高山嵩草草甸初级生产力、多样性与土壤因子的关系
2016-01-28石红霄侯向阳师尚礼吴新宏李鹏杨婷婷
石红霄,侯向阳*,师尚礼,吴新宏,李鹏,杨婷婷
(1.甘肃农业大学草业学院, 草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,
甘肃 兰州 730070; 2.中国农业科学院草原所,内蒙古 呼和浩特 010010)
高山嵩草草甸初级生产力、多样性与土壤因子的关系
石红霄1,2,侯向阳1,2*,师尚礼1,吴新宏2,李鹏2,杨婷婷2
(1.甘肃农业大学草业学院, 草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,
甘肃 兰州 730070; 2.中国农业科学院草原所,内蒙古 呼和浩特 010010)
摘要:为了揭示高寒草甸初级生产力与土壤因子及多样性指数的关系,本文选择青藏高原三江源区具有代表性的高山嵩草矮嵩草草地型、高山嵩草杂类草草地型和高山嵩草圆穗蓼草地型,进行植物多样性、生产力及土壤养分的分析研究。结果表明:不同类型高山嵩草草甸生物量、群落多样性、土壤理化性质都存在显著差异(P<0.05),其中高山嵩草圆穗蓼草地生产力最高,物种数最多,土壤营养成分含量最高。相关性分析表明:地上生物量及优良牧草生物量与土壤理化性质呈显著相关(P<0.05),优良牧草生物量与土壤有机质呈极显著正相关(P<0.01)。物种数的多寡与土壤养分呈显著相关(P<0.05),而群落结构的均匀度及多样性是与植物都需要的大量元素钾呈正相关的(P<0.01)。优势物种的竞争能力是通过土壤的有限养分(速效磷)体现的,两者呈显著正相关(P<0.05),因而地上生物量及优良牧草生物量与速效磷呈极显著正相关(P<0.01)。而矿物氮(包含碱解氮)对丰富度指数、初级生产力有不可替代的作用。
关键词:高寒草甸;初级生产力;多样性;土壤理化性质
Relationships between plant diversity, soil property and productivity in an alpine meadow
SHI Hong-Xiao1,2, HOU Xiang-Yang1,2*, SHI Shang-Li1, WU Xin-Hong2, LI-Peng2, YANG Ting-Ting2
1.CollegeofPrataculturalScience,GansuAgriculturalUniversity,KeyLaboratoryofGrasslandEcosystem,MinistryofEducation,Sino-U.S.CentersforGrazingLandEcosystemSustainability,Lanzhou730070,China; 2.GrasslandResearchInstituteofChineseAcademyofAgricultureScience,Hohhot010010,China
Abstract:The relationships among primary productivity, soil factors and plant diversity in alpine meadows were investigated by analyzing plant community diversity, productivity and soil nutrients of three grassland types in the Three Rivers District, Kobresia pygmaea+Kobresia humilis, Kobresia pygmaea+forbs and Kobresia pygmaeaPolygonum macrophyllum meadow. The biomass, community diversity and soil physical and chemical properties of the different alpine meadows types differed significantly (P<0.05). The Kobresia pygmaea+P. macrophyllum meadow had the highest productivity, number of species and soil nutrient content. Correlation analysis showed that forage biomass and soil physical-chemical properties (P<0.05) and soil organic matter (P<0.01) were significantly positively correlated. The number of plant species present was also correlated with soil nutrients (P<0.01), while uniformity and diversity of community structure were positively correlated (P<0.01) with soil potassium content. The competitive advantage of dominant species was associated with tolerance of low fertility(P<0.05), particularly phosphorus. Aboveground biomass and forage biomass and available phosphorus were significantly positively correlated (P<0.01). Nitrogen was associated with species richness and productivity.
Key words:alpine meadow; primary productivity; community diversity; soil properties
生物多样性、生产力与土壤因子的关系对揭示生物多样性与生态系统功能的关系有重要意义[1]。土壤性质的变化直接影响着植被的生长、发育及演替过程[2],同样,植被的变化也改变着土壤的特征。因此,从植被与土壤环境的关系入手,研究高山嵩草草甸不同植被类型群落生产力和土壤环境的变异规律以及二者的相互关系,对区域可持续发展和促进退化高寒草甸的恢复具有指导意义。有关生物多样性、生产力与土壤因子关系的研究多涉及森林群落[3]、草原群落[4]、高寒草甸群落等[5-6],但对不同类型高寒草甸群落物种多样性、生产力与土壤特性定量关系的研究较少。青藏高原是世界上面积最大、海拔最高的高原[7],其主要草地类型为高寒草甸[8]。高寒草甸的健康决定着青藏高原载畜量的稳定,对维持青藏高原高寒草甸的草-畜平衡有重要的作用。而由于长期超载过牧,鼠虫危害、人为破坏等因素的影响,高寒草甸退化已相当严重,畜草矛盾日益尖锐[9]。大面积的草甸退化,威胁着高原草地畜牧业的可持续发展和人类的生存环境[10]。为改善高寒草甸草-畜矛盾,缓解高寒草甸不断恶化的草甸生态系统,本文通过定量分析不同类型高山嵩草(Kobresiapygmaea)草甸群落的生物多样性、生产力与土壤养分的关系,试图从草甸植被、土壤的角度,探究草甸初级生产力、多样性与土壤养分的相互关系。力求为合理利用优良牧草草甸、缓解载畜压力提供科学参考。
1材料与方法
1.1 研究区概况
本研究在农业部玉树高寒草原资源与生态重点野外科学观测试验站试验样地进行,该站位于我国重要江河长江、黄河和澜沧江的源头,青藏高原腹地。具体位置为青海省玉树州称多县珍秦乡,地理坐标为北纬33°24′30″,东经97°18′00″,海拔为4270 m,气候为典型的高原大陆性气候,该气候特点主要表现为:年温差较小,日温差偏大,年平均气温为-5.6~3.8℃,极端气温最高达到28℃,极端最低气温达-48℃。 试验选择高山嵩草草甸3种具有代表性的植被型,即高山嵩草矮嵩草(Kobresiahumilis)型(KK)、高山嵩草圆穗蓼(Polygonummacrophyllum)型(KP)和高山嵩草杂类草型(KH)。3个草地类型样地均为夏秋放牧草场,放牧强度为中度放牧。每个草地类型内设置3个固定样地,各样地面积为100 m2(10 m×10 m)。样地具体情况见表1。
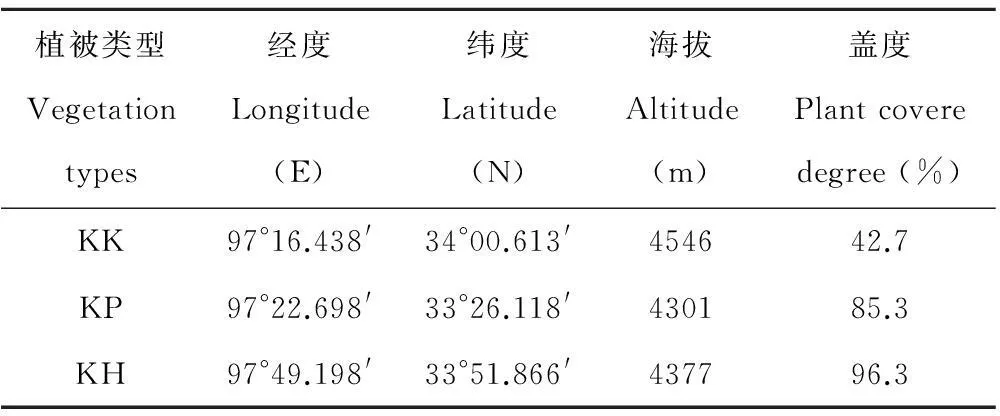
表1 样地基本情况
1.2 试验方法
在研究区内的固定样地内,每个样地分别选择具有典型特征的样方,取1.0 m×1.0 m的3个小样方。调查记录植物高度、盖度、密度、地上生物量、经纬度、海拔等环境因子。地上生物量测定采用齐地面剪割,取样面积为50 cm×50 cm,按照经济类群划分为禾草类、杂类草和莎草类3类,9次重复;将剪下的植株按要求分装后带回室内,80℃恒温箱内烘干至恒重。同时,在剪完植被后的样方上采用对角线法,用取土钻采集0~30 cm土壤,分干、过筛,处理后的土壤样品送检分析土壤理化性质。清选过筛后植物根系清洗干净,自然烘干后分别称根重,为地下生物量。
1.3 数据处理
在野外与试验室进行植物科、属、种的鉴定统计,进行丰富度指数、多样性指数、均匀度指数和生态优势度指数的计算[11],计算公式如下:
丰富度指数:R=S
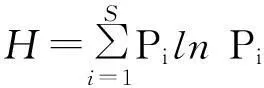
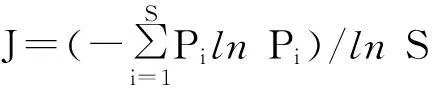
生态优势度指数:I=(相对高度+相对盖度+相对密度)/3
式中,Pi为种i的相对重要值[(相对高度+相对盖度+相对密度)/3],S为种i所在样方的物种总数。
1.4 统计分析
利用SPSS13.0进行单因素最小显著差数法统计分析,一元线性回归及双变量相关分析各草甸初级生产力与土壤因子的关系。
2结果与分析
2.1 高山嵩草草甸群落生产力
从表2可以看出,不同植被类型之间地上生物量、优良牧草生物量、杂类草生物量、地下生物量都存在显著差异(P<0.05)。其中,地上总生物量和优良牧草生物量为KP>KK>KH,杂类草生物量为KK>KH>KP;从表2可以看出高山嵩草草甸3种不同植被型地下生物量主要集中在0~10cm的土层中,地下总生物量、地下(0~10cm)、地下(10~20cm)均为KK>KH>KP。总生物量为KK>KP>KH,说明高山嵩草矮嵩草植被型群落生产力最高,高山嵩草圆穗蓼型次之,高山嵩草杂类草型最差。
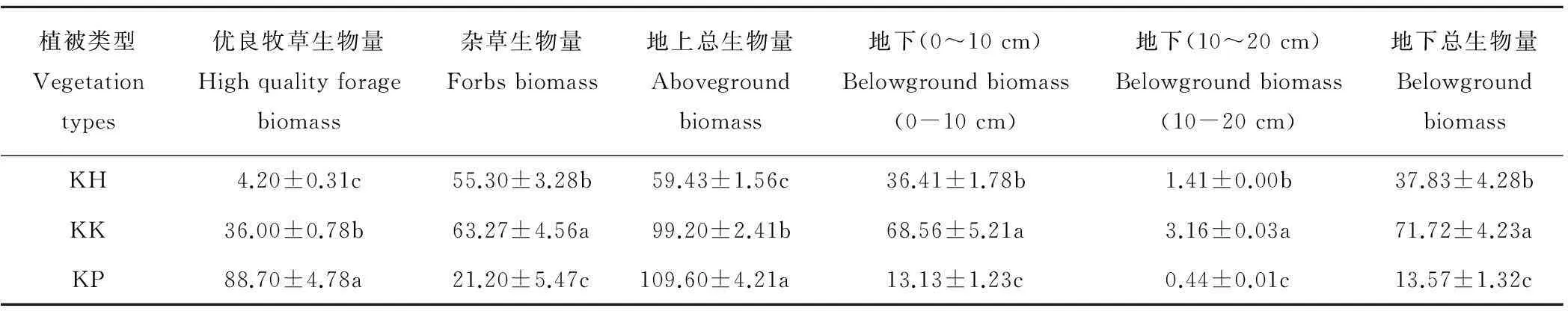
表2 不同植被类型生物量
注: 同一列不同字母表示生境之间有显著差异(P<0.05)。 下同 。
Note: Different letters indicate significant differenceP<0.05). The same below.
2.2 高山嵩草草甸群落生物多样性
不同类型草甸的物种丰富度、均匀度、生态优势度和群落多样性指数都存在显著差异(P<0.05)(表3)。高山嵩草圆穗蓼型群落物种丰富度最高,为24种,高山嵩草矮嵩草型和高山嵩草杂类草型物种丰富度为17种,说明高山嵩草圆穗蓼型群落有较多的物种组成;群落均匀度指数和多样性指数高山嵩草杂类草型最高,分别为0.65和1.85,高山嵩草圆穗蓼型次之,高山嵩草矮嵩草型最低;生态优势度指数高山嵩草矮嵩草型最高,为0.42,高山嵩草圆穗蓼型次之, 高山嵩草杂类草型最低。
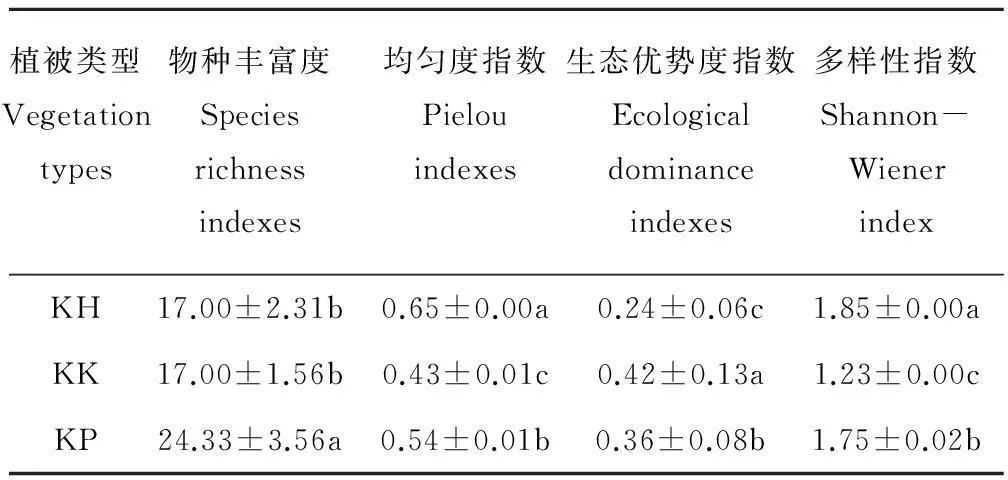
表3 不同植被类型群落多样性
2.3 高山嵩草草甸土壤理化性质
不同植被类型土壤理化性质差异显著(P<0.05)(表4)。其中,高山嵩草圆穗蓼样地的有机质、全氮、全磷、全钾、碱解氮、速效钾、速效磷均为最大,分别为268.02 g/kg, 18.25 g/kg, 4.10 g/kg, 45.78 g/kg, 844.00 mg/kg, 883.23 mg/kg, 19.57 mg/kg;而高山嵩草杂类草样地的有机质、速效磷为最小值(144.60 g/kg和9.37 mg/kg);高山嵩草矮嵩草样地的全氮、全磷、全钾、碱解氮以及速效钾均为最小值。
2.4 生物量和群落多样性与土壤理化性质的相关分析
高山嵩草草甸不同植被型群落多样性、生产力以及土壤理化性质的相关性不同(表5)。相关性分析表明:高山嵩草草甸物种丰富度指数与土壤全氮、有机质、速效钾、速效磷及碱解氮呈极显著正相关(P<0.01);均匀度指数、群落多样性指数与全钾呈极显著正相关(P<0.01);而优势度指数与全钾呈极显著负相关(P<0.01),与速效磷呈显著正相关(P<0.05)。地上生物量与土壤有机质呈显著正相关(P<0.05);优良牧草生物量与土壤全氮、有机质、速效钾、速效磷及碱解氮呈极显著正相关(P<0.01);杂类草生物量、地下生物量与土壤全氮、全磷、速效钾及碱解氮呈极显著负相关(P<0.01),而地下生物量与土壤全钾呈显著负相关(P<0.05)。
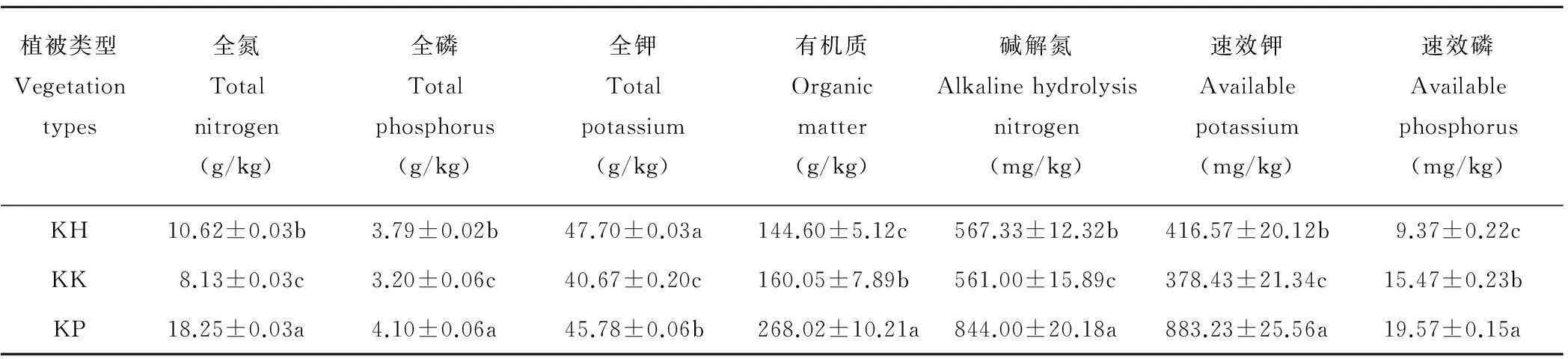
表4 不同植被类型土壤理化性质
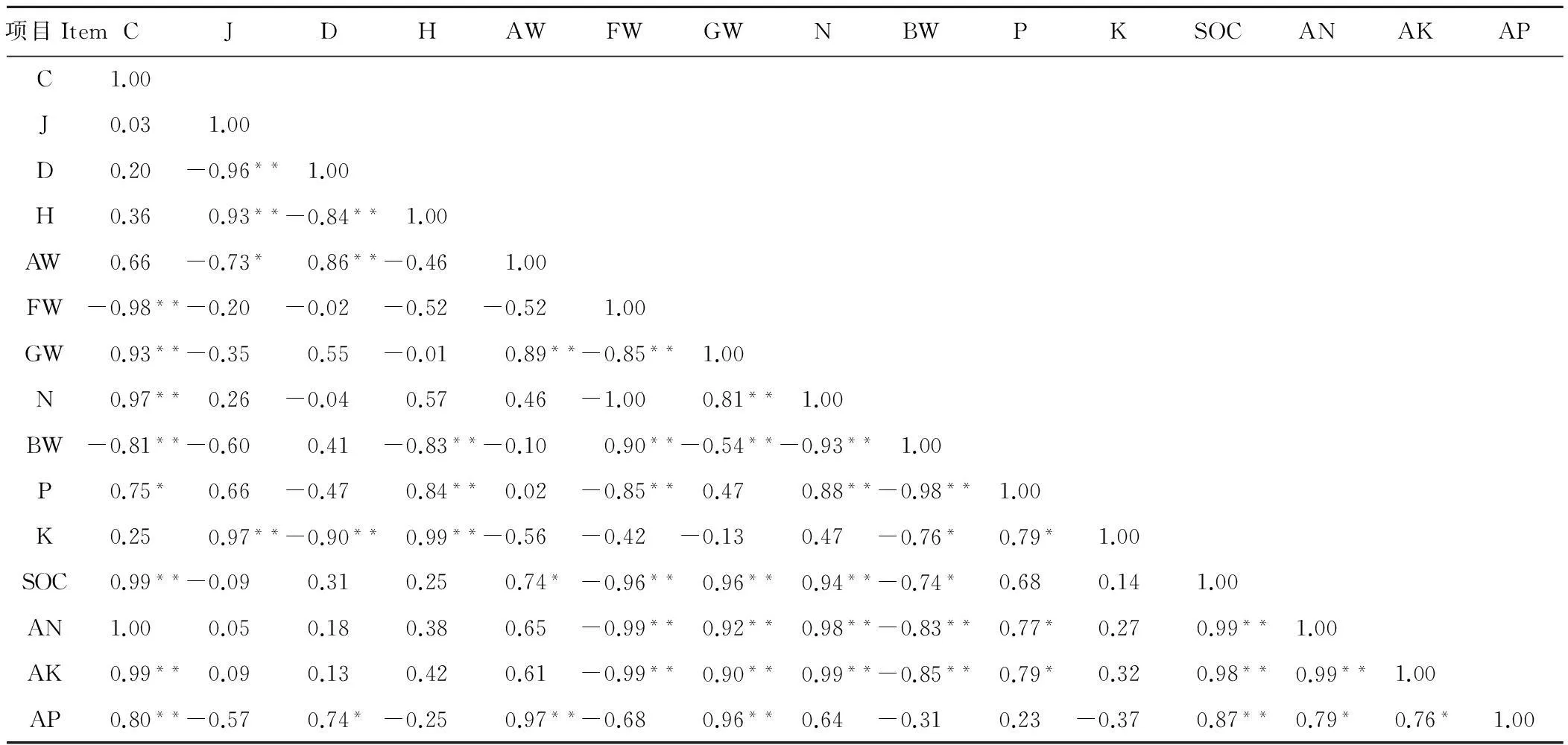
表5 不同草甸类型土壤理化性质与生产力和多样性的关系
C为物种丰富度指数;J为均匀度指数;D为生态优势度指数;H为多样性指数;AW代表地上生物量;FW代表杂类草生物量;GW代表优良牧草生物量;BW代表地下生物量;N代表土壤全氮;P代表土壤全磷;K代表土壤全钾;SOC代表土壤有机质;AN代表土壤碱解氮;AP代表土壤速效磷;AK代表土壤速效钾;*表示P<0.05,**表示P<0.01。
C: species richness indexes; J: Pielou indexes; D: ecological dominance indexes; H: Shannon-Wiener; AW: aboveground biomass; FW: forbs biomass; GW: high quality forage biomass; BW: belowground biomass; N: total nitrogen; P: total phosphorus; K: total potassium; SOC: organic matter; AN: alkaline hydrolysis nitrogen; AP: available phosphorus; AK: available potassium; *indicates significant atP<0.05 level and ** indicates significant atP<0.01 level.
3结论和讨论
3.1 高山嵩草草甸生产力与群落多样性
物种丰富度、均匀度和生态优势度指数不仅是反映群落组成结构特征的定量指标,而且能反映各物种对环境的适应能力和对资源的利用能力[12]。本文研究表明,高山嵩草圆穗蓼植被型,物种丰富度指数高,物种数较多,较多的物种占据相似的生态位,不同物种的竞争使得均匀度指数提高;相反,丰富度指数降低,物种数减少,寡优势种的群落结构使得优势种的竞争力提高,生态优势度指数显著提高[13]。对于青藏高原高寒草甸生物多样性与生产力之间的关系,有研究表明物种丰富度和多样性指数与地上生物量呈显著正相关[14-15],这与本研究物种丰富度与生产力呈显著正相关,而多样性指数与地上生物量呈负相关的结论是不一致的,与朱源等[16]研究高山嵩草+珠芽蓼(Polygonumviviparum)群落生物多样性与生产力没有显著关系的结论是一致的。一般认为,生物多样性与地上生物量呈现单调的关系,往往是调查不足导致的[16]。Wang等[1]指出,生物量的分配特征是由优势物种决定的。高山嵩草矮嵩草和高山嵩圆穗蓼群落的优势度指数达到了0.42和0.36,两者对地上生物量的贡献率分别为81%和37%,表明地上生物量的分配与优势种的生物量是呈极显著正相关的(P<0.01);但杂草生物量与丰富度指数及优良牧草生物量呈极显著负相关(P<0.01),表明杂草的入侵,改变了草甸生态系统的平衡,不利于高寒草甸植被结构的稳定[10]。地下生物量的高低、分配与地上生物量不呈显著相关,这主要是高寒草甸莎草科植物形成近地表的根状茎[17],独特的“地毯式”地表结构,不定根盘根错节;同时,青藏高原长年较低的气温使得“地毯”结构非常稳定,从而在近地表贮存大量的生物量[18],而以双子叶植物为主的杂类草则并不存在这种结构[19]。
3.2 多样性与土壤理化性质
Chandra等[20]指出,动物对不同植物的可食率,人类对某些植物的过度利用、放牧等因素都影响多样性指数;而Kala[21]指出,随着海拔的增加,物种多样性和丰富度指数都呈降低的趋势。本文在海拔、休牧期等条件相似的情况下,探讨了土壤理化性质对多样性、均匀度指数等的影响。从表3可以看出,全氮、有机质、速效钾、速效磷及碱解氮与丰富度指数呈极显著正相关(P<0.01);全钾与均匀度指数、群落多样性指数呈极显著正相关(P<0.01),而与优势度指数呈显著负相关(P<0.05);全磷与群落多样性指数呈现极显著正相关(P<0.01);而速效磷与优势度指数呈显著正相关(P<0.05)。这与白永飞等[22]对锡林河流域草原植物群落的研究结果相似。物种多样性不仅能够度量群落的组成结构和功能的复杂性,而且也能指示环境状况。有关土壤养分与物种多样性的关系存在不同看法,多数人认为,植物群落高的物种多样性出现在土壤养分梯度的中间位置[23-24]。有研究表明,全氮[19]对初级生产力、植物丰富度有重要作用,而且矿物氮(包含碱解氮)对丰富度指数、初级生产力更为重要。Hrevušová等[25]指出,施入氮肥能够促进植物对磷的吸收,使得速效磷较低的草甸能维持相对较高的物种数。除此之外,Tilman[26]也指出,在较低肥力的土壤中,群落的物种丰富度的变化是受有限营养的竞争驱动的。显然,速效磷对群落结构具有重要作用。有学者[27]指出,钾对物种组成是没有影响的,这与本实验结果是不一致的。分析原因,钾是植物生长必须的大量元素[28],土壤中的速效钾对植物的生长具有重要作用,当土壤中含有充足的钾离子时,各物种均能生长,因而钾与均匀度指数、多样性指数呈显著正相关(P<0.01),而与优势度指数呈负相关。而Crawley等[29]探讨的则是钾肥对牧草生长的作用,如果施入大量钾肥,钾对植物的作用就会减小。
3.3 生物量与土壤理化性质
高寒草甸的初级生产力是通过生物量反映的,高寒草甸生物量的积累主要依靠光合作用,而只有为光合作用提供能量和化学物质,植物才能积累有机质。由此可见,土壤理化性质对高寒草甸生物量的积累具有重要的作用。生物量尤其是地下生物量与土壤理化性质呈显著差异(P<0.05);不同草甸土壤理化性质也呈显著差异(P<0.05),表明植物的生物量和土壤理化性质是显著相关的[30]。本文研究指出,地上生物量及优良牧草生物量与速效磷呈极显著正相关(P<0.01)。根据德国化学家Liebig的“最小限制因子率”,作物产量不受大量元素限制,而受微量的营养元素限制。而速效磷是能直接被植物吸收的磷组分,是反映土壤养分供应能力的重要指标[31],是地上生物量的限制因子[29]。 有研究表明[32],当磷含量低于10 g/kg时,牧草初级生产力显著降低。本文发现,3种草甸土壤的速效磷含量是除全磷以外的最小值,表明速效磷是地上生物量的最小限制因子。地上生物量、优良牧草生物量与速效磷具有强的正相关(P<0.01)则证实了笔者的推测。
生物量尤其是地下生物量显著影响到土壤有机质和矿物质层[31]。优良牧草生物量与土壤有机质呈极显著正相关(P<0.01),反映出土壤有机质的主要来源是植物死亡的根系以及枯落物;反之,土壤有机质是土壤微生物利用的主要碳源,微生物改善土壤特性和结构,提高土壤肥力,从而有助于植物的生长。优良牧草生物量及地下生物量与全氮、全磷、碱解氮、速效钾以及地上生物量/地下生物量呈极显著负相关(P<0.01),而杂草生物量则相反。有研究表明[33-34]:全氮、全磷、碱解氮、速效钾等被认为是限制生物量的主要因子,而对于健康的草甸生态系统,优良牧草占据了群落的主要生态位,在不同植被共同竞争土壤营养元素的情况下,杂草完全处于劣势。
从上文的分析中,地下生物量(0~10 cm)远高于地下生物量(10~20 cm),而地下生物量与全氮、全磷、全钾、有机质及速效氮呈极显著负相关(P<0.01)。高寒草甸土壤营养元素集中在土壤表面[35],根系的趋肥效应使得地下生物量主要集中在0~10 cm土壤;另外,高寒草甸具有独特的“地毯式”地表结构[7],地下生物量多半是腐殖质,或者是已死亡的根状茎,因而已停止生长,那么土壤营养元素对其的生长并无多大的作用。
References:
[1]Wang Z R, Yang G J, He X B,etal. Relationship between plant community and environmental factors in the source regions of Yangtze River. Journal of Glaciology and Geocryology, 2011, 33(3): 640-645.
[2]He J S, Fang J Y, Ma K P,etal. Biodiversity and ecosystem productivity: why is there a discrepancy in the relationship between experimental and natural ecosystems. Acta Phytoecologica Sinica, 2003, 27(6): 835-843.
[3]Wang H M, Zhang F, Pang C H,etal. Interrelation between plant species diversity and soil factors in the middle and lower reaches of Fenhe River. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(10): 2077-2085.
[4]Bai K Y, Rong Y P, Yang Y H,etal. Relationships between grassland biodiversity and primary productivity and soil condition in farming-pastoral regions of northern China. Chinese Journal of Ecology, 2013, 32(1): 22-26.
[5]Wang X T, Zhang S H, Chen D D,etal. The effects of natural graszing intensity on plant community and soil nutrients in alpine meadow. Acta Agrestia Sinica, 2010, 18(4): 510-516.
[6]Wang C T, Long R J, Wang Q J,etal. Distribution of organic matter, nitrogen and phosphorus along an altitude gradient and productivity change and their relationships with environmental factors in the alpine meadow. Acta Prataculturae Sinica, 2005, 14(4): 15-20.
[7]Wang J L, Zhong Z M, Wang Z H,etal. Soil C/P distribution characteristics of alpine steppe ecosystems in the Qinghai Tibetan Plateau. Acta Prataculturae Sinica, 2014, 23(2): 9-19.
[8]Shang Z H, Long R J. Formation reason and recovering problem of the ‘black soil type’ degraded alpine grassland in Qinghai-Tibetan Plateau. Chinese Journal of Ecology, 2005, 24 (6): 652-656.
[9]Song R D, Hasagawa N, Li G M,etal. Botanical composition and grazing behaviour of Qinghai Yaks of plateau type in the natural rangeland. Acta Ecologiae Animalis Domastici, 2008, 29(5): 31-35.
[10]Dong Q M, Zhao X Q, Ma Y S,etal. Effects of stocking rate and grazing time of yaks on soil nutrient contents inKobreciaparvaalpine meadow. Chinese Journal of Ecology, 2005, 24(7): 729-735.
[11]Smith R S, Rushton S P. The effects of grazing management on the vegetation of mesotrophic (meadow) grassland in Northern England. Journal of Applied Ecology, 1994, 31: 13-24.
[12]Shan G L, Xu Z, Ning F. The changes of community structure and species diversity in different succession stage in typical steppe. Journal of Arid Land Resources and Environment, 2010, 24(2): 163-169.
[13]Dong Q M, Zhao X Q, Ma Y S,etal. Niche of main plant populations on a warm-seasonal pastureland of alpineKobreciaparvameadow. Chinese Journal of Ecology, 2006, 25(11): 1323-1327.
[14]Tan G L, Du G Z , Li Z Z ,etal. Relationship between species richness and productivity in an alpine meadow plant community. Acta Phytoecologica Sinica, 2002, 26 (Supp l): 57-62.
[15]Zuo X A, Zhao X Y, Zhao H L,etal. Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin Sand Land. Environmental Science, 2007, 28(5): 945-951.
[16]Zhu Y, Kang M Y, Liu Q R,etal. Relationship between biodiversity and aboveground biomass in Alpine Meadow on M t Helan China. Chinese Journal of Applied and Environmental Biology, 2007, 13(6): 771-776.
[17]Wang Q J, Wang W Y, Deng Z F. The dynamics of biomass and the allocation of energy in alpineKobresiameadow communities, Haibei region of Qinghai province. Acta Phytoecologica Sinica, 1998, 22(3):222-230.
[18]Farley R A, Fitter A H. The responses of seven co-occurring woodland herbaceous perennials to localized nutrient-rich patches. Journal of Ecology, 1999, 87:849-859.
[19]Hejcman M, Klaudisová M,tursa J,etal. Revisiting a 37 years abandoned fertilizer experiment on Nardus grassland in the Czech Republic. Agriculture, Ecosystems & Environment, 2007, 118: 231-236.
[20]Chandra P R, Kala A, Singh S K,etal. Effects of sheep and goat grazing on the species diversity in the alpine meadows of Western Himalaya. The Environmentalist, 2002, 22: 183-189.
[21]Kala C P. Ecology and Conservation of Alpine Meadows in the Valley of Flowers National Park, Garhwal Himalaya[M]. Dehradun: Forest Research Institute, 1998: 180.
[22]Bai Y F, Li L H, Wang Q B,etal. Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River basin, Inner Mongolia. Acta Phytoecologica Sinica, 2000, 24(6): 667-673.
[23]Zhang L J, Yue M, Gu F X,etal. Coupling relationship between plant communities species diversity and soil factors in ecotone between desert and oasis in Fukang, Xinjiang. Chinese Journal of Applied Ecology, 2002, 13(6): 658-662.
[24]Luo Y Y, Meng Q T, Zhang J H,etal. Species diversity and biomass in relation to soil properties of alpine meadows in the eastern Tibetan Plateau in different degradation stages. Journal of Glaciology and Geocryology, 2014, 36(5): 1298-1305.
[26]Tilman D. Resource Competition and Community Structure[M]. Princeton: Princeton University Press, 1982.
[28]Janssens F, Peeters A, Tallowin J R B,etal. Relationship between soil chemical factors and grassland diversity. Plant and Soil, 1998, 202: 69-78.
[29]Crawley M J, Johnston A E, Silvertown J,etal. Determinants of species richness in the Park Grass Experiment. The American Naturalist, 2005, 165: 179-192.
[30]Cheng Y H, Zhao R X, Dong K H. Research on plasma K+channel protein. Journal of Shanxi Agricultural Sciences, 2008, 36(2): 3-7.
[31]Zhang H M, Wang Z Y, Yu R G,etal. Study on soil physical and chemical properties under different inter-cropping for orchards on Hilly region in northern Jiangxi province. Research of Soil and Water Conservation, 2010, 17(4): 258-261, 268.
[33]Vanguelova1 E I, Nortcliff S, Moffat A J,etal. Morphology, biomass and nutrient status of fine roots of Scots pine (Pinussylvestris) as influenced by seasonal fluctuations in soil moisture and soil solution chemistry. Plant and Soil, 2005, 270: 233-247.
[34]Güsewell S. High nitrogen: phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytologist, 2005, 166: 537-550.
[35]Honsová D, Hejcman M, Klaudisová M,etal. The species composition of an alluvial meadow in the Czech Republic after 40 years of applying nitrogen, phosphorus and potassium fertilizer. Preslia, 2007, 79: 245-258.
参考文献:
[1]王增如, 杨国靖, 何晓波, 等. 长江源区植物群落特征与环境因子的关系. 冰川冻土, 2011, 33(3): 640-645.
[2]贺金生, 方精云, 马克平, 等. 生物多样性与生态系统生产力:为什么野外观测和受控实验结果不一致. 植物生态学报, 2003, 27(6): 835-843.
[3]王慧敏, 张峰, 庞春花, 等. 汾河流域中下游植物群落物种多样性与土壤因子的关系. 西北植物学报, 2013, 33(10): 2077-2085.
[4]白可喻, 戎郁萍, 杨云卉, 等. 北方农牧交错带草地生物多样性与草地生产力和土壤状况的关系. 生态学杂志, 2013, 32(1): 22-26.
[5]王向涛, 张世虎, 陈懂懂, 等. 不同放牧强度下高寒草甸植被特征和土壤养分变化研究. 草地学报, 2010, 18(4): 510-516.
[6]王长庭, 龙瑞军, 王启基, 等. 高寒草甸不同海拔梯度土壤有机质氮磷的分布和生产力变化及其与环境因子的关系. 草业学报, 2005, 14(4): 15-20.
[7]王建林, 钟志明, 王忠红, 等. 青藏高原高寒草原生态系统土壤碳磷比的分布特征. 草业学报, 2014, 23(2): 9-19.
[8]尚占环, 龙瑞军. 青藏高原“黑土型”退化草地成因与恢复. 生态学杂志, 2005, 24(6): 652-656.
[9]宋仁德, 长谷川信美, 李国梅, 等. 天然草地放牧牦牛采食行为及食性选择的研究. 家畜生态学报, 2008, 29(5): 31-35.
[10]董全民, 赵新全, 马玉寿, 等. 牦牛放牧率和放牧季节对小嵩草高寒草甸土壤养分的影响. 生态学杂志, 2005, 24(7): 729-735.
[12]单贵莲, 徐柱, 宁发. 典型草原不同演替阶段群落结构与物种多样性变化. 干旱区资源与环境, 2010, 24(2): 163-169.
[13]董全民, 赵新全, 马玉寿, 等. 高寒小嵩草草甸暖季草场主要植物种群的生态位. 生态学杂志, 2006, 25(11): 1323-1327.
[14]覃光莲, 杜国祯 , 李自珍, 等. 高寒草甸植物群落中物种多样性与生产力关系研究. 植物生态学报, 2002, 26(Supp l): 57-62.
[15]左小安, 赵学勇, 赵哈林, 等. 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系. 环境科学, 2007,28(5):945-951.
[16]朱源, 康慕谊, 刘全儒, 等. 贺兰山高山草甸生物多样性和地上生物量的关系. 应用与环境生物学报, 2007, 13(6): 771-776.
[17]王启基, 王文颖, 邓自发. 青海海北地区高山嵩草草甸植物群落生物量动态及能量分配. 植物生态学报, 1998, 22(3): 222-230.
[22]白永飞, 李凌浩, 王其兵, 等. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究. 植物生态学报, 2000, 24(6): 667-673.
[23]张林静, 岳明, 顾峰雪, 等. 新疆阜康绿洲荒漠过渡带植物群落物种多样性与土壤环境因子的耦合关系. 应用生态学报, 2002, 13(6): 658-662.
[24]罗亚勇, 孟庆涛, 张静辉, 等. 青藏高原东缘高寒草甸退化过程中植物群落物种多样性、生产力与土壤特性的关系. 冰川冻土, 2014, 36(5): 1298-1305.
[30]程钰宏, 赵瑞雪, 董宽虎. 植物钾(K+)离子通道的研究. 山西农业科学, 2008, 36(2): 3-7.
[31]张华明, 王昭艳, 喻荣岗, 等. 汤崇军赣北丘陵区果园不同套种模式对退化红壤理化性质的影响. 水土保持研究, 2010, 17(4): 258-261, 268.
《草业学报》
《草业学报》由中国科协技术协会主管,中国草学会、兰州大学主办,国内外公开发行的高级学术期刊。2013年《草业学报》的影响因子为2.443,在全国被统计的1989种期刊中排名第4位,现入编全国中文核心期刊,荣获“百强报刊”,“百种中国杰出学术期刊”,“中国精品科技期刊”,“中国科技论文在线优秀期刊一等奖”。
《草业学报》主要报道国内外草业科学及其相关领域,如畜牧学、农学、林学、经济学等领域的高水平理论研究和技术创新成果,发表国内外草业领域创新性的研究论文,刊载学术价值较高的草业科学专论、综述、评论等,探讨草业发展的新理论与新构思,是草业新秀成长的园地,推动草业科学发展的论坛。其读者对象主要是从事农林牧渔、园林绿化、生态环境、国土资源等领域的科研管理及教学等专业人员。
本刊为:中国科学引文数据库(CSCD)核心期刊,中国科技论文统计源期刊,英国CABI文摘数据库来源期刊,《中国生物学文摘》中国生物学文摘数据库收录期刊,中国核心期刊(遴选)数据库收录期刊,《中国学术期刊(光盘版)》全文收录期刊,《万方数据—数字化期刊群》入网期刊。
《草业学报》为月刊,全铜版印刷,逢每月20日出版,大16开本,250页,定价25元,全年300元。国内邮发代号:54-84,全国各地邮局均可订阅,若错过订期,可在本编辑部直接办理订阅。
地址:兰州市嘉峪关西路768号《草业学报》编辑部;邮政编码:730020;网址:http://cyxb.lzu.edu.cn;
电话、传真:0931-8913494;E-mail:cyxb@lzu.edu.cn
http://cyxb.lzu.edu.cn
石红霄,侯向阳,师尚礼,吴新宏,李鹏,杨婷婷. 高山嵩草草甸初级生产力、多样性与土壤因子的关系. 草业学报, 2015, 24(10): 40-47.
SHI Hong-Xiao, HOU Xiang-Yang, SHI Shang-Li, WU Xin-Hong, LI-Peng, YANG Ting-Ting. Relationships between plant diversity, soil property and productivity in an alpine meadow. Acta Prataculturae Sinica, 2015, 24(10): 40-47.
通讯作者*Corresponding author. E-mail:houxy16@126.com
作者简介:石红霄(1980-),男,青海门源人,助理研究员,在读博士。E-mail:axiao8003@sina.com
基金项目:国家重点基础研究发展计划(973计划)(2014CB138802),国家科技支撑计划课题(2012BAD13B07)和中央级公益科研院所基本科研业务专项(1610332014006)资助。
收稿日期:2014-12-08;改回日期:2015-02-11
DOI:10.11686/cyxb2014509
