1~6秒时距认知分段性特征*
2016-02-01尹华站陈盈羽黄希庭
尹华站 李 丹 陈盈羽 黄希庭
(1重庆师范大学教育科学学院, 重庆沙坪坝 401331) (2西南大学心理学部, 重庆北碚 400715)
1 前言
自上世纪八九十年代以来, 研究者开始系统关注一个重要的科学问题:数秒以内时距认知是否具有分段性?对于不同时距, 人们加工机制和表征方式不同, 这称为时距认知的分段性(黄希庭, 李伯约,张志杰, 2003)。迄今为止, 围绕这一主题, 学术界涌现出两类假说:非分段性假说和分段性假说。前者主张不同长度的时距认知机制一致(Church,1984), 后者则主张不同长度时距认知的机制不一致,分界点涉及3~5 s、2~3 s、1 s、1/2 s及1/3 s等(Fraisse,1984; Musterberg, 1889; Lewis & Miall, 2003a; Michon,1985; Pöppel,1997)。近些年来, 研究者对1 s以内与1 s以上之间的加工机制差异探讨较多(Cellini, Fabbri,Martoni, Tonetti, & Natale, 2014; Cordes, & Meck,2014;Gupta, 2014; Hayashi, Kantele, Walsh, Carlson,& Kanai, 2014), 而对1 s以上时距认知的分段性问题的探讨方兴未艾。基于此, 本研究主要探讨1 s以上时距认知的分段性问题, 这不仅为两类假说提供可能新证据, 继而为彻底揭示1 s以上时距认知机制奠定初步基础, 而且也为人类合理运用时距认知规律, 指导生活实践提供理论参考。
分段性假说主要体现在Fraisse和Pöppel等的观点上。Fraisse指出, 时间知觉范围大约过了上述限度, 我们对持续时间的认知就不是对现实的复制,而只能对长时记忆进行重构, 这是时间估计或时间记忆(Fraisse, 1984)。时间知觉与“知觉到的现在”的内涵大致相同。Pöppel认为“知觉到的现在”的上限就是意识的限度, 在该限度内, 相继的事件信息可以被整合成为一个单元, 或一个“格式塔” (gestalt),在这个格式塔所决定的时间范围内, 意识才得以体现。意识的限度约为3 s钟, 不同人“知觉到的现在”是不同的, 有的人为2 s, 有的人可能是4 s (Pöppel, 1997)。
不同领域的研究均提供了1 s以上时距认知具有分段性的证据。在一系列涉及动作事件任务、语言加工任务、感觉运动同步任务、双可图识别任务及节奏感识别任务的研究中, 发现2~3 s可能是一个关键的时间节点(Schleidt, Eibl-Eibesfeldt, & Pöppel, 1987;Kien & Kemp, 1994; Mates, Müller, Radil, & Pöppel,1994; Gomez, Argandona, Solier, Angulo, & Vazquez,1995; Szelag, 1997; Szelag, von Steinbüchel, Reiser,Gilles de Langen, & Pöppel, 1996; Szelag, Kowalska,Rymarczyk, & Pöppel, 1998)。当然, 一系列采用心理物理法的时间信息加工研究也提供了类似的证据。研究发现, 500 ms至2 s标准时距的韦伯系数是相当稳定的(Kristofferson, 1976), 对于人类而言, 估计200 ms至2 s范围内的时距, 韦伯系数大约是标准时距的10% (Hirsh, Monahan, Grant, & Singh, 1990)。同时在另一些研究中发现标准时距长于2~3 s时,韦伯系数会显著地增加, 这可能意味着人类对估计时距的敏锐性在2 s以上会发生改变(Woodrow, 1930)。另外, 脑损伤研究(Kagerer, Wittmann, Szelag, & von Steinbüchel, 2002)、药理学研究(Wittmann et al., 2007)、ERP研究(Elbert, Ulrich, Rockstroh, & Lutzenberger,1991)等均证实了1 s以上时距认知具有分段性, 分段临界点大约在2~3 s。
非分段性假说则体现在Church (1984)等的观点上。这一观点虽然没有直接断定数秒以内时距认知不具有分段性, 但都主张数秒以内时距加工机制均采用以“内部时钟”为核心思想的系列模型来解释, 譬如标量计时模型。该模型认为人类判断时间包括时钟、记忆和决策三个水平。时钟阶段通过起搏器以一定的频率产生脉冲, 在起搏器和累加器之间有一开关, 注意时间特征时开关闭合, 脉冲通过开关进入累加器。累加器对脉冲进行累加。记忆阶段由工作记忆和参照记忆组成, 累加器把有关的时间信息传送至工作记忆和参照记忆中, 并形成相应的时间表征。决策阶段主要对工作记忆中的当前时距和参照记忆中的时距表征进行比较, 进而完成判断(Church, 1984)。当然, 在一项元分析研究(Eisler,1976))、双任务研究(Fortin & Couture, 2002)和ERP研究(Gibbons & Rammsayer, 2004)中没有发现1 s以上时距认知具有分段性的证据。
综上所述, 目前关于1 s以上时距认知的分段性(2~3 s的分段临界点)的证据仍存在分歧。本研究拟准备采用两条较新颖的途径探讨该问题。第一条途径是通过比较高、低工作记忆容量(memory working capacity, WMC)的被试在完成1~6 s时距加工的表现来推断时距认知的分段性。WMC是指个体进行在线信息加工过程中同时保持的信息量, 这一指标可以综合体现个体的工作记忆存储能力、信息加工效率及注意控制能力(McCabe, Roediger, McDaniel,Balota, & Hambrick, 2010)。第二条途经是通过比较被试在完成1~6 s时距加工的通道效应(视觉和听觉)来推断时距认知的分段性。回顾以往文献, 仅有一项研究采用过这两条途经(Ulbrich, Churan, Fink, &Wittmann, 2007)。在这项颇具代表性的研究中, Ulbrich等招募了21至84岁的被试, 然后采用Corsi-block测试甄别出高低空间WMC的两组被试, 比较这两组被试对1~5 s的听觉和视觉时距进行复制的成绩,结果发现复制听觉时距时, 两组被试在1~3 s差异不显著, 在4 s和5 s WMC大的被试复制时距长; 复制视觉时距时, 两组被试在1 s、2 s和4 s差异不显著,在3 s和5 s WMC大的被试复制时距长。然而, Ulbrich等(2007)研究可能存在有待商榷之处。(1)被试的筛选:在Ulbrich等(2007)的实验中, 筛选出的高、低WMC被试仅是根据两组被试在Corsi-block测试得分上存在统计学差异, 这不意味着两组被试是“真正”的高、低WMC个体。(2)WMC的测量:Corsi积木测验是测量视觉一空间工作记忆容量的经典任务, 主要应用于测量成年人(Smyth & Scholey, 1992)、儿童(Orsini, Schiappa, & Grossi, 1981)和神经心理障碍患者(Vilkki, Hoist, Öhman, J., Servo, & Heiskanen,1989)的视空间工作记忆。而在Ulbrich等(2007)的实验中涉及视觉和听觉信号的呈现, 所以要测量工作记忆容量就必须兼顾视觉工作记忆和听觉工作记忆两个方面更为合适。(3)额外变量的干扰:不能排除生理节律等无关因素的干扰。在Ulbrich等(2007)的实验中, 高WMC组年龄显著小于低WMC组。年老被试的生理节律周期也下降了, 因此年老被试在复制任务中低估时距, 也可能是由于内部时钟(与生理节律成正相关)速度变慢的影响所致(Block,Zakay, & Hancock, 1998)。
基于此, 本研究拟设计实验1(时间复制任务)和实验2(时间产生任务)来探讨1 s以上时距认知的分段性问题。实验1在Ulbrich等(2007)研究的基础上进行了三个方面的改变:其一, 自变量的测量。在一项研究中, 自变量的测量应该与该研究的目的是一致的。Corsi积木测验是测量视觉一空间工作记忆容量的经典任务, 主要应用于测量成年人(Smyth &Scholey, 1992)、儿童(Orsini et al., 1981)和神经心理障碍患者(Vilkki et al., 1989)的视空间工作记忆。本研究的目的之一是在于考察不同工作记忆容量的个体在完成视觉和听觉通道计时任务的表现, 所以要测量出来的工作记忆容量就必须兼顾视觉工作记忆和听觉工作记忆两个方面。操作广度任务和对称广度任务恰好分别就是两种常见的听觉工作记忆任务和视觉工作记忆任务, 且经常被研究者搭配使用(Conway et al., 2005; Unsworth, Redick, Heitz,Broadway, & Engle, 2009)。其二, 自变量水平的操纵。在Ulbrich等(2007)的实验中, 对WMC的操纵仅是根据Corsi-block测试得分上统计学差异而区分出高、低WMC两个水平, 这样未必能够充分体现自变量的效应。所以, 本研究通过收集大样本正常成年个体的WMC数据, 期待寻找符合正态分布的样本群体, 以确保找到“真正”意义的高、低WMC组被试, 对WMC的水平进行充分的操纵; 其三,额外变量的控制。为了避免生理节律(内部时钟等)的差异所带来的对复制时距的额外影响, 本研究尽可能采用年龄相对较小、年龄区间相对较窄的被试群体(19~35岁), 而Ulbrich等(2007)的实验被试年龄跨度为21~84岁。根据Fraisse和Pöppel的理论假说, 2~3 s以上时间估计受工作记忆及信号通道的影响, 2~3 s以下时间知觉少受注意和工作记忆及信号通道的影响, 那么实验1可以预期高WMC的个体较低WMC的个体复制2~3 s以上时距会更长,而复制2~3 s以下时距不存在显著差异。复制2~3 s以上的听时距较视时距更长, 复制2~3 s以下的听时距与视时距无显著差异。Baudouin, Vanneste, Isingrini和Pouthas (2006)认为不同时间估计方法基于不同加工机制, 时间产生法主要与内部时钟的速度和注意监控有关, 而时间复制法更多地依赖于注意监控和工作记忆。实验2拟要求被试完成时间产生任务,旨在为时距加工分段性寻找新的证据。实验2预期高WMC的个体较低WMC的个体产生2~3 s以上时距会更短, 而产生2~3 s以下时距不存在显著差异。产生2~3 s以上的听时距较视时距更短, 产生2~3 s以下的听时距与视时距无显著差异。
2 实验1 1~6 s时距复制加工分段性特征研究
2.1 目的
实验1以高、低WMC组的个体为被试, 要求他们完成1~6 s的听时距和视时距复制任务, 为1 s以上时距认知的分段性提供新证据。
2.2 方法
2.2.1 被试
本实验中招募的被试是来自事先通过测试被区分出的低WMC和高WMC两组群体。所有被试都来自重庆市沙坪坝区大学城虎溪校区的当地社区的居民或者重庆师范大学、重庆科技学院、重庆医科大学等学校的在校本科生, 年龄介于19~35岁,均签订了知情同意书。为了筛选“真正”的高WMC和低WMC个体, 课题组曾花费1年多时间收集了1000多名被试的WMC测验的数据(操作广度任务和对称广度任务得分), 这些分数一致被认为呈现出正态分布。
操作广度任务和对称广度任务均是把WMC界定为一种与执行控制有关的领域普适性能力的常用测试(Conway et al., 2005; Unsworth et al., 2009)。操作广度任务是一项针对WMC的言语测试。在测试中, 首先给被试学习一系列听觉呈现的字母(譬如F, H, J, K, L, N, P, Q, R, S, T和Y), 然后, 安排被试完成3~7个方程式(每次测试中学习与回忆之间的方程式数量是随机变化的), 继而要求被试通过鼠标在一个4×3的填满12个字母的栅格中按键, 以按着原来顺序回忆刚才学习的字母。为了保证所回忆字母的相对位置, 当被试对某一个字母不能回忆时,按空格键代替(Unsworth, Heitz, Schrock, & Engle,2005)。在该测试中, 采用严格的系列位置计分法,在1个正确的系列中报告正确1个项目得1分。譬如, 如果学习的字母是JRKT, 报告“JRK”得3分,报告“空格键RKT”得3分, 但报告“RKT”得0分。计分由计算机程序自动完成。这种评分方法产生的WMC分数已被证明具有良好的信度和效度(Conway et al., 2005)。操作广度任务的得分范围为(0, 75)。
对称广度任务是一项针对WMC的视觉空间测试。在测试中, 首先给被试学习一系列依次出现在一个4×4栅格中的红色正方形, 然后, 安排被试完成2~5次黑白图片对称判断(每次测试中学习与回忆之间的对称图形判断数量是随机变化的), 继而要求被试通过鼠标在一个4×4的空栅格中按键, 以按着原来顺序回忆刚才学习的红色方块。为了保证所回忆红色方块的相对位置, 当被试对某一个方块不能回忆时, 按空格键代替(Unsworth et al., 2009)。在该测试中, 采用严格的系列位置计分法, 在1个正确的系列中报告正确1个项目得1分。譬如, 如果学习的红色正方形是□□□□, 报告□□□(三个正方形位置均正确, 且属于前三个正方形的位置)得3分, 但是报告□□□(三个正方形位置均正确, 但不属于前三个正方形的位置, 而是后三个正方形的位置)得0分, 然而报告成“空格键□□□” (三个正方形位置均正确, 且是后三个正方形的位置)依然可得3分。对称广度任务得分范围为(0, 42)。
所有被试在两项WMC任务得分如下(原始分数):操作广度作业:M=59.75,SD=12.15, 对称广度作业:M=28.64,SD=8.49。两项WMC任务之间得分相关显著,r=0.58,p< 0.001。每个被试每项WMC任务的得分都被转化为Z分数, 继而求得两项WMC任务的Z分数平均值, 形成复合Z得分。依照个体复合Z分数在所有复合Z分数分布中所处的位置, 筛选出高WMC个体和低WMC个体, 复合Z分数最高的25%为高WMC组, 最低的25%为低WMC组。之后实验1、2的被试均从筛选出的高WMC组和低WMC中随机选取。
最后确定参加实验1的被试44名(22名高WMC,12名女性; 22名低MWC, 14名女性)。两组被试在WMC得分上差异显著,t(42)=−15.24,p< 0.001 (高WMCM=0.99,SD=0.18; 低WMCM=−1.18,SD=0.22)。所有被试均熟悉计算机基本操作, 事先并不知道实验目的, 也未参与过类似实验, 并被告知实验结束之后会获取一定费用。被试无既往精神疾病或神经功能障碍史, 均为右利手。
2.2.2 设备和刺激材料
奔腾Ⅳ (1024 MB内存64 MB显存3.2 GHz处理器, 21″显示器, 1024×768分辨率)一台, 标准MS键盘。采用E-Studio (E-prime V2.0)编写实验程序,并收集数据。为了避免被试容易疲劳, 且研究发现无意识颜色知觉对时距判断无显著影响(Hays,Huybers, & Varakin, 2014), 实验将Ulbrich等(2007)研究中编码阶段和复制阶段的白色刺激和黄色刺激分别调整为黑色正方形和灰色正方形, 正方形的边长为1 cm; 听觉材料为通过扬声器(SONY CD 450)呈现的响度为45 dB的, 频率分别为1000 Hz和1200 Hz的正弦波。
2.2.3 实验设计
采用2×2×6混合设计。WMC (高、低)为被试间变量, 通道(视觉、听觉)和时距长度(1 s、2 s、3 s、4 s、5 s和6 s)为被试内变量。因变量指标是平均时间复制长度、时间复制准确性和时间复制变异性。平均时间复制长度, 即每一种客观时距条件下, 多次复制的平均值; 准确性用复制比率表示, 即每一种条件下主观复制时距长度除以客观时距长度, 复制比率大于1表明高估时间, 小于1表明低估时间, 等于1表明准估时间; 变异性用变异系数表示, 即每一种条件下主观复制时距标准差除以平均值, 变异系数越大意味着变异性越大。
2.2.4 实验程序
练习阶段开始之前, 通过指导语告知被试在时距加工过程中不要采用数数等策略, 这种方法已经在以往研究中被证实是预防数数策略最有效的(Rattat & Droit-Volet, 2012)
刺激流程如图1所示, 首先在屏幕中央呈现一个红色词组”准备”, 待被试准备好之后, 按回车键继续实验。空屏500 ms后, 出现一黑色正方形(或1000 Hz的纯音), 黑色正方形(纯音)呈现的总时间随机选取1~6 s时距之一, 要求被试记住黑色正方形(纯音)呈现的整体时间长度。接着空屏1500 ms,最后屏幕上立刻出现一个灰色正方形(或1200 Hz的纯音), 当觉得灰色正方形和黑色正方形(或两个纯音)持续时间相等时, 按回车键。练习实验时, 每个被试完成12种处理的测试, 每种测试重复2次,共计24次。目的在于让被试熟悉实验刺激的流程。正式试验阶段, 包括4个视觉复制任务组块和4个听觉复制任务组块, 每个组块30次测试, 总共需要完成240次测试, 视觉复制任务组块和听觉复制任务按照ABBA的顺序在被试间平衡, 整个实验过程大概持续约40~50 min。实验结束之后, 主试对实验过程(如是否出现疲劳、误答、程序显示及任务完成过程中的感受问题)进行询问和记录。
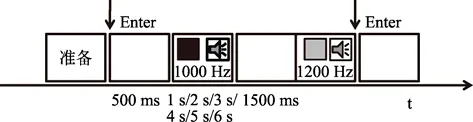
图1 实验1的刺激流程
2.3 结果与分析
以平均时间复制长度为指标, 进行WMC (被试间变量)、通道(视觉、听觉)和时距长度(被试内变量)的重复测量方差分析(见图2)。结果表明, WMC主效应显著,F(1,43)=20.01,p< 0.001,=0.34。时距长度主效应显著,F(5,215)=737.72,p< 0.001,=0.42。事后检验发现, 6 s的平均时间复制长度依次显著长于5 s、4 s、3 s、2 s及1 s,ps < 0.001。通道主效应显著,F(1,43)=13.33,p< 0.001,=0.36。WMC与时距长度交互效应显著,F(5,215)=5.68,p<0.001,=0.51。简单效应分析发现, 1 s和2 s条件下, 高低WMC组复制时间无显著差异,ps> 0.05。3 s、4 s、5 s及6 s条件下, 高WMC组复制时间显著长于低WMC组,ps < 0.001。通道与时距长度交互效应显著,F(5,215)=8.41,p< 0.001,=0.54。简单效应分析发现, 被试复制1 s和2 s视时距与复制听时距无显著差异,ps > 0.05, 被试复制3 s、4 s、5 s及6 s的听时距显著长于复制同一客观长度的视时距,ps < 0.001。其他效应均不显著。
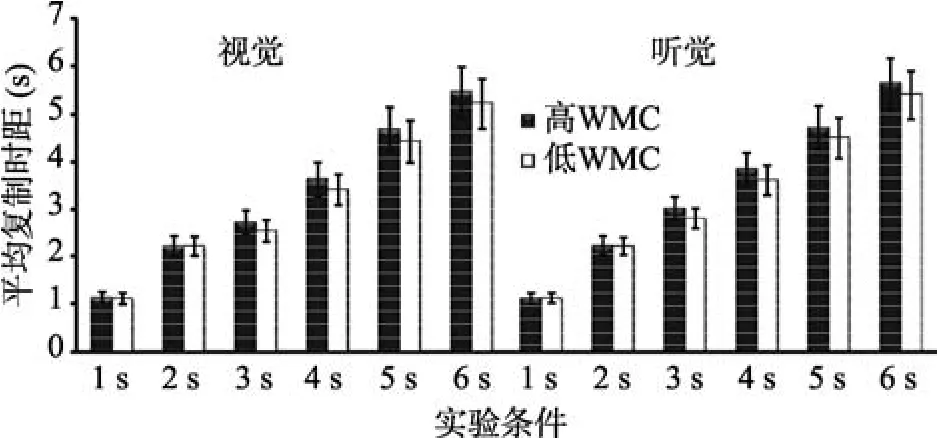
图2 高、低WMC组个体复制1~6 s的平均时距
以复制比率为指标, 进行WMC (被试间变量)、通道(视觉、听觉)和时距长度(被试内变量)的重复测量方差分析(见图3)。结果表明, WMC主效应显著,F(1,43)=15.95,p< 0.001,=0.31。时距长度主效应显著,F(5,215)=28.67,p< 0.001,=0.43。事后检验发现, 6 s的复制比率依次显著大于5 s、4 s、3 s、2 s及1 s,ps < 0.001。通道主效应显著,F(1,43)=14.22,p=0.001,=0.31。WMC与时距长度交互效应显著,F(5,215)=2.45,p=0.043,=0.49。简单效应分析发现, 1 s和2 s条件下, 高、低WMC组复制比率无显著差异,ps > 0.05。3 s、4 s、5 s及6 s条件下, 高WMC组复制比率显著高于低WMC组,ps < 0.001。通道与时距长度交互效应显著,F(5,215)=11.42,p< 0.001,=0.37。简单效应分析发现, 被试复制1 s和2 s视时距与复制听时距的复制比率无显著差异,ps > 0.05, 被试复制3 s、4 s、5 s及6 s的听时距的比率系数显著大于复制同一客观长度的视时距,ps < 0.001。其他交互效应均不显著。
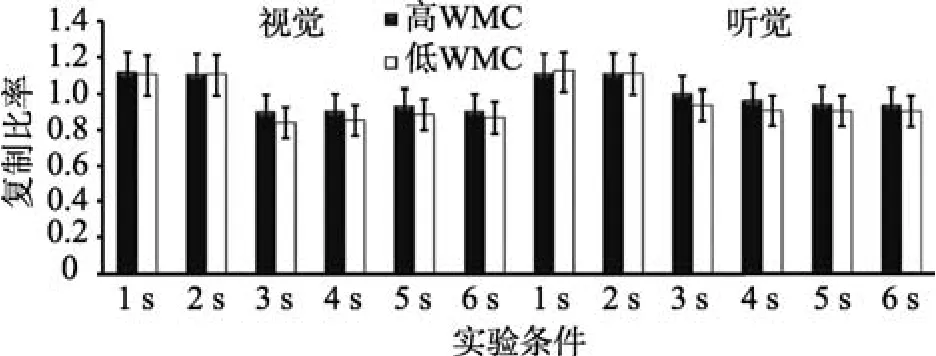
图3 高、低WMC组个体复制1~6 s的比率系数
以变异系数为指标, 进行WMC (被试间变量)、通道(视觉、听觉)和时距长度(被试内变量)的重复测量方差分析(见图4)。结果表明, WMC主效应显著,F(1,43)=18.92,p< 0.001,=0.38。时距长度主效应显著,F(5,215)=28.67,p< 0.001,=0.43。事后检验发现, 6 s的变异系数依次显著小于5 s、4 s、3 s、2 s及1 s,ps < 0.001。通道主效应显著,F(1,43)=24.23,p< 0.001,=0.32。WMC与时距长度交互效应显著,F(5,215)=15.35,p< 0.001,=0.47。简单效应分析发现, 1 s和2 s条件下, 高低WMC组变异系数无显著差异,ps > 0.05。3 s、4 s、5 s及6 s条件下, 高WMC组变异系数显著小于低WMC组,ps < 0.001。通道与时距长度交互效应显著,F(5,215)=13.48,p< 0.001,=0.35。简单效应分析发现, 被试复制1 s和2 s视时距与复制听时距的变异系数无显著差异,ps > 0.05, 被试复制3 s、4 s、5 s及6 s的听时距的变异系数显著小于复制同一客观长度的视时距,ps < 0.001。其他交互效应均不显著。
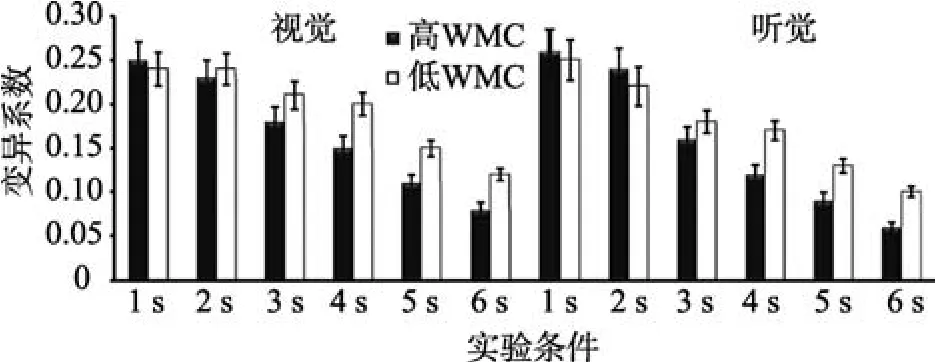
图4 高、低WMC组个体复制1~6 s的变异系数
2.4 小结
实验1通过比较高、低WMC组被试在完成1~6 s视、听时距加工的成绩, 旨在为1 s以上时距认知的分段性寻找新证据。实验1结果发现, 不管哪种WMC类型的被试均表现出高估短时距, 低估长时距的趋势,这与以往多项研究结果一致(Rammsayer &Lima, 1991; Szelag, 1997; Kagerer et al., 2002)。实验1结果还发现, 高、低WMC组在平均复制时距、复制比率及变异系数上均存在主效应, WMC越高的被试平均复制时距越长、复制比率越大、变异系数越小。这可能意味着高WMC个体存储信息容量大,加工信息的变异性较小。同时还发现WMC与时距长度的交互作用以及通道与时距长度的交互作用(1 s、2 s与3 s、4 s、5 s及6 s表现出不同趋势)。根据Fraisse(1984)和Pöppel (1997)的理论假说, 2~3 s以上时间估计受工作记忆及信号通道的影响, 2~3 s以下时间知觉少受注意和工作记忆及信号通道的影响。上述结果明显与Fraisse (1984)和Pöppel (1997)的理论假说是吻合的。
时间产生法是一种不同于时间复制法的方法,涉及认知过程不完全相同。时距产生法由主试给定具体的靶时距值(如2 s) 让被试控制刺激呈现的时距。时距复制法首先由主试呈现一个刺激时距, 然后被试复制一个同样长短的操作时距。Baudouin等(2006)的研究认为不同的时间估计方法基于不同的加工机制, 时间产生法主要与内部时钟的速度和注意控制有关, 而时间复制法更多地依赖于注意监控和工作记忆。实验2拟采用时间产生法要求被试完成时间加工任务, 旨在为1 s以上时距加工分段性提供新证据。
3 实验2 1~6 s时距产生加工分段性特征研究
3.1 目的
实验2以高、低WMC组的个体为被试, 要求他们完成1~6 s听时距和视时距产生任务, 为1 s以上时距加工分段性提供新证据。
3.2 方法
3.2.1 被试
从符合要求的被试选取被试48名(24名高WMC,14名女性; 24名低WMC, 10名女性)。两组被试在WMC得分上差异显著,t(46)=21.15,p< 0.001 (高WMCM=1.01,SD=0.167; 低WMCM=−1.09,SD=0.124)。所有被试均熟悉计算机基本操作, 事先并不知道实验目的, 也未参与过类似实验, 并被告知实验结束之后会获取一定费用。被试无既往精神疾病或神经功能障碍史, 均为右利手。
3.2.2 设备和刺激材料
设备同实验1。时距产生阶段采用的视觉材料为一个黑色正方形, 边长为1 cm; 听觉材料为通过扬声器(SONY CD 450)呈现响度为45 dB、频率为1000 Hz的正弦波。
3.2.3 实验设计
采用2×2×6混合设计。WMC (高、低)为被试间变量, 通道(视觉、听觉)和时距长度(1 s、2 s、3 s、4 s、5 s和6 s)为被试内变量。因变量指标是平均产生时间长度、时间产生准确性和时间产生变异性。平均产生时间长度, 即每一种客观时距条件下, 多次产生时距的平均值; 准确性用产生比率表示, 即每一种条件下主观产生时距长度除以客观时距长度, 产生比率大于1表明高估时间, 小于1表明低估时间, 等于1表明准估时间; 变异性用变异系数表示, 即每一种条件下主观产生时距标准差除以平均值, 变异系数越大意味着变异性越大。
3.2.4 实验程序
刺激流程如图5所示, 首先在屏幕中央呈现一个红色词组“准备”, 待被试准备好之后, 按回车键继续实验。空屏500 ms后, 屏幕上出现所要产生的目标时距(如1 s), 按压回车键同时出现一个刺激(黑色正方形或1000 Hz纯音), 当被试主观感觉刺激呈现时间达到目标时距后, 即按压回车键刺激消失。计算机自动记录被试产生的时距(精确到0.1 s)。
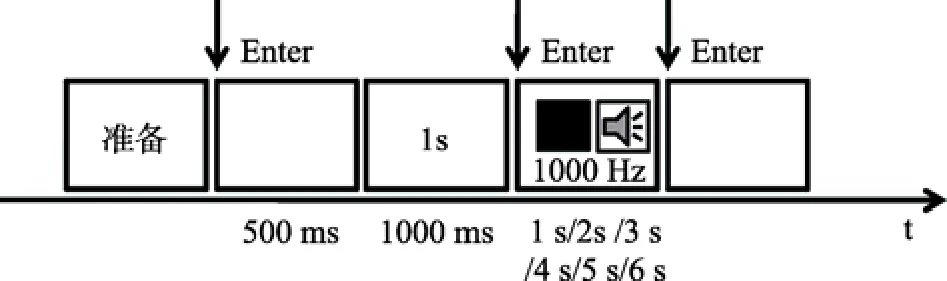
图5 实验2的刺激流程
练习实验时, 每个被试完成12种处理的测试,每种测试重复2次, 共计24次。目的在于让被试熟悉实验刺激的流程。正式试验阶段, 包括4个视觉产生任务组块和4个听觉产生任务组块, 每个组块30次测试, 总共需要完成240次测试, 视觉产生任务组块和听觉产生任务按照ABBA顺序在被试间平衡, 整个实验过程大概持续约20~30 min。
实验结束之后, 主试对实验过程(如是否出现疲劳、误答、程序显示及任务完成过程中的感受问题)进行询问和记录。
3.3 结果与分析
以平均时间产生长度为指标, 进行WMC (被试间变量)、通道(视觉、听觉)和时距长度(被试内变量)的重复测量方差分析(见图6)。结果表明, WMC主效应显著,F(1,47)=25.64,p< 0.001,=0.37。时距长度主效应显著,F(5,235)=694.42,p< 0.001,=0.45。事后检验发现, 6 s的平均时间产生长度依次显著长于5 s、4 s、3 s、2 s及1 s,ps < 0.001。通道主效应显著,F(1,47)=17.31,p< 0.001,=0.32。WMC与时距长度交互效应显著,F(5,235)=12.44,p< 0.001,=0.47。简单效应分析发现, 1 s和2 s条件下, 高低WMC组产生时间无显著差异,ps > 0.05。3 s、4 s、5 s及6 s条件下, 高WMC组复制时间显著短于低WMC组,ps < 0.001。通道与时距长度交互效应显著,F(5,235)=9.14,p< 0.001,=0.43。简单效应分析发现, 被试产生1 s和2 s视时距与产生听时距无显著差异,ps > 0.05, 被试产生3 s、4 s、5 s及6 s的听时距显著短于产生同一客观长度的视时距,ps < 0.001。
其他效应均不显著。
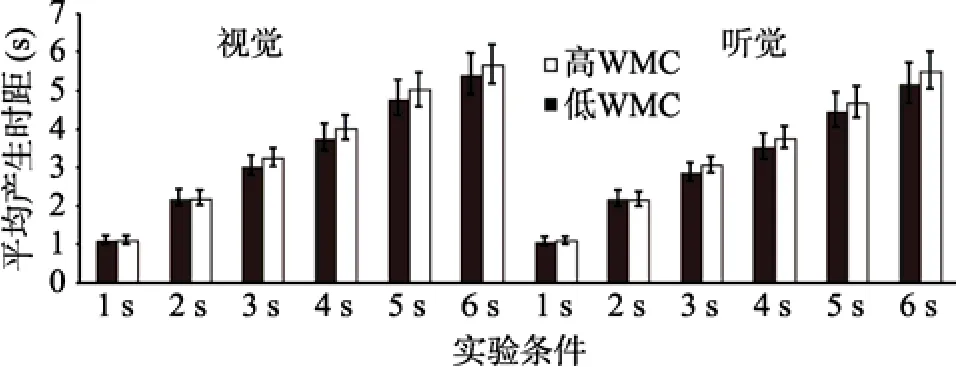
图6 高、低WMC组个体产生1~6 s的平均时距
以产生比率为指标, 进行WMC (被试间变量)、通道(视觉、听觉)和时距长度(被试内变量)的重复测量方差分析(见图7)。结果表明, WMC主效应显著,F(1,47)=18.22,p< 0.001,=0.37。时距长度主效应显著,F(5,235)=21.42,p< 0.001,=0.41。事后检验发现, 6 s的产生比率依次显著大于5 s、4 s、3 s、2 s及1 s,ps < 0.001。通道主效应显著,F(1,47)=14.25,p< 0.001,=0.35。WMC与时距长度交互效应显著,F(5,235)=12.46,p< 0.001,=0.44。简单效应分析发现, 1 s和2 s条件下, 高、低WMC组产生比率无显著差异,ps > 0.05。3 s、4 s、5 s及6 s条件下, 高WMC组产生比率显著小于低WMC组,ps < 0.001。通道与时距长度交互效应显著,F(5,235)=16.44,p< 0.001,=0.35。简单效应分析发现, 被试产生1 s和2 s视时距与产生听时距的比率无显著差异,ps > 0.05, 被试产生3 s、4 s、5 s及6 s的听时距的比率系数显著小于产生同一客观长度的视时距,ps < 0.001。其他交互效应均不显著。
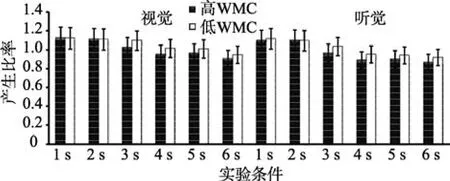
图7 高、低WMC组个体产生1~6 s的比率系数
以变异系数为指标, 进行WMC (被试间变量)、通道(视觉、听觉)和时距长度(被试内变量)的重复测量方差分析(见图8)。结果表明, WMC主效应显著,F(1,47)=20.44,p< 0.001,=0.36。时距长度主效应显著,F(5,235)=22.29,p< 0.001,=0.46。事后检验发现, 6 s的变异系数依次显著小于5 s、4 s、3 s、2 s及1 s,ps < 0.001。通道主效应显著,F(1,47)=21.36,p< 0.001,=0.35。通道与时距长度交互效应显著,F(5,235)=24.45,p< 0.001,=0.39。简单效应分析发现, 被试产生1 s和2 s视时距与听时距的变异系数无显著差异,ps > 0.05, 被试产生3 s、4 s、5 s及6 s的听时距的比率系数显著小于产生同一客观长度的视时距,ps < 0.001。其他交互效应均不显著。
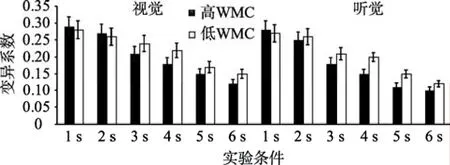
图8 高、低WMC组个体产生1~6 s的变异系数
3.4 小结
实验2通过比较高、低WMC组被试在完成1~6 s视、听时距产生过程中的成绩,旨在为1 s以上时距认知分段性寻找新证据。实验2结果发现, 高、低WMC组在平均产生时距、产生比率及变异系数上均存在主效应, WMC越高的被试平均产生时距越短、产生比率越低、变异系数越小。同时还发现WMC与时距长度以及通道与时距长度的交互作用(1 s、2 s与3 s、4 s、5 s及6 s表现出不同趋势)。根据Fraisse (1984)和Pöppel (1997)的理论假说, 2~3 s以上时间估计受工作记忆及计时信号呈现通道的影响, 2~3 s以下时间知觉少受注意和工作记忆及信号通道的影响。上述结果也明显与Fraisse (1984)和Pöppel (1997)的理论假说吻合。
4 总讨论
实验1和实验2通过比较高、低WMC组被试在完成1~6 s视、听时距复制或者时距产生中的成绩,旨在为1 s以上时距认知的分段性寻找新证据。实验1结果发现, 高、低WMC组在平均复制时距、复制比率及变异系数上均存在主效应, WMC越高的被试平均复制时距越长、复制比率越大、变异系数越小, 这与Ulbrich等(2007)的研究结果基本一致,部分不同点在于Ulbrich等(2007)研究中, 对3 s、4 s、5 s及6 s的复制, 表现出视觉主观时间估计(复制比率)长于听觉主观时间估计(复制比率), 而在实验1中, 3 s、4 s、5 s及6 s的复制, 表现出听觉主观时间估计(复制比率)长于视觉主观时间估计(复制比率), 这与多项研究发现的视、听通道效应是一致的(黄希庭, 郑云, 1993), 即听觉判断要比视觉判断更加精确。另外, 在Ulbrich等(2007)研究中, 以变异系数为指标, 发现了时距长度和通道的主效应, 但是WMC效应只是临界显著, 且没有发现其他交互效应, 而在实验1中, 我们同样发现了时距长度和通道以及WMC的主效应, 但是也发现了时距长度与WMC以及时距长度与通道的交互效应, 这似乎说明了实验1的结果反映出1 s和2 s的时距加工,与3 s、4 s、5 s及6 s的加工模式不太一致。至于两项研究的差异, 一方面可能与两项研究对于WMC的测量不太一致, 另一方面可能也与在两项研究中被试的个体差异性、实验环境、实验程序等因素无法保证完全一致有关。另外, 实验1的结果与Fortin 和 Couture (2002)研究所发现的工作记忆负荷对1.85~6.45 s时距复制的影响无显著差异的结果不一致。这可能是由于两项研究中所采用的实验程序不一致造成的。首先, Fortin和Couture (2002)在研究中采用非时间任务操纵时距复制过程中的工作记忆负荷, 而在实验1中是以自然出现的WMC差异代替记忆负荷的操纵。其次, 在Fortin和Couture(2002)研究中操纵非时间任务的记忆负荷主要是影响时距复制阶段, 而实验1中被试之间的WMC差异是贯穿在整个实验阶段的。在这两项研究中探测工作记忆效应的认知操作, 可能涉及不同记忆成份,执行非时间任务涉及记忆提取加工, 被认为消弱了脉冲累加过程, 导致复制时间偏短或偏长(取决于在编码阶段还是在复制阶段执行非时间任务), 而WMC可能涉及用来表征标准时距脉冲的存储加工能力。由于有限的存储容量, 一部分脉冲可能会随时间流逝或受非时间任务干扰而丢失。实验1的结果支持这一点。在整个时间复制任务过程中, 被试的WMC是一种稳定的特征, 如果累加过程受到影响, 那么高、低组WMC被试应该是没有任何差异,因为WMC对编码阶段(标准时距的脉冲累加过程)和复制阶段(复制时距的脉冲累加过程)应该影响基本一致。然而, 实验1中确实观察到高WMC被试复制时距较长, 因此, WMC涉及关键记忆过程很有可能是标准时距脉冲的存储。换言之, 高WMC的被试对信息保持的好, 导致相对长估, 低WMC被试累加脉冲更容易丢失, 导致相对低估(特别是2~3 s以上时距)。
实验2采用时间产生任务发现, 高、低WMC组在平均产生时距、产生比率及变异系数上均存在主效应, WMC越高的被试平均产生时距越短、产生比率越低、变异系数越小。这可能是因为高WMC的个体在产生过程中注意监控较好, 累加脉冲效率较高, 所以达到目标脉冲数所需的客观时间较少, 所以对同一目标时间(如6 s)而言, 高WMC组产生出的时间较短, 产生比率较低, 变异系数较小。实验1和实验2结果还发现, WMC与时距长度以及通道与时距长度的交互作用, 高、低WMC组被试在加工1 s和2 s的视、听时距过程没有差异, 而加工3 s、4 s、5 s及6 s视、听时距过程存在显著差异, 这暗示着,2~3 s以下与以上时距加工存在两种机制, 2~3 s以下时距加工即为时间知觉, 整体性是时间知觉的一个重要特点。存在一种整合加工将3 s内信息整合一个整体(Fraisse, 1984), θ和α波参与自动时间整合加工(Chen, Chen, Kuang, & Huang, 2015)。Baddeley和Hitch (1974)提出了工作记忆模型, 该模型认为人类主要是通过中央执行系统和语音回路及视空间模板负责对认知任务过程中的信息进行暂时储存与加工。语音回路功能在于暂时储存听觉和语言信息, 视空间模板功能在于保持和操纵视觉和空间信息, 中央执行器涉及这两个系统注意控制。因此,可以说人类在加工时间信息的过程中, 也会需要进行注意控制和暂时储存时间信息。已有研究表明时间工作记忆是不同于空间和词语工作记忆的一种新类型的工作记忆(陈有国, 2010)。实验1和2在工作记忆模型的基础上提出了解释分段性的时间工作记忆说, 其基本观点如下:在加工2~3 s以下时距时, 人能够将2~3 s以下的时距整合为一个整体,整合加工是在感觉记忆加工的基础上进行的, 不受工作记忆和通道的影响; 在2~3 s以上, 时距可能以离散的形式表征, 累加过程受工作记忆和通道的影响。WMC决定了分界点的大小, 老人和小孩的WMC较正常人小, 因此他们的分界点比正常人小(Ulbrich et al., 2007) 。
时间工作记忆说也可以对Lewis等的实验结果进行解释(Lewis & Miall, 2003b)。在Lewis等的实验中, 0.6 s时距位于300 ms分界点附近, 该时距会被整合为一个整体。但由于在分界点附近, 注意对0.6 s时距的调制作用还很弱。3 s时距位于3 s分界点, 这时已经达到WMC的临界点, 工作记忆负荷很大, 需要持续的注意维持时距加工, 所以加工3 s时距比0.6 s时距需要更多的注意资源和更大的工作记忆负荷。Lewis等的实验结果发现, 3 s时距更加激活了前扣带皮质和顶下叶。以往研究发现前扣带皮质与注意控制有关(Posner & Rothbart, 2007),顶下叶皮质可能与工作记忆有关(Owen, McMillan,Laird, & Bullmore, 2005)。时间认知分段综合模型既指出认知时间的分段性, 也指出注意和工作记忆会影响时间信息加工, 因此解释分段性的时间工作记忆说是时间认知分段综合模型解释分段性的具体表现形式(黄希庭等, 2003)。
另外, 分段性的时间工作记忆说只能解释短时距范围的分段性, 还不能解释更长范围内时间认知的分段性。黄希庭等发现过去和未来具有相似的心理结构, 以秒和分为计时单位的“较近的过去”和“较近的未来”,以小时、日和月为计时单位的“近的过去”和“近的未来”, 以及以年为计时单位的“远的过去”和“远的未来” (黄希庭, 孙承惠, 胡维芳,1994; 黄希庭, 1998)。这些分界点的认知基础还有待进一步研究。另外, 随着脑磁图等(CHEN, et al.,2015)技术等迅猛发展, 未来研究可以采用EEG分析, 以alpha波幅为指标, 为2~3 s分段临界点寻找新证据。
5 结论
本研究证明了1~6 s时距认知具有分段性, 分段临界点大约处在2~3 s, 且提出时间工作记忆说解释时间认知的分段性, 该假说是时间认知分段综合模型解释短时距加工分段性的具体表现形式。
Baddeley, A. D., & Hitch, G. (1974). Working memory. In G. A.Bower (Ed.),The psychology of learning and motivation(pp. 48−79). New York: Academic Press.
Baudouin, A., Vanneste, S., Isingrini, M., & Pouthas, V. (2006).Differential involvement of internal clock and working memory in the production and reproduction of duration: A study on older adults.Acta Psychologica, 121(3), 285−296.
Block, R. A., Zakay, D., & Hancock, P. A. (1998). Human aging and duration judgments: A meta-analytic review.Psychology &Aging, 13(4), 584−596.
Cellini, N., Fabbri, M., Martoni, M., Tonetti, L., & Natale, V.(2014). Discontinuity in the perception of sub-second intervals.Procedia - Social and Behavioral Sciences, 126,222−223.
[陈有国. (2010). 时间知觉自动与受控加工的神经机制. 博士论文, 西南大学.]
Chen,Y.G.(2010). Neural mechanisms of automatic and controlled processing of time perception. (Doctoral dissertation, SWNU)
Chen, Y. G., Chen, X., Kuang, C. W., & Huang, X. T. (2015).Neural oscillatory correlates of duration maintenance in working memory.Neuroscience, 290(4), 389−397.
Church, R. M. (1984). Properties of the internal clock.Annals of the New York Academy of Sciences, 423, 566−582.
Conway, A. R. A., Kane, M. J., Bunting, M. F., Hambrick, D. Z.,Wilhelm, O., & Engle, R. W. (2005). Working memory span tasks: A methodological review and user’s guide.Psychonomic Bulletin & Review, 12(5), 769−786.
Cordes, S., & Meck, W. H. (2014). Ordinal judgments in the rat: An understanding of longer and shorter for suprasecond,but not subsecond, durations.Journal of Experimental Psychology: General, 143(2), 710−720.
Eisler, H. (1976). Experiments on subjective duration 1868~1975: A collection of power function exponents.Psychological Bulletin, 83(6), 1154−1171.
Elbert, T., Ulrich, R., Rockstroh, B., & Lutzenberger, W.(1991). The processing of temporal intervals reflected by CNV-like brain potentials.Psychophysiology, 28(6), 648−655.
Fortin, C., & Couture, E. (2002). Short-term memory and time estimation: Beyond the 2-second "critical" value.Canadian Journal of Experimental Psychology-Revue Canadienne De Psychologie Experimentale, 56(2), 120−127.
Fraisse, P. (1984). Perception and estimation of time.Annual Review of Psychology, 35(1), 1−37.
Gibbons, H., & Rammsayer, T. H. (2004). Current-source density analysis of slow brain potentials during time estimation.Psychophysiology, 41(6), 861−874.
Gomez, C., Argandona, E. D., Solier, R. G., Angulo, J. C., &Vazquez, M. (1995). Timing and competition in networks representing ambiguous figures.Brain and Cognition, 29(2),103−114.
Gupta, D. S. (2014). Processing of sub- and supra-second intervals in the primate brain results from the calibration of neuronal oscillators via sensory, motor, and feedback processes.Frontiers in Psychology, 5(3), 816.
Hayashi, M. J., Kantele, M., Walsh, V., Carlson, S., & Kanai,R. (2014). Dissociable neuroanatomical correlates of subsecond and suprasecond time perception.Journal of Cognitive Neuroscience, 26(8), 1685−1693.
Hays, J., Huybers, B., & Varakin, A. (2014). Coloring time!the effect of color in pictures on time perception.Journal of Vision, 14(10), 376-376.
Hirsh, I. J., Monahan, C. B., Grant, K. W., & Singh, P. G. (1990).Studies in auditory timing: 1. Simple patterns.Perception& Psychophysics, 47(3), 215−226.
Huang, X. T. (1994). Psychological structure of future time.Acta Psychologica Sinica, 26(2), 121−127.
[黄希庭. (1994). 未来时间的心理结构.心理学报, 26(2),121−127.]
Huang, X. T., Sun, C. H., Hu, W. F. (1998). Psychological structure of past time.Psychological Science, 30(1), 1−4, 93−94
[黄希庭, 孙承惠, 胡维芳(1998). 过去时间的心理结构.心理科学, 30(1),1−4,93−94.]
Huang, X. T., Li, B. Y., & Zhang, Z. J. (2003). The research of the range-synthetic model of temporal cognition.Journal of Southwest China Normal University (Humanities and Social Sciences Edition), 29(2), 5–9.
[黄希庭, 李伯约, 张志杰. (2003). 时间认知分段综合模型的探讨.西南师范大学学报(人文社会科学版), 29(2), 5–9].
Huang, X. T., & Zheng,Y. (1993). An experimental study of auditory-visual modality effect in time judgement.Acta Psychologica Sinica, 25(3), 225−232.
[黄希庭, 郑云. (1993). 时间判断的视听通道效应的实验研究.心理学报, 25(3), 225−232.]
Kagerer, F. A., Wittmann, M., Szelag, E., & von Steinbüchel, N.(2002). Cortical involvement in temporal reproduction: Evidence for differential roles of the hemispheres.Neuropsychologia,40(3), 357−366.
Kien, J., & Kemp, A. (1994). Is speech temporally segmented?Comparison with temporal segmentation in behavior.Brain and Language, 46(4), 662−682.
Kristofferson, A. B. (1976). Low-variance stimulus-response latencies: Deterministic internal delays?Perception &Psychophysics, 20(2), 89−100.
Lewis, P. A., & Miall, R. C. (2003a). Brain activation patterns during measurement of sub- and supra-second intervals.Neuropsychologia, 41(12), 1583−1592.
Lewis, P. A., & Miall, R. C. (2003b). Distinct systems for automatic and cognitively controlled time measurement:Evidence from neuroimaging.Current Opinion in Neurobiology,13(2), 250−255.
Mates, J., Müller, U., Radil, T., & Pöppel, E. (1994). Temporal integration in sensorimotor synchronization.Journal of Cognitive Neuroscience, 6(4), 332−340.
McCabe, D. P., Roediger III, H. L., McDaniel, M. A., Balota,D. A., & Hambrick, D. Z. (2010). The relationship between working memory capacity and executive functioning: Evidence for a common executive attention construct.Neuropsychology,24(2), 222−243.
Michon, J. A. (1985). The compleat time experiencer. InTime,mind, and behavior(pp. 20−52). Berlin Heidelberg: Springer.
Musterberg, H. (1889).Beiträge zur experimentellen Psychologie:Heft 2. Freiburg, Germany: Akademische Verlagsbuchhandlung.
Orsini, A., Schiappa, O., & Grossi, D. (1981). Sex and cultural differences in children's spatial and verbal memory span.Perceptual and Motor Skills, 53(1), 39−42.
Owen, A. M., Mcmillan, K. M., Laird, A. R., & Bullmore, E.(2005). N-back working memory paradigm: a meta-analysis of normative functional neuroimaging studies.Human Brain Mapping, 25(1), 46−59.
Posner, M. I., & Rothbart, M. K. (2007). Research on attention networks as a model for the integration of psychological science.Annual Review of Psychology, 58(1), 1−23.
Pöppel, E. (1997). A hierarchical model of temporal perception.Trends in Cognitive Sciences, 1(2), 56−61.
Rammsayer, T. H., & Lima, S. D. (1991). Duration discrimination of filled and empty auditory intervals: cognitive and perceptual factors..Attention Perception & Psychophysics,50(6), 565−574.
Rattat, A. C., & Droit-Volet, S. (2012). What is the best and easiest method of preventing counting in different temporal tasks?.Behavior Research Methods, 44(1), 67−80.
Schleidt, M., Eibl-Eibesfeldt, I., & Pöppel, E. (1987). A universal constant in temporal segmentation of human short-term behavior.Naturwissenschaften, 74(6), 289−290.
Smyth, M. M., & Scholey, K. A. (1992). Determining spatial span: The role of movement time and articulation rate.The Quarterly Journal of Experimental Psychology: Section A,45(3), 479−501.
Szelag, E. (1997). Temporal integration of the brain as studied with the metronome paradigm. InTime, temporality, now(pp. 121−131). Berlin Heidelberg: Springer.
Szelag, E., Kowalska, J., Rymarczyk, K., & Pöppel, E. (1998).Temporal integration in a subjective accentuation task as a function of child cognitive development.Neuroscience Letters, 257(2), 69−72.
Szelag, E., von Steinbüchel, N., Reiser, M., Gilles de Langen,E., & Pöppel, E. (1996). Temporal constraints in processing of nonverbal rhythmic patterns.Acta Neurobiologiae Experimentalis, 56(1), 215−225.
Ulbrich, P., Churan, J., Fink, M., & Wittmann, M. (2007). Temporal reproduction: Further evidence for two processes.Acta Psychologica, 125(1), 51−65.
Unsworth, N., Heitz, R. P., Schrock, J. C., & Engle, R. W.(2005). An automated version of the operation span task.Behavior Research Methods, 37(3), 498−505.
Unsworth, N., Redick, T. S., Heitz, R. P., Broadway, J. M., &Engle, R. W. (2009). Complex working memory span tasks and higher-order cognition: A latent-variable analysis of the relationship between processing and storage.Memory,17(6), 635−654.
Vilkki, J., Holst, P., Öhman, J., Servo, A., & Heiskanen, O.(1989). Cognitive deficits related to computed tomographic findings after surgery for a ruptured intracranial aneurysm.Neurosurgery, 25(2), 166−172.
Wittmann, M., Carter, O., Hasler, F., Cahn, B. R., Grimberg,U., Spring, P., … Vollenweider, F. X. (2007). Effects of psilocybin on time perception and temporal control of behaviour in humans.Journal of Psychopharmacology, 21(1),50−64.
Woodrow, H. (1930). The reproduction of temporal intervals.Journal of Experimental Psychology, 13(6), 473−499.
