisgigantean[J].JournalofGuangxiAgric.andBiol.Science,2007,26(3):247-249.
[2]高柱,王小玲,马焕成,等.干热河谷牛角瓜育苗技术及幼苗生长节律[J].云南农业大学学报,2012,27(4):503-507.
GAO Z,WANG X L,MA H C,etal.Breeding technology and seedling growth rhythm ofCalotropisgiganteanL.in dry-hot valley[J].JournalofYunnanAgriculturalUniversity,2012,27(4):503-507.
[3]TUNTAWIROON N,SAMOOTSAKORN P,THEERARAJ G.The environmental implications of the use ofCalotropisgiganteaas a textile fabric[J].Agriculture,Ecosystems&Environment,1984,11(3):203-212.
[4]ASHORI A,BAHREINI Z.Evaluation ofCalotropisgiganteaas a promising raw material for fiber-reinforced composite[J].JournalofCompositeMaterials,2009,43(11):1 297-1 304.
[5]KUMAR P S,CHEZHIAN A,etal.Computational selections of terpenes present in the plantCalotropisgiganteaas mosquito larvicide's by blocking the sterol carrying protein,AeSCP-2[J].BangladeshJournalofPharmacology,2012,7(1):1-5.
[6]MURUGAN T.Antimicrobial activity of leaves and latex extract of the herbal plantCalotropisgigantea(Erukku In Tamil)[J].Ijbpas,April,2012,1(3):261-270.
[7]李瑞,曾建立,王晓东,等.耐盐碱植物牛角瓜产能成分分析[J].过程工程学报,2007,7(6):1 217-1 220.
LI R,ZENG J L,WANG X D,etal.Energy-produced components inCalotropisgigantean[J].TheChineseJournalofProcessEngineering,2007,7(6):1 217-1 220.
[8]王茂媛.牛角瓜的生理活性成分研究[D].海口:海南大学,2008.
[9]ENDRESS M E,LIEDE-SCHUMANN S,MEVE U.Advances in apocynaceae:the enlightenment,an introduction[J].AnnalsoftheMissouriBotanicalGarden,2007,94(2):259-267.
[10]WYATT R,BROYLES S B.Ecology and evolution of reproduction in milkweeds[J].AnnualReviewofEcologyandSystematics,1994,25:423-441.
[11]刘鹏,刘惠民,王连春,等.牛角瓜花部综合特征与繁育系统[J].园艺学报,2015,42(10):2 002-2 014.
LIU P,LIU H M,WANG L C,etal.Floral syndrome and breeding system ofCalotropisgigantean[J].ActaHorticulturaeSinica,2015,42(10):2 002-2 014.
[12]DAVIS G L.Systematic embryology of the angiosperms[M].New York,1966.
[13]胡适宜.被子植物胚胎学[M].北京:人民教育出版社,1982:30-72.
[14]ERDTMAN G.An introduction to pollen analysis[J].NabuPress,1943.
[15]黄衡宇,马绍宾,李鹂.八角莲小孢子发生和雄配子形成[J].植物研究,2001,21(4):561-566.
HUANG H Y,MA S B,LI L.The genesis of microspore and the formation of male gametophyte inDysosmaversipellis(Hance) M.Cheng[J].BulletinofBotanicalResearch,2001,21(4):561-566.
[16]王定康,孙桂芳,翟书华,等.青阳参小孢子发生和雄配子体发育[J].植物研究,2011,31(4):392-396.
WANG D K,SUN G F,etal.Microsporogenesis and male gametophyte development ofCynanchunotophyllumSchneid[J].BulletinofBotanicalResearch,2011,31(4):392-396.
[17]赵兴峰,孙卫邦,杨华斌,等.极度濒危植物西畴含笑的大小孢子发生及雌雄配子体发育[J].云南植物研究,2008,30(5):549-556.
ZHAO X F,SUN W B,YANG H B,etal.Mega- and microsporogenesis and development of female and male gametophytes ofMicheliacoriacea(Magnoliaceae),a globally criticical endangered plant in south-east Yunnan of China[J].ActaBotanicaYunnanica,2008,30(5):549-556.
[18]樊莉丽,彭方仁,周琦,等.楸树大小孢子发生与雌雄配子体发育的研究[J].西北植物学报,2011,31(3):431-438.
FAN L L,PENG F R ,ZHOU Q,etal.Sporogenesis and gametogenesis ofCatalpabungei(Bignoniaceae)[J].ActaBot.Boreal.-Occident.Sin.,2011,31(3):431-438.
[19]廖菊够,康洪梅,代瑾然,等.烟草大小孢子发生和雌雄配子体发育研究[J].西北植物学报,2013,33(1):11-16.
LIAO J G,KANG H M,DAI J R,etal.Megasporogensis,microsporogensis and development of female and male gametophyte ofNicotianadebneyi[J].ActaBot.Boreal.-Occident.Sin.,2013,33(1):11-16.
[20]邱璐.金花菜的小孢子发生及雄配子体形成过程中几种物质的细胞化学定位[J].云南农业大学学报,1993,8(1):57-62.
QIU L.Cytochemical localization of several substance of microsporogenesis and the formative process of male gametophyte ofCamelliachrysanthaTuyama[J].JournalofYunnanAgriculturalUniversity,1993,8(1):57-62.
[21]许小连.濒危植物羊角槭生殖生物学的初步研究[D].浙江临安:浙江农林大学,2012.
[22]PACICI E,FRANCHI G G.Pollen dispersal unit,gynoecium and pollination[M]//OWENS S J,RUDALL P I.Reproductive Biology.Kew:Royal Botanic Gardens,1998.
[23]PACICI E,FRANCHI G G.Types of pollen dispersal units and pollen competition[M]//CLEMENT C,PACINI E,AUDRAN J C,Anther and Pollen:from Biotechnology.Berlin:Springer-Verlag,1999.
[24]张大勇.植物生活史进化与繁殖生态学[M].北京:科学出版社,2004.
[25]BISWAS I.Embryological studies inDaemiaextensaBr.[J].IndianBot.Soc.,1957,36:207-222.
[26]SADEQ M A,PATHAK M R,SALIH A A,etal.Somatic embryogenesis and organogenesis for regeneration of endangered multipurpose desert plantLeptadeniapyrotechnicaforsk.Decne in the kingdom of Bahrain[J].AmericanJournalofPlantSciences,2014,15(5):2 342-2 353.
[27]FRYE T C.A morphological study of certain Asclepiadaceae[J].BotanicalGazette,1902,34:389-413.
[28]SABEt Y S.Development of the embryo-sac inCalotropisprocerawith especial reference to endosperm formation[J].AnnalsofBotany,1931,(3):503-518.
[29]王定康,孙桂芳,翟书华,等.青阳参的大孢子发生与雌配子体发育[J].植物研究,2011,31(3):265-270.
WANG D K,SUN G F,ZHAI S H,etal.Megasporogenesis and development of female gametophyte inCynanchunotophyllumSchneid[J].BulletinofBotanicalResearch,2011,31(3):265-270.
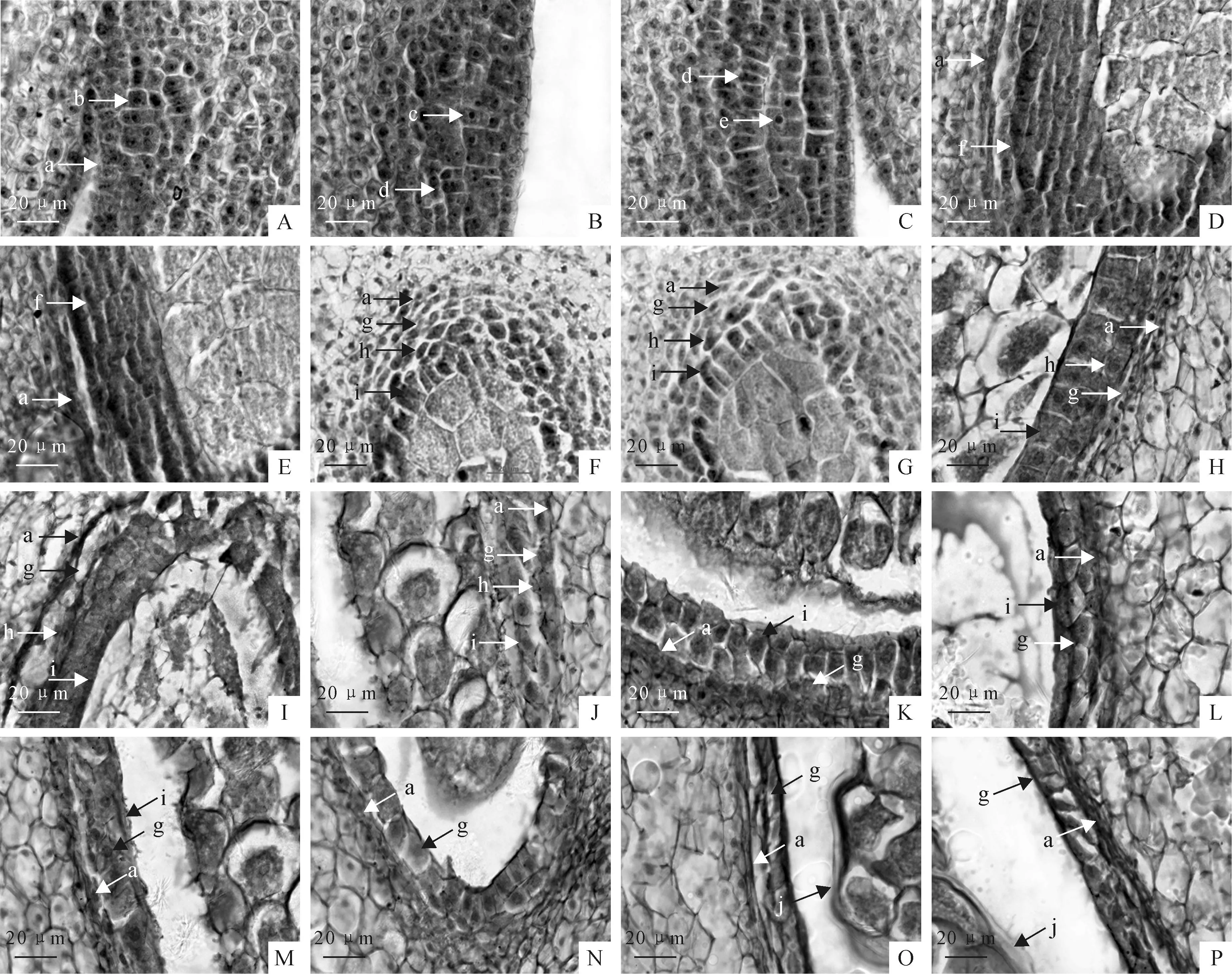
图版 Ⅰ牛角瓜花药壁的发育
a.表皮细胞;b.孢原细胞;c.初生造孢细胞;d.初生壁细胞;e.造孢细胞;f.次生壁细胞;g.药室内壁;h.中层;i.绒毡层;j.包裹花粉块的膜
A.孢原细胞平周分裂,向外形成初生壁细胞;B、C.初生壁细胞平周分裂,形成两层次生壁细胞;D、E.两层次生壁细胞进行多次垂周分裂;F、G.小孢子母细胞时期,形成四层药壁;H、I.小孢子母细胞减数分裂期,中层趋于解体;J.小孢子收缩期,中层仅留残余;K、L.单核小孢子靠边期,绒毡层少量解体;M、N.二核花粉粒时期,绒毡层基本上解体完成,仅剩余少量残留,开始形成包围花粉块的膜;O、P.成熟花粉粒时期,绒毡层完全解体,形成包围花粉块的膜。
Plate ⅠThe development of anther wall ofC.gigantea
a.Apidermis cells;b.Archesporial cell;c.Primary sporogenous cell;d.Primary parietal cells;e.Sporogenous cell;f.Secondary wall cells;g.Endothecium cells;h.Middle layer cells;i.Tapetum cells;j.Note the thick tapetal secretion enclosing the pollen mass
Fig.A.Archesporial cell periclinal division,forming primary parietal cells from the outer;Fig.B,C.Primary parietal cells anticlinal division,forming two layers of secondary wall;Fig.D,E.The stage of two secondary walls,the inner layer anticlinal division more than once;Fig.F,G.The stage of microspore mother cell,formed four layers of anther wall;Fig.H,I.The meiosis stage of microspore mother cell,middle layer going to degenerate;Fig.J.The stage of contraction of microspore,middle layer disappear;Fig.K,L.Microspore with the nucleus located aside,a small amount of tapetum cells degenerated;Fig.M,N.Two nuclear pollen stage,the tapetum disintegrated basically completed,only a small amount of surplus residue,began to form surrounded the pollinia membrane;Fig.O,P.Mature pollen grains,surrounding by uninucleate tapetal cells,note the thick tapetal secretion enclosing the pollen mass.
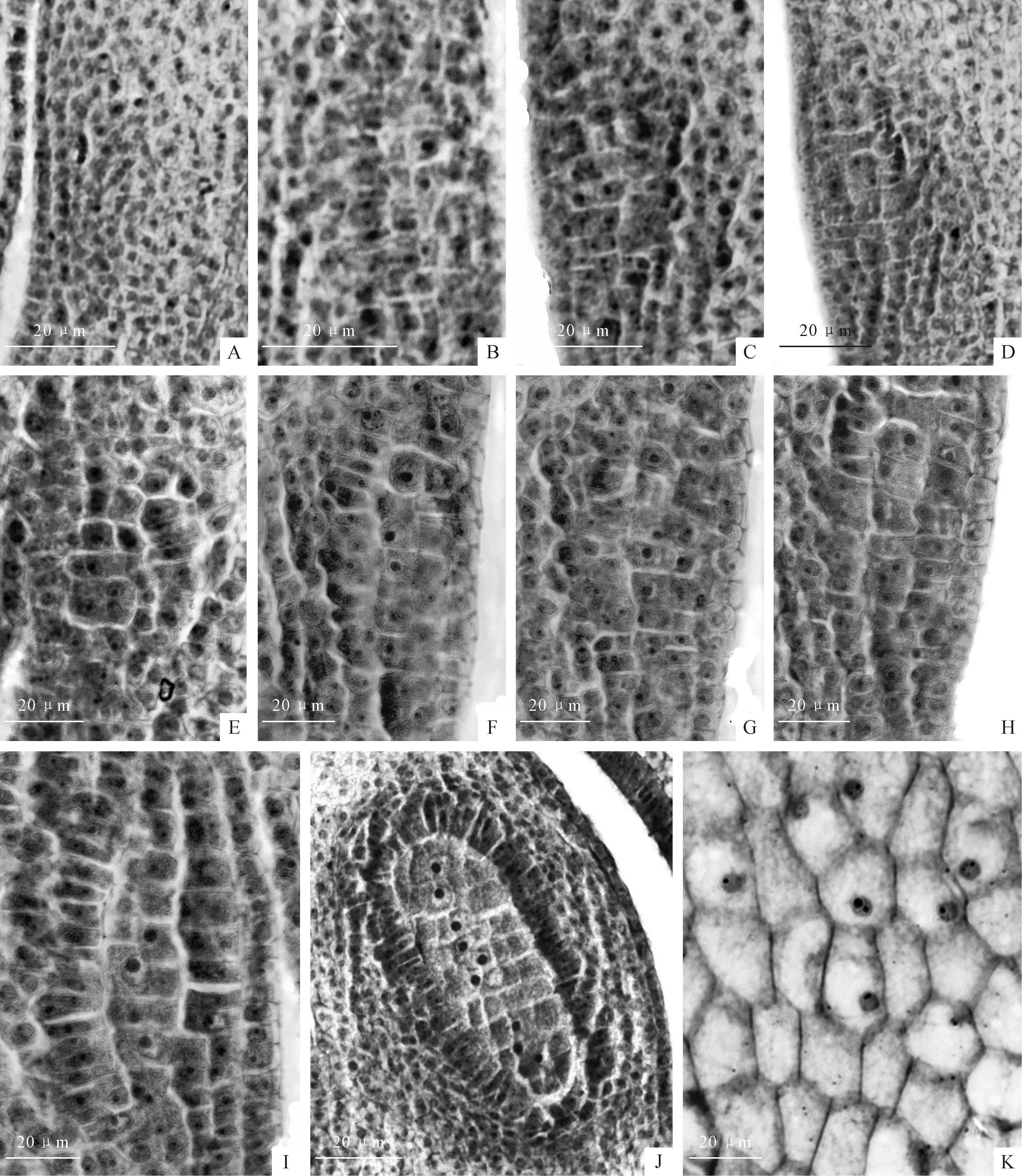
图版 Ⅱ牛角瓜小孢子的发育
A.未分化的花药;B.孢原细胞;C.孢原细胞平周分裂;D.早期的初生造孢细胞;E.初生造孢细胞;F.造孢细胞;G、H.初生造孢细胞直接行使小孢子母细胞功能;I.早期的小孢子母细胞;J、K.小孢子母细胞。
Plate ⅡThe microsporogenesis ofC.gigantea
Fig.A.The undifferentiated young anther;Fig.B.Archesporial cells;Fig.C.Archesporial cells anticlinal division;Fig.D.The early stage of primary sporogenous cells;Fig.E.Primary sporogenous cells;Fig.F.Primary sporogenous cells;Fig.G,H.Primary sporogenous cells directly exercise their microspore mother cells function;Fig.I.Early microspore mother cells;Fig.J,K.Microspore mother cells.
图版 Ⅲ牛角瓜雄配子体的发育Ⅰ
A.减数分裂前期Ⅰ细线期;B、C.减数分裂前期Ⅰ偶线期;D.减数分裂前期Ⅰ双线期;E、F.减数分裂前期Ⅰ终变期;G、H.减数分裂中期Ⅰ;I、J.减数分裂中后期Ⅰ;K、L.减数分裂后期Ⅰ
Plate ⅢThe development of male gametophyte Ⅰ ofC.gigantea
Fig.A.Leptotene stage of meiotic prophase Ⅰ;Fig.B,C.Pachytene stage of meiotic prophase Ⅰ;Fig.D.Diplotene stage of meiotic prophase Ⅰ;Fig.E,F.Diakinesis stage of meiotic prophase Ⅰ;Fig.G,H.Metaphase of meiosis Ⅰ;Fig.I,J.After anaphase of meiosis Ⅰ;Fig.K,L.Anaphase of meiosis Ⅰ.

图版 Ⅳ牛角瓜雄配子体的发育Ⅱ
A、B.减数分裂末期Ⅰ;C.二分体形成;D.减数分裂后期Ⅱ;E.四分体时期;F.收缩期小孢子;G.中央期小孢子;H.靠边期小孢子;I.靠边期小孢子核分裂;J.二核花粉粒;K.生殖细胞分裂;L.3-细胞型成熟花粉粒,具1营养核和2个精子。
Plate ⅣThe development of male gametophyte Ⅱ ofC.gigantea
Fig.A,B.Telophase of meiosis Ⅰ;Fig.C.Formed dyad;Fig.D.Anaphase of meiosis Ⅱ;Fig.E.Formed tetrads;Fig.F.The contraction microspore;Fig.G.The middle microspore;Fig.H.Microspore with the nucleus located aside;Fig.I.The mitosis of the microspore;Fig.J.Binuclear pollen;Fig.K.The mitosis of germ cell;Fig.L.Mature pollen with one vegetative nucleus and two sperm.

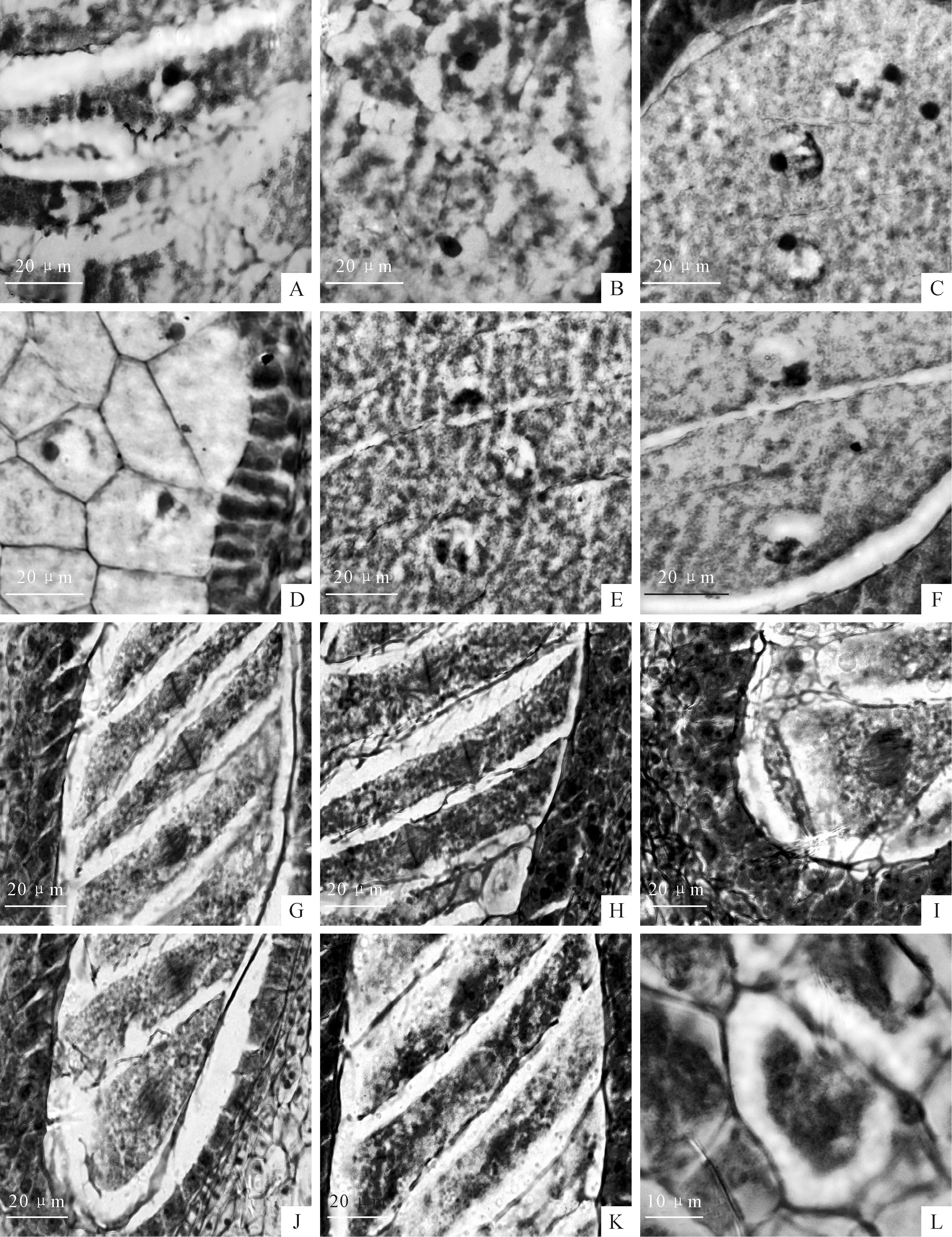
图版 Ⅴ牛角瓜大孢子的发育
A.心皮原基;B.胎座表皮下层细胞产生突起;C.胚珠原基;D.珠心(箭头指);E.大孢子母细胞(箭头指);F.大孢子母细胞第一次减数分裂后期(箭头指);G.二分体(箭头指);H.第二次减数分裂(箭头指);I.线形四分体(箭头指);J、K.四分体的近珠孔端3个细胞退化,合点端大孢子发育为功能大孢子(箭头指);L.单核胚囊(箭头指)。
Plate ⅤThe megasporogenesis ofC.gigantea
Fig.A.Carpellary primordia;Fig.B.Placentation the epidermal layer of cells and neurite outgrowth;Fig.C.Ovule primordium;Fig.D.Nucellus (arrow);Fig.E.Megasporocyte(arrow);Fig.F.Megaspore mother cell meiosis period for the first time (arrow);Fig.G.The megaspore dyad (arrow);Fig.H.Megaspore mother cell meiosis period for the second time (arrow);Fig.I.Four megaspores forming one liner tetrad (arrow);Fig.J,K.The chalazal megaspore was functional,while the other three megaspores in micropyle degenerated (arrow);Fig.L.The uninucleate embryo sac (arrow).

图版 Ⅵ牛角瓜雌配子体发育
A.单核胚囊(箭头指);B.单核胚囊第一次有丝分裂(箭头指);C~E.2核胚囊(箭头指);F、G.4核胚囊(箭头指);H.8核胚囊中,两端各有1核移向胚囊中间,形成极核(箭头指);I、J.连续切片示8核胚囊,上端为合点端,呈“品”字形的反足细胞;中间为2个极核所在的中央细胞;下端为珠孔端,中间一个为卵细胞,其他2个为助细胞(箭头指);8核胚囊内合点端的反足细胞开始退化(上端箭头指);K.2个助细胞(箭头指);L.3个呈“品”字形的反足细胞(箭头指)。
Plate ⅥThe development of female gametophyte ofC.gigantea
Fig.A.The uninucleate embryo sac (arrow);Fig.B.The uninucleate embryo sac mitotic division for the first time (arrow);Fig.C-E.Two-nucleus embryo sac (arrow);Fig.F,G.Four-nucleus embryo sac (arrow);Fig.H.Eight-nucleus embryo sac,both ends each have 1 nuclear move to embryo sac,form a polar nuclei (arrow);Fig.I,J.The mature embryo sac;Eight-nucleus embryo sac chalaza end of the antipodal cells began to degenerate (the upper arrow);Fig.K.Two synergid cells (arrow);Fig.L.The antipodal cells in mature embryo sac (arrow).
(编辑:潘新社)
Study on the Megasporogenesis,Microsporogenesis and Development of Female and Male Gametophytes ofCalotropisgiganteaL.
LIU Peng,ZHANG Taikui,LI Lu,LIU Ting,WANG Lianchun,LUO Mingcan,LIU Huimin*
(College of Forestry,Southwest Forestry University,Kunming 650224,China)
Abstract:To observe megasporogenesis,microsporogenesis and development of female and male gametophytes,we studied the fresh flowers of Calotropis gigantea using paraffin wax.The results were as follows:(1)Archesporial cell of C.gigantea microspore differentiated from two areas of subcutaneous tissue.The mature anther wall was composed of epidermis,endothecium (1 layer) and middle layer (2 layers),glandular tapetum (1 layer) and its development model belonged to basic type.The glandular tapetum was a secretory structure.Cytokinesis of microspore mother cells was continuously split twice.Three cells were observed in the mature pollen grain:2 small sperm cells and one large vegetative cell.(2)C.gigantea had two carpels in pistil and 1 locule in ovary.The ovule was anatropous,marginal placentation,unitegmic and tenuinucellus.The megaspore tetrads were linear and the chalazal megaspore was functional.The mature embryo sacs had seven cells with eight nucleis,and belonged to polygonum type sac.(3)C.gigantea pistil developed later than stamen,and the female and stamen developed synchronized before buds open.It was considered that the barrenness of C.gigantea was not due to male or female sterility,although some proportional abortion exists in the development of pistil and stamen.Maybe it was related to pollination fertilization.
Key words:Calotropis gigantea L.;mega- and microsporogenesis;female and male gametophyte
中图分类号:Q944.58
文献标志码:A
作者简介:刘鹏(1989-),男,在读硕士研究生,主要从事经济林栽培研究。E-mail:18388115168@163.com*通信作者:刘惠民,博士,教授,主要从事森林培育的教学与研究。E-mail:hmliu@swfu.edu.cn
基金项目:林业公益性行业科研专项(201304810)
收稿日期:2015-10-09;修改稿收到日期:2015-12-14
文章编号:1000-4025(2016)01-0048-11
doi:10.7606/j.issn.1000-4025.2016.01.0048
