基于记忆的注意捕获和注意抑制效应:ERP证据*
2016-02-02胡艳梅
胡艳梅 张 明
(1东北师范大学教育学部心理学院, 长春 130024) (2苏州大学教育学院心理学系, 苏州 215000)
1 引言
Desimone和Duncan (1995)提出的偏向竞争模型(Bias Competition Model)首次关注了客体工作记忆在注意捕获中的作用。视野内的各种刺激总是在互相竞争以获得注意。目标刺激凸显时, 会自动获取注意。目标刺激不凸显时, 为了将之与干扰刺激区分,需要把目标特征保持在工作记忆中。Desimone和Duncan(1995)将目标特征称为“注意模板(attentional template)”。注意模板是工作记忆的表征, 工作记忆内容总是会获取注意。
许多研究结果都支持了偏向竞争模型观点(Mannan, Kennard, Potter, Pan, & Soto, 2010; Olivers,Peters, Houtkamp, & Roelfsema, 2011; Pan, Xu, &Soto, 2009; Soto, Humphreys, & Heinke, 2006; 潘毅,2010; 张豹, 黄赛, 祁禄, 2013; 张豹, 黄赛, 侯秋霞, 2014)。行为证据方面, Downing (2000)发现当记忆刺激与目标刺激先后在同一位置上出现时, 被试对目标特征的辨别速度更快。Olivers, Meijer和Theeuwes (2006)利用视觉搜索任务探索工作记忆内容对注意的影响。要求被试在记忆一个视觉刺激的同时完成一项搜索任务。结果发现, 即便当工作记忆内容仅与搜索分心物匹配时, 记忆匹配刺激仍能捕获注意。脑神经研究方面, Kumar, Soto和Humphreys (2009)采用类似的范式, 在搜索任务中呈现了3个分心物和1个目标(分列视野两侧; 每侧视野2个项目)。控制条件下, 所有搜索项目均与记忆项目不同; Invalid条件下将有一个搜索分心物与记忆项目匹配。记忆匹配刺激可能出现在目标同侧,也可能出现在目标对侧。结果发现, 目标刺激诱发了N2pc成分。更重要的是, 与控制条件相比, 当记忆匹配刺激与目标刺激分列视野两侧时, 目标刺激所诱发的N2pc波幅变小了。N2pc成分与注意选择过程密切相关。视野某侧刺激捕获注意时, 将在其对侧脑区诱发比同侧脑区更大的负电位, 即 N2pc成分(Eimer & Kiss, 2007; Luck & Hillyard, 1994;Woodman, 2010; 姚树霞, 杨东, 齐森青, 雷燕,Ding, 2012)。因此, Kumar等(2009)的发现表明对目标刺激的注意受到了记忆匹配刺激的干扰。
尽管偏向竞争模型的观点得到了大量实验证据的支持, 但在工作记忆内容是否一定能捕获注意这一问题上已有研究仍存在争议。其中, 认知控制观点强调工作记忆内容对注意的引导过程是灵活可控的(Woodman & Luck, 2007; Hu, Xu, & Hitch,2011; 张明, 王爱君, 2012; 张豹, 黄赛, 2013)。与工作记忆内容匹配的刺激既可以捕获注意(后称为“基于记忆的注意捕获效应”), 也可以受到抑制(后称为“基于记忆的注意抑制效应”)。
一方面, 任务需求可以决定记忆内容会否成为注意内容。Woodman和Luck (2007)首先要求被试记忆若干视觉刺激, 然后完成一项搜索任务, 最后进行记忆测试。前 4个实验中, 匹配条件下, 记忆项目仅与搜索分心物一致而不同于搜索目标。结果发现, 匹配条件下搜索速度快于控制条件。这表明记忆匹配分心物受到了抑制, 从而促进了搜索任务表现。而实验5中, 匹配条件下记忆项目既可能与分心物一致, 也可能与搜索目标一致(该情况出现概率仅为 16.67%)。结果发现匹配条件下搜索速度更慢。这表明, 一旦记忆项目可能成为搜索目标,个体就会主动优先注意记忆项目, 从而影响了搜索任务表现。因此, Woodman和Luck (2007)指出, 工作记忆内容不仅可以是“注意模板” (Desimone &Duncan, 1995), 也可以是“抑制模板(reject temple)”。
另一方面, 认知控制调节的有效性受到许多因素的影响, 如认知控制动机水平和准备时间等(Carlisle & Woodman, 2011a; Han & Kim, 2009;胡艳梅, 张明, 徐展, 李毕琴, 2013)。时程方面, Han和Kim (2009)发现基于记忆的注意抑制过程只出现在信息加工速度较慢的复杂搜索任务中。简单搜索任务中, 记忆匹配刺激则捕获了注意。Hu等(2011)也发现基于记忆的注意抑制过程需要一定准备时间才能发挥作用。记忆项目呈现时间较长时, 记忆匹配刺激可以受到抑制。记忆项目呈现时间较短时,被试没有充足的时间进行认知控制准备, 难以有效抑制记忆匹配刺激。
虽然上述两项研究均表明, 基于记忆的注意抑制需要耗费一定时间, 但这并不意味着该效应的时程是固定不变的。胡艳梅等(2013)发现, 当个体进行认知控制的动机水平足够高时, 对记忆匹配刺激的抑制过程可以在一定程度上提前。一方面, 他们通过改变匹配试次在实验中出现的概率(20%/50%/80%)来操作被试的认知控制动机水平。匹配试次出现概率越高, 被试对记忆匹配刺激进行抑制的动机水平也越高。另一方面, 他们利用反应时分布等分法, 将被试的反应时数据从快到慢划分出 10个十分位点。取第一个十分位点的数据计算被试在快速加工过程中的表现。结果发现:快速加工过程中, 记忆匹配刺激在20%和50%条件下捕获了注意; 而在 80%条件下受到了抑制。可见, 当认知控制动机水平很高时, 对记忆匹配刺激的抑制过程被提前了。胡艳梅等(2013)的研究提示我们基于记忆的注意抑制效应不仅能发生在较晚的信息加工阶段, 也可能出现在较早的信息加工阶段。但鉴于行为研究方法在时间精度上的局限性, 我们并不能断定在加工早期记忆匹配刺激是否真的受到了抑制。胡艳梅等(2013)的研究中, 即便是第一个十分位点上的反应时数据, 其平均值也超过了500 ms。而认知神经研究指出, 注意转移过程在刺激呈现后200 ms左右就可以发生; 数据上反映为N2pc成分的出现(Luck & Hillyard, 1994; Woodman & Luck,2003)。Carlisle和 Woodman (2011b)利用 ERP技术探索加工早期工作记忆内容是否会捕获注意。结果发现, 记忆匹配刺激不但没有在加工早期(刺激呈现后200~300 ms左右)诱发N2pc成分, 反而在加工后期(300~500 ms左右)诱发了与抑制过程相关的差异正波。该差异正波表现为在记忆匹配刺激的对侧脑区出现了比同侧脑区更大的正电位。虽然Carlisle和Woodman (2011b)的发现表明, 工作记忆内容在加工早期没有捕获注意; 但却没有证据支持它们能迅速地受到抑制。
基于已有研究发现, 我们认为工作记忆对注意的引导作用可能分为两个阶段:(1)加工早期(200~300 ms左右)注意首次发生转移的时段; (2)加工后期(300 ms以后)认知控制发挥主导作用的时段。当注意指向工作记忆内容时, 该引导作用表现为“基于记忆的注意捕获”效应; 当注意偏离工作记忆内容时, 该引导作用表现“为基于记忆的注意抑制”效应。虽然在加工后期, 许多研究都一致发现了基于记忆的注意抑制效应。但在加工早期, 认知控制是否也能发挥作用还不甚明确。一方面, ERP研究结果还存在争议(Carlisle & Woodman, 2011b;Kumar et al., 2009); 另一方面, 行为研究结果虽然在时间精度上存在局限, 却提示我们认知控制作用的时程受到控制动机水平的影响(胡艳梅等, 2013)。
因此, 为了进一步确认信息加工早期基于记忆的注意抑制效应是否真的存在, 本研究利用 ERP技术, 在操纵认知控制动机水平的基础上, 考察基于记忆的注意引导过程的时程特点。实验采用工作记忆与视觉搜索双任务范式。搜索任务包括一个目标和一个分心物。匹配试次中, 搜索分心物颜色与记忆颜色相同。此外, 在记忆序列和搜索序列之间还插入了线索序列。ERP统计分析采用线索序列数据。一方面, 基于客体工作记忆的注意引导过程会出现在线索序列中。我们明确告知被试“记忆项目永远不同于搜索目标”以及“线索刺激的颜色和位置与搜索项目一一对应”。即匹配条件下, 线索序列和搜索序列中各有一个刺激与记忆项目匹配。如果客体工作记忆内容会引导注意, 那么与记忆项目匹配的线索刺激也将引导注意。并且, 与传统范式中搜索序列加工过程一致, 匹配条件下线索序列的注意转移也是基于客体的, 而与空间信息无关。首先,线索刺激位置(屏幕两侧)与记忆项目位置(屏幕中央)无关。其次, 线索刺激与记忆项目在客体特征上的相似性是影响注意转移的根本原因。因此, 线索序列中的注意转移过程反映了客体工作记忆内容对注意的引导作用。另一方面, 线索刺激不包含目标特征, 在该序列发生的注意引导过程将不会受到与搜索目标相关的知觉判断和按键反应的干扰。我们假设:如果加工早期记忆匹配刺激不能受到有效抑制, 那么无论认知控制动机水平如何, 记忆匹配刺激都将首先捕获注意(加工早期), 而后才可以被抑制(加工后期)。数据上表现为线索序列记忆匹配刺激首先诱发 N2pc成分, 随后诱发与抑制过程相关的差异正波。如果加工早期记忆匹配刺激可以受到有效抑制, 那么当认知控制动机水平很高时, 记忆匹配刺激将不再捕获注意, 而被迅速抑制。数据上表现为线索序列记忆匹配刺激直接诱发与抑制过程相关的差异正波。
2 方法
2.1 被试
15名大学生(5男, 10女, 年龄在19~25岁之间,M
= 21.13岁,SD
= 1.88)自愿报名参加本实验。每个被试均分4次完成实验, 每次实验持续约3小时(包括实验准备时间和正式实验时间)。所有被试视力或矫正视力正常, 辨色能力正常。2.2 实验材料
记忆项目为一个彩色圆形(直径 1.5°), 颜色包括红、黄、蓝、绿四种。线索序列包括两个彩色的空心方框(视角1.5°× 1.5°), 分别位于注视点左右两侧, 距离屏幕中心点3°。线索刺激的颜色和位置与随后出现的搜索项目一一对应。即如果被试在线索序列中看到一个位于屏幕左侧的红色方框和一个位于屏幕右侧的蓝色方框。则在随后的搜索序列中,他们也将在屏幕左侧看到一个红色图形, 在屏幕右侧看到一个蓝色图形。搜索序列包括两个颜色不同的圆形(直径 1.5°), 其中每个圆形内各嵌有一个黑色开口方框(方框视角 0.5°× 0.5°, 开口视角 0.12°,方框线条宽0.08°)。如果黑色方框朝垂直方向开口(即向上或者向下), 则该项目为搜索目标; 如果方框朝水平方向开口(即向左或向右), 则该项目为搜索分心物。搜索项目的可选颜色也包括红、黄、蓝、绿四种。记忆探测项目与记忆项目一致, 为一个彩色圆形。
2.3 实验设计
实验为3(概率条件:20%/50%/80%)×2(试次类型:匹配试次/控制试次)被试内设计。20%、50%和 80%条件下, 匹配试次数量分别占试次总量的1/5、1/2和 4/5。匹配试次中, 记忆颜色与搜索分心物相同, 但不同于搜索目标。控制试次中, 搜索目标和分心物的颜色均与记忆颜色不同。行为指标包括搜索反应时、搜索正确率和记忆正确率。ERP指标主要为:(1) N2pc成分; (2) N2pc结束后出现的与抑制过程相关的差异正波(后统一简称“抑制性差异正波”)。
2.4 实验程序
实验流程如图1所示。首先向被试呈现一个注视点(“+”) 500 ms。然后呈现记忆项目, 持续 500 ms。要求被试记住记忆项目的颜色。随后, 间隔500 ms,向被试呈现线索序列, 持续时间为500 ms。线索序列出现时, 仅要求被试注视线索刺激, 而不做任何按键反应。线索序列消失后间隔250 ms, 向被试呈现搜索序列。持续时间最长为 1500 ms, 被试按键反应后立即消失。要求被试迅速找出搜索目标, 并指明其中黑色方框的开口方向, 向上开口按字母键“A”, 向下开口按字母键“Z”。如果 1500 ms后被试仍未能反应, 则记为错误反应。搜索序列结束后,间隔500 ms, 再呈现记忆探测序列。记忆探测项目最长持续时间为 2000 ms, 被试按键反应后随即消失。要求被试判断记忆探测项目的颜色是否与记忆项目颜色一致, 一致按数字键“1”, 不一致按数字键“2”。每两个试次间间隔800~1000 ms。搜索任务反应要求又快又准, 记忆任务反应要求尽量准确。所有被试均被明确告知“记忆项目永远不同于搜索目标” 以及“线索刺激的颜色和位置与搜索项目一一对应”。
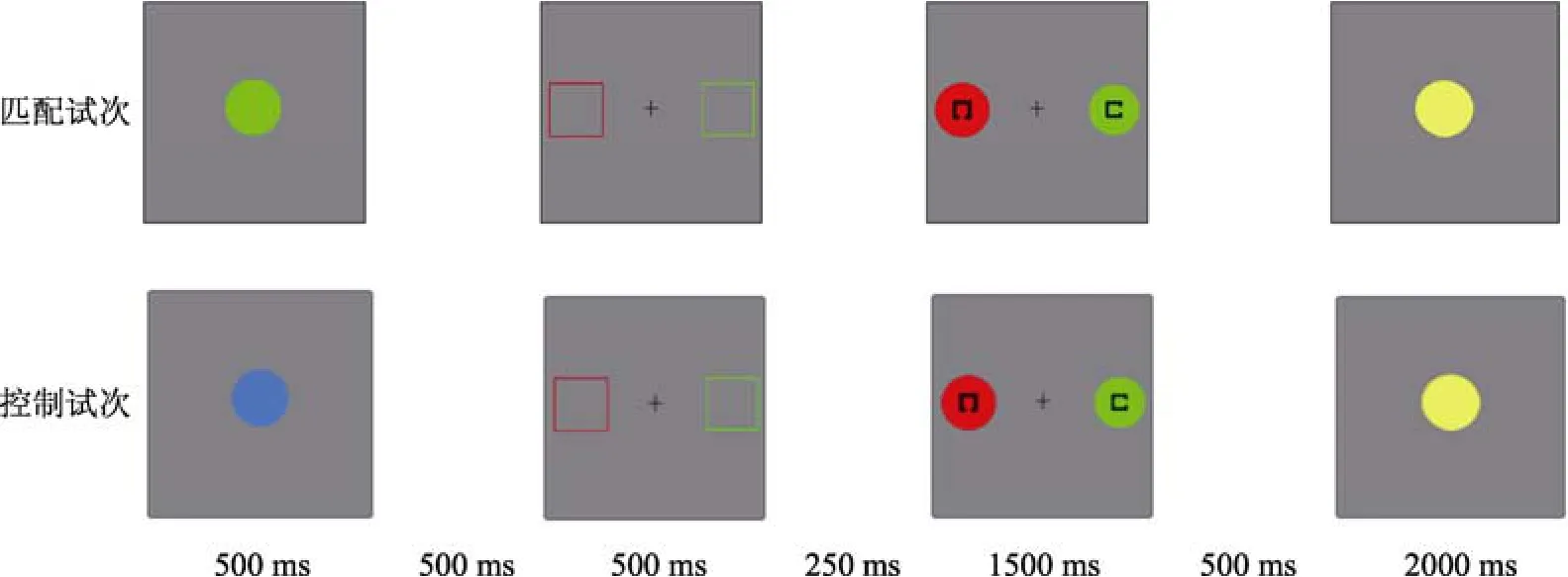
图1 实验流程
每48个试次为一个block。每个block开始前被试都将看到一个 4位数, 要求被试在整个 block中, 以每秒 1~2次的速度不断默默复述该数字(不出声)。每个block结束后, 主试将询问被试刚才所要求复述的数字是多少。不出声的发音抑制任务不涉及嘴唇运动, 因此不影响 ERP数据收集(Carlisle& Woodman, 2011b)。同时我们还告诉被试, 主试可以通过脑电信号监视他们复述数字的情况, 以进一步确保被试按要求进行发音抑制任务。
Woodman (2010)指出当以N2pc成分为指标时,单个条件叠加次数在250次左右或以上为宜。因此,为了收集到足够的匹配试次数据, 以供统计分析(线索序列中)由记忆匹配刺激诱发的 N2pc成分,20%条件下被试需完成 1440个试次。该条件分两次完成, 每次完成 720个试次(其中匹配试次 144个)。50%和80%条件下, 被试各需完成720个试次。这两种条件也分两次完成, 每次完成一种条件。因此, 被试总共将分 4次完成实验, 每两次实验间至少间隔一天。三种概率条件的顺序在被试间平衡。
2.5 数据统计方法
行为数据。
以搜索反应时, 搜索正确率和记忆正确率为指标, 分别对三种因变量进行 3(概率条件)×2(试次类型)重复测量方差分析。ERP数据记录。
脑电数据由Neuroscan ERP工作站采集, 用64导电极帽(根据国际10-20系统扩展)记录脑电信号。以左侧乳突为参考电极, 前额接地, 在双眼外侧约1.5 cm处安置电极记录水平眼电,左眼上下安置电极记录垂直眼电。采用 AC采集,滤波带通为 0.05~100 Hz, 采样频率为 500 Hz/导,头皮与电极间阻抗小于5 kΩ。对连续记录的EEG数据进行离线分析时, 将左侧乳突参考转换为双侧乳突参考, 并进行30 Hz/24 bt的低通滤波。同时排除波幅大于±60 μV的眼电或±80 μV的其他电极伪迹信号。分析时程截取线索刺激呈现前100 ms至呈现后600 ms, 并以刺激呈现前100 ms作为基线。ERP数据分析。
本研究主要考察线索序列记忆匹配刺激同侧和对侧脑区的脑电激活差异。因此ERP统计分析仅涉及匹配试次。具体指标包括:①由记忆匹配刺激所诱发的 N2pc成分, 它反映了记忆匹配刺激对注意的捕获过程; ②N2pc结束后出现的抑制性差异正波, 它反映了对记忆匹配刺激的抑制过程。选取头皮后部两侧区域(P5/6, P7/8,PO5/6, PO7/8)激活进行分析。首先, 分别在 180~250 ms和250~ 320 ms时间窗口对记忆匹配刺激激活进行 3(概率条件)×2(脑区:记忆匹配刺激同侧/对侧)×4(电极对)重复测量方差分析。其次, 根据ERP波形分别在180~320 ms, 180~280 ms和180~250 ms时间窗口对20%、50%和80%条件下的N2pc平均波幅和50%面积潜伏期(将统计窗口内ERP波形面积分为1/2的时刻点)进行3(概率条件)×4(电极对)重复测量方差分析。最后, 在320~550 ms时间窗口对记忆匹配刺激激活进行 3×2×4重复测量方差分析, 以检验随后可能出现的抑制性差异正波。3 结果与分析
3.1 行为结果
行为实验结果见表1。对数据进行3×2重复测量方差分析。

表1 被试的平均任务表现(M ± SE)
搜索反应时方面, 概率条件主效应显著,F
(2, 28) =26.16,MSE
= 100753.18,p
< 0.001, η= 0.65。试次类型主效应显著,F
(1, 14) = 190.55,MSE
= 222939.74,p
< 0.001, η= 0.93。概率条件与试次类型交互作用显著,F
(2, 28) = 30.30,MSE
= 90817.45,p
< 0.001,η= 0.68。简单效应分析结果表明, 20%条件下, 试次类型差异不显著,t
< 1; 50%和80%条件下, 匹配试次搜索反应时均快于控制试次,t
(14) = 27.08和19.01,ps
< 0.001, 即搜索任务受到了促进。搜索正确率方面, 试次类型主效应显著,F
(1,14) = 19.03,MSE
= 0.001,p
= 0.001, η= 0.58。其余主效应和交互作用不显著。记忆正确率方面, 所有主效应和交互作用均不显著。3.2 ERP结果
线索序列记忆匹配刺激两侧脑区平均激活水平如图2所示。
(1) N2pc成分
分别在180~250 ms和250~320 ms时间窗口,对头皮后部两侧脑区(P5/6, P7/8, PO5/6, PO7/8)激活进行 3(概率条件)×2(脑区)×4(电极对)重复测量方差分析。
180~250 ms时间窗口内, 概率条件主效应不显著。脑区主效应显著,F
(1, 14) = 76.68,MSE
=166.28,p
< 0.001, η= 0.85。记忆匹配刺激对侧脑区比同侧脑区激活了更大的负电位。电极对主效应显著,F
(3, 42) = 4.43,MSE
= 24.46,p
< 0.05, η=0.24。概率条件与脑区的交互作用显著,F
(2, 28) =7.13,MSE
= 5.57,p
< 0.01, η= 0.33。概率条件与电极对的交互作用显著,F
(6, 84) = 4.83,MSE
= 4.653,p
< 0.01, η= 0.26。其余交互作用不显著。对概率条件与脑区的交互作用进行简单效应检验, 结果显示:20%、50%和 80%条件下, 记忆匹配刺激对侧脑区都比同侧脑区诱发了更大的负电位,t
(14) =4.88, 11.15和6.28,ps
< 0.001。250~320 ms时间窗口内, 概率条件主效应显著,F
(2, 28) = 9.59,MSE
= 173.28,p
< 0.01, η= 0.41。脑区主效应显著,F
(1, 14) = 5.11,MSE
= 9.18,p
<0.05, η= 0.27。记忆匹配刺激对侧脑区比同侧脑区激活了更大的正电位。电极对主效应显著,F
(3, 42) =13.06,MSE
= 54.74,p
< 0.001, η= 0.48。概率条件与脑区的交互作用显著,F
(2, 28) = 36.18,MSE
=34.52,p
< 0.001, η= 0.72。其余交互作用不显著。对概率条件与脑区的交互作用进行简单效应检验。结果显示, 20%条件下, 记忆匹配刺激对侧脑区电位更负,t
(14) = 7.45,p
< 0.001。50%条件下, 记忆匹配刺激两侧脑区激活水平差异不显著,t
(14) =1.11。80%条件下, 记忆匹配刺激对侧脑区电位更正,t
(14) = ‒5.55,p
< 0.001。从图2可见, 不同概率条件下 N2pc成分的持续时间差异较大。分别在180~320 ms, 180~280 ms和180~250 ms时间窗口内统计20%、50%和80%条件下 N2pc的平均波幅和 50%面积潜伏期, 并进行3(概率条件)×4(电极对)重复测量方差分析。N2pc波幅方面, 概率条件差异显著,F
(2, 28) = 10.11,MSE
= 10.11,p
= 0.001, η= 0.42。20%和 80%条件下 N2pc波幅均小于 50%条件,ps
< 0.01; 20%和80%条件之间差异不显著,p
> 0.05。电极对主效应和交互作用不显著。N2pc潜伏期方面, 概率条件主效应显著,F
(2, 28) = 24.86,MSE
= 8557.96,p
<0.001, η= 0.64。20%条件下N2pc潜伏期慢于50%和80%条件,p
< 0.05和p
< 0.001; 50%条件下N2pc潜伏期慢于80%条件,p
= 0.001。电极对主效应不显著。概率条件与电极对交互作用显著,F
(6, 84) =4.50,MSE
= 895.94,p
< 0.05, η= 0.24。(2)抑制性差异正波
在 320~550 ms时间窗口内, 对记忆匹配刺激激活的平均波幅进行3×2×4重复测量方差分析。结果显示, 概率条件主效应显著,F
(2, 28) = 6.12,MSE
= 306.60,p
< 0.05, η= 0.30。脑区主效应显著,F
(1, 14) = 62.97,MSE
= 281.76,p
< 0.001,ŋ
= 0.82。记忆匹配刺激对侧脑区激活了更大的正电位。电极对主效应显著,F
(3, 42) = 17.90,MSE
= 74.36,p
< 0.001, η= 0.56。概率条件与脑区的交互作用显著,F
(2, 28) = 34.20,MSE
= 90.11,p
< 0.001,ŋ
= 0.71。脑区与电极对交互作用显著,F
(3, 42) = 20.42,MSE
=26.22,p
< 0.001, η= 0.59。三阶交互作用显著,F
(6,84) = 10.28,MSE
= 8.70,p
< 0.001, η= 0.42。
图2 线索序列记忆匹配刺激两侧脑区平均激活水平差异
进一步分析各概率条件下记忆匹配刺激两侧脑区激活水平的差异。20%条件下, 记忆匹配刺激两侧脑区激活水平差异不显著,t
(14) = 1.07; 50%和80%条件下, 记忆匹配刺激对侧脑区比同侧脑区激活了更大的正电位,t
(14) = 8.98和7.22,ps
< 0.001。分别计算各概率条件下记忆匹配刺激两侧脑区激活水平的差值, 并进行两两比较。20%条件下该差值(M
= 0.22 μV)小于 50%条件(M
= 2.23 μV)和 80%条件(M
= 2.86 μV),ps
< 0.001。50%条件下该差值小于80%条件,p
< 0.05。4 讨论
总体上, 行为结果和ERP结果发现:(1)刺激呈现后 200~300 ms左右, 记忆匹配刺激总是会诱发N2pc成分, 但其平均波幅和潜伏期受到认知控制动机水平的调节; (2)刺激呈现后300 ms以后记忆匹配刺激会诱发与抑制过程相关的差异正波, 其平均波幅受到认知控制动机水平的调节。
4.1 基于记忆的注意引导过程包括两个阶段
本研究结果表明, 与客体工作记忆匹配的任务无关刺激并不能直接受到抑制。客体工作记忆内容对注意的引导过程包括两个阶段:(1)加工早期的注意捕获阶段(200~300 ms左右); (2)加工后期的认知控制阶段(300 ms以后)。
加工早期, 与工作记忆内容匹配的刺激总会捕获注意。ERP结果显示刺激出现后200~300 ms左右, 所有条件下记忆匹配刺激均诱发了N2pc成分。支持了记忆匹配刺激对注意的捕获效应(潘毅, 2010;张明, 王爱君, 李毕琴, 张阳, 2013; 张豹等, 2013)。该结果与 Kumar等(2009)的发现一致。但 Carlisle和Woodman (2011b)却发现记忆匹配刺激没有诱发N2pc成分。这与实验所采用的刺激参数有关。Olivers (2009)曾指出, 基于记忆的注意捕获效应受到刺激强度的影响。Carlisle和 Woodman (2011b)的研究中, 搜索刺激数量大(20个左右), 视角小(0.33°×0.33°), 导致记忆匹配刺激在视野中不够突出(刺激强度弱), 难以迅速吸引注意, 因而没有观察到显著的 N2pc成分。本研究中, 搜索项目数量小(2 个), 视角大(1.5°×1.5°), 记忆匹配刺激的强度足够吸引被试注意, 因而发现了显著的N2pc成分。此外, Kumar等(2009)所采用的刺激数量(4个)和大小(最刺激小视角为1.5°×1.5°)与本研究更接近。其结果也与本研究结果更一致, 表明记忆匹配刺激的出现会吸引注意。
加工后期, 记忆匹配刺激还可以受到抑制。ERP结果显示在刺激出现后300 ms左右以后, 记忆匹配刺激会诱发与抑制过程相关的差异正波。该结果与许多已有研究结果一致。如, Carlisle和Woodman (2011b)在刺激呈现后375 ms左右发现了与抑制过程相关的差异正波, 表明工作记忆内容受到了抑制。Kumar等(2009)也发现, Invalid条件下记忆匹配刺激先捕获了注意, 后受到了抑制。可见信息加工后期, 工作记忆内容对注意的引导过程是非常灵活的, 当被试发现记忆匹配刺激不是任务目标后, 便会立即抑制它们。同时, 该抑制过程受到认知控制动机水平的调节。首先, 认知控制动机水平的高低会影响抑制过程的有效性。20%条件下,对记忆匹配刺激进行抑制的难度大而收益小; 被试的认知控制动机水平很低, 难以克服记忆匹配刺激对注意的捕获作用。其次, 认知控制动机水平的高低还会影响抑制过程的效应量。80%条件比50%条件诱发了更大的抑制性差异正波。这表明, 随着认知控制动机水平的提高, 记忆匹配刺激受到了更大的抑制。
4.2 认知控制在信息加工早期的作用
虽然加工早期(200~300 ms左右)工作记忆内容总是会捕获注意, 但随着认知控制动机水平的提高,注意捕获的效应量会减小, 持续时间会变短。即加工早期基于记忆的注意捕获效应也可以受到认知控制作用的调节。
与中等水平的认知控制动机(50%条件)相比,当认知控制动机水平较高时(80%条件), 记忆匹配刺激诱发的N2pc成分更小。注意捕获量上的变化表明:虽然工作记忆内容对注意的捕获作用是不可逆的, 但却是可控的。即虽然注意向工作记忆内容转移的趋势不受自上而下加工的影响, 但注意转移后, 对相应刺激的加工程度却可以受到认知控制的调节。关于N2pc成分的研究也表明, N2pc不仅反映了注意转移过程, 也反映了注意转移后在注意位置上的信息加工过程(Kiss, van Velzen, & Eimer,2008)。此外还需注意, 20%条件下 N2pc波幅也小于50%条件。这可能与20%条件下记忆匹配刺激出现概率太小, 没有引起被试足够的注意有关。Carlisle和Woodman (2011a)发现, 即便在有意注意的情况下低概率条件中记忆匹配刺激对注意的捕获作用也更低。本研究中, 被试甚至没有主动注意记忆匹配刺激的倾向。因此, 20%条件下注意捕获效应较小也是合理的。此外, 该结果还排除了刺激新异性的干扰。意外而不经常出现的刺激更容易吸引注意(Squires, Squires, & Hillyard, 1975; Polich,2007)。因此, 如果刺激新异性影响了实验结果, 那么20%条件下记忆匹配刺激最为罕见, 也最能捕获注意。但本研究结果却发现20%条件下注意捕获效应更小。这表明, 记忆匹配刺激对注意的捕获作用与刺激新异性关系甚微, 而反映了客体工作记忆内容对注意选择的影响。
更重要的是, 随着认知控制动机水平的提高,N2pc潜伏期越来越短。一般而言, N2pc潜伏期与注意转移速度成正相关(Brisson, Robitaille, &Jolicoeur, 2007)。然而考虑到250~320 ms时间窗口内各概率条件下 N2pc波形, 本研究中潜伏期差异实际体现的是注意捕获持续时间差异, 而非简单的注意捕获速度差异。80%条件下, 记忆匹配刺激对注意的捕获仅持续了70 ms左右(180~250 ms); 50%条件下, 持续了100 ms左右(180~280 ms); 20%条件下, 持续了140 ms左右(180~320 ms)。注意捕获持续时间的变化表明:当认知控制动机水平提高时,认知控制更早地介入了基于记忆的注意引导过程,从而促使注意更快地脱离工作记忆内容。Theeuwes(2010)指出, 在客体特征识别前, 凸显刺激对注意的捕获不受自上而下加工调节; 而当客体特征得到识别后, 认知控制就可以发挥作用了。如果被试发现所注意的刺激是分心物而非目标, 那么就会使注意迅速脱离该刺激(自上而下加工)。本研究进一步指出, 注意脱离的速度与认知控制动机水平有关。认知控制动机水平很低时, 基于记忆的注意捕获过程几乎没有受到自上而下加工的调节, 将持续到捕获效应自然消失(20%条件)。而随着认知控制动机水平的不断提高, 自上而下调节作用的力度越来越大, 注意脱离记忆内容的速度越来越快, 因而基于记忆的注意捕获效应的持续时间就越来越短了(50%和80%条件)。
4.3 对偏向竞争理论的补充
本研究结果支持和补充了偏向竞争模型(Desimone & Duncan, 1995)。 首先, 记忆匹配刺激的注意捕获效应支持了偏向竞争模型对工作记忆内容和注意模板关系的假设, 即工作记忆内容即是注意模板。本研究采用ERP技术, 直观的揭示了无论认知控制动机水平如何变化, 在信息加工早期(200~300 ms左右)记忆匹配刺激都会捕获注意, 并诱发显著的N2pc成分。该结果有力地证实了“记忆的即是注意的”这一观点。但需要强调的是, 偏向竞争模型还假设注意模板即是目标特征。而本研究中,工作记忆内容虽然捕获了注意, 但却不同于搜索目标。这表明, 工作记忆内容虽然总是注意的内容,但却不一定符合目标特征。由此可见, 基于记忆的注意捕获效应在很大程度上是机械化的——由于目标与干扰刺激的区别仅在于是否保持在工作记忆中, 因此默认“工作记忆内容与目标特征一致”,并进一步将记忆内容当做注意模板, 从而使记忆匹配刺激捕获注意。这里, “工作记忆内容与目标特征一致”仅仅是一个自上而下的假设前提而非知觉判断的结果。因此实际加工过程中, 即便客体工作记忆内容不符合目标特征, 与之匹配的刺激也会捕获注意。
其次, 本研究对抑制性认知控制作用的探讨弥补了偏向竞争模型在灵活性上的缺陷。前已述及,偏向竞争模型假设工作记忆内容与目标特征一致,是注意模板。然而, 当工作记忆内容与任务无关时,如果仍然维持对记忆内容的持续注意就太机械了。因此, 在基于记忆的注意引导过程中必须考虑认知控制(主要指对任务无关刺激的抑制)的影响。一方面, 本研究结果表明基于记忆的注意捕获效应虽然不能被认知控制作用所消除, 但却可以受到认知控制动机水平的调节。另一方面, 本研究结果也支持了认知控制需要一定准备时间才能发挥作用。刺激出现后, 即便被试知道“记忆项目永远不同于搜索目标”, 也无法立即进行抑制性认知控制。但随着加工过程的推进, 认知控制会逐渐发挥主导作用。此外, Engle (2002)指出认知控制的效力受到个体差异的影响。虽然本研究重点关注被试认知控制动机水平随概率变化的一致性趋势, 但不同被试受概率变化影响的程度可能存在差异。后续研究可深入考察个体差异对基于记忆的注意引导作用的影响, 以进一步揭示客体工作记忆对注意选择的影响机制。
5 结论
(1)加工早期(200~300 ms左右), 与工作记忆内容匹配的刺激总是会捕获注意, 但其注意捕获量和持续时间受到认知控制动机水平的调节;
(2)加工后期(300 ms以后), 与工作记忆内容匹配的刺激可以受到有效的抑制, 该过程受到认知控制动机水平的调节。
Brisson, B., Robitaille, N., & Jolicoeur, P.(2007).Stimulus intensity affects the latency but not the amplitude of the N2pc.NeuroReport, 18,
1627−1630.Carlisle, N.B., & Woodman, G.F.(2011a).Automatic and strategic effects in the guidance of attention by working memory representations.Acta Psychologica, 137
(2), 217−225.Carlisle, N.B., & Woodman, G.F.(2011b).When memory is not enough: Electrophysiological evidence for goal-dependent use of working memory representations in Guiding visual attention.Journal of Cognitive Neuroscience, 23
(10),2650−2664.Desimone, R.& Duncan, J.(1995).Neural mechanisms of selective visual attention.Annual Review of Neuroscience,18
, 193−222.Downing, P.E.(2000).Interactions between visual working memory and selective attention.Psychological Science,11
(6), 467−473.Eimer, M., & Kiss, M.(2007).Attentional capture by task-irrelevant fearful faces is revealed by the N2pc component.Biological Psychology, 74
(1), 108−112.Engle, R.W.(2002).Working memory capacity as executive attention.Current Directions in Psychological Science, 11
,19−23.Han, S.W., & Kim, M.S.(2009).Do the contents of working memory capture attention? Yes, but cognitive control matters.Journal of Experimental Psychology: Human Perception and Performance, 35
(5), 1292−1302.Hu, Y.M., Xu, Z., & Hitch, G.J.(2011).Strategic and automatic effects of visual working memory on attention in visual search.Visual Cognition, 19
(6), 799−816.Hu, Y.M., Zhang, M., Xu, Z., & Li, B.Q.(2013).Guidance of working memory on attention: The effects of inhibition incentive.Acta Psychologica Sinica, 45
(2), 127−138.[胡艳梅, 张明, 徐展, 李毕琴.(2013).客体工作记忆对注意的导向作用: 抑制动机的影响.心理学报, 45
(2), 127−138.]Kiss, M., van Velzen, J., & Eimer, M.(2008).The N2pc component and its links to attention shifts and spatially selective visual processing.Psychophysiology, 45
(2), 240−249.Kumar, S., Soto, D., & Humphreys, G.W.(2009).Electrophysiological evidence for attentional guidance by the contents of working memory.European Journal of Neuroscience, 30
, 307−317.Luck, S.J.(2005).An Introduction to the Event-Related Potential Technique.
Cambridge, MA: A Bradford Book.Luck, S.J.& Hillyard, S.A.(1994).Spatial filtering during visual search: Evidence from human electrophysiology.Journal of Experimental Psychology: Human Perception and Performance, 20
, 1000–1014.Mannan, S.K., Kennard, C., Potter, D., Pan, Y., & Soto, D.(2010).Early oculomotor capture by new onsets driven by the contents of working memory.Vision Research, 50
, 1590−1597.Olivers, C.N.L., Meijer, F., & Theeuwes, J.(2006) Feature-based memory-driven attentional capture: Visual working memory content affects visual attention.Journal of Experimental Psychology: Human Perception and Performance, 32
,1243−1265.Olivers, C.N.L.(2009).What drives memory-driven attentional capture? The effects of memory type, display type, and search type.Journal of Experimental Psychology: Human Perception and Performance, 35
, 1275–1291.Olivers, C.N.L., Peters, J., Houtkamp, R., & Roelfsema, P.R.(2011).Different states in visual working memory: When it guides attention and when it does not.Trends in Cognitive Sciences, 15
(7), 327−334.Pan, Y.(2010).Content-based working memory-driven visual attention.Advances in Psychological Science, 18
(2), 210−219.[潘毅.(2010).基于工作记忆内容的视觉注意.心理科学进展, 18
(2), 210−219.]Pan, Y., Xu, B.H., & Soto, D.(2009).Dimension-based working memory-driven capture of visual selection.The Quarterly Journal of Experimental Psychology, 62
(6),1123−1131.Polich, J.(2007).Updating P300: An integrative theory of P3a and P3b.Clinical Neurophysiology, 118
(10), 2128−2148.Soto, D., Humphreys, G.W., & Heinke, D.(2006).Working memory can guide pop-out search.Vision Research, 46
,1010−1018.Squires, N.K., Squires, K.C., & Hillyard, S.A.(1975).Two varieties of long-latency positive waves evoked by unpredictable auditory stimuli in man.Electroencephalography and Clinical Neurophysiology, 38
, 387−401.Theeuwes, J.(2010).Top-down and bottom-up control of visual selection.Acta Psychologica, 135
(2), 77−99.Woodman, G.F.(2010).A brief introduction to the use of event-related potentials (ERPs) in studies of perception and attention.Attention, Perception, & Psychophysics, 72
(8),2031−2046.Woodman, G.F., & Luck, S.J.(2003).Serial deployment of attention during visual search.Journal of Experimental Psychology: Human Perception and Performance, 29
,121−138.Woodman, G.F., & Luck, S.J.(2007).Do the contents of visual working memory automatically influence attentional selection during visual search?Journal of Experimental Psychology: Human Perception and Performance, 33
,363−377.Yao, S.X., Yang, D., Qi, S.Q., Lei, Y., & Ding, C.(2012).Studies on the N2pc component in visual spatial attention.Advances in Psychological Science, 20
(3), 365−375.[姚树霞, 杨东, 齐森青, 雷燕, Ding, C.(2012).视觉空间注意研究中的N2pc成分述评.心理科学进展, 20
(3), 365−375.]Zhang, B., & Huang, S.(2013).Mechanisms underlying the attentional guidance form working memory representations.Advances in Psychological Science, 21
(9), 1578−1584.[张豹, 黄赛.(2013).工作记忆表征对视觉注意的引导机制.心理科学进展, 21
(9), 1578−1584.]Zhang, B., Huang, S., & Hou, Q.X.(2014).The priority of color in working-memory-driven ocular capture.Acta Psychologica Sinica, 46
(1), 17−26.[张豹, 黄赛, 侯秋霞.(2014).工作记忆表征捕获眼动中的颜色优先性.心理学报, 46
(1), 17−26.]Zhang, B., Huang, S., & Qi, L.(2013).Working memory representation does guide visual attention: Evidence from eye movements.Acta Psychologica Sinica, 45
(2), 139−148.[张豹, 黄赛, 祁禄.(2013).工作记忆表征引导视觉注意选择的眼动研究.心理学报, 45
(2), 139−148.]Zhang, M., & Wang, A.J.(2012).Working memory contentbased attentional capture and suppression in the visual search.Advances in Psychological Science, 20
(12), 1899−1907.[张明, 王爱君.(2012).视觉搜索中基于工作记忆内容的注意捕获与抑制.心理科学进展, 20
(12), 1899−1907.]Zhang, M., Wang, A.J., Li, B.Q., & Zhang, Y.(2013).Contentbased Working Memory-driven Visual Attention Under the Condition of Color Highlight.Journal of Psychological Science 2013, 36
(6), 1307−1311.[张明, 王爱君, 李毕琴, 张阳.(2013).颜色凸显条件下基于工作记忆内容的视觉注意.心理科学, 36
(3), 1307−1311.]