饥饿诱导胃癌细胞自噬发生及Beclin-1表达的变化
2016-01-28王丽媛
沈 薇 隋 璐 赵 慧 王丽媛
(沈阳医学院病理生理教研室,辽宁 沈阳 110034)
饥饿诱导胃癌细胞自噬发生及Beclin-1表达的变化
沈薇隋璐赵慧1王丽媛2
(沈阳医学院病理生理教研室,辽宁沈阳110034)
摘要〔〕目的探讨饥饿诱导胃癌细胞MKN1自噬的发生及自噬相关蛋白Beclin-1表达的变化。方法培养胃癌细胞MKN1至对数生长期后,以无氨基酸培养液EBSS代替RPMI1640培养液培养细胞,培养12h后收集细胞,应用Western印迹和定量PCR(qRT-PCR)方法分别检测特异性标记自噬的微管相关蛋白1轻链3B(LC3B)以及自噬相关蛋白Beclin-1的蛋白和mRNA表达。免疫荧光检测饥饿诱导后细胞自噬体的产生。结果饥饿处理12 h后,Western印迹和qRT-PCR 检测LC3B和Beclin-1蛋白和mRNA表达与对照组比较均明显增加(P<0.05)。免疫荧光下可见点状自噬体。结论饥饿可以诱导胃癌细胞MKN1发生自噬,Beclin-1与饥饿诱导胃癌细胞MKN1的自噬相关,为进一步研究Beclin-1在胃癌细胞自噬中的作用机制奠定了基础。
关键词〔〕微管相关蛋白1轻链3B;自噬;Beclin-1;饥饿;胃癌
1沈阳医学院机能教研室
2沈阳医学院2008级医学检验专业学生
第一作者:沈薇(1976-),女,博士,副教授,主要从事肿瘤生物学行为及其分子机制研究。
胃癌的发生发展是一个多因素和多阶段演化的过程,目前认为可能与自噬和凋亡有关〔1〕。自噬作用是细胞在饥饿、能量缺乏等代谢压力下的一种生理过程,细胞内的蛋白、细胞器和胞质通过自噬作用被包裹、消化并降解成核苷、氨基酸和脂肪酸循环利用,合成新的大分子和ATP〔2〕。自噬作用不仅具有维持细胞稳态、促进细胞生存的作用,过度上调的自噬作用也可以引起细胞死亡,即“自噬性细胞死亡”,也称为Ⅱ型程序化细胞死亡。自噬作用的调节异常与肿瘤的发生、发展密切相关〔3,4〕,细胞自噬研究可能为恶性肿瘤的治疗提供更新的途径。Beclin-1是近十多年来才发现的与自噬相关的一种抑癌基因,它通过诱导自噬而抑制肿瘤的发生、发展〔5〕,但其在胃癌细胞自噬中的具体作用及其机制尚不清楚。本研究采用Earle′s平衡盐缓冲液(EBSS)代替培养液处理人胃癌细胞MKN1,免疫荧光及蛋白质印迹法检测自噬的标记蛋白微管相关蛋白1轻链3B(LC3B)和自噬相关蛋白Beclin-1表达的变化,以进一步探讨胃癌中自噬作用的分子机制。
1材料与方法
1.1材料胃癌细胞MKN1由本实验室保存。RPMI 1640培养基,10%胎牛血清(GFBS),EBSS均购自美国Gibco公司;AMV逆转录试剂盒购自美国Promega公司;兔抗人β-actin,Beclin-1单克隆抗体,羊抗人LC3B多克隆抗体,羊抗兔,驴抗羊辣根过氧化物酶IgG购自美国Santa Cruz Biotechnology公司;增强型化学发光试剂(ECL)购自美国Pierce公司;Trizol购自美国Invitrogen公司;SYBR Green I Master 购自日本Takara公司。
1.2细胞培养及处理MKN1细胞在含10%FBS的RPMI1640培养液中,37℃,5%CO2条件下培养。培养至70%~80%细胞融合,对数生长期时,实验组细胞以EBSS洗3遍后,以EBSS代替RPMI1640培养液培养(饥饿处理),并在12 h后收获细胞或进行相关检测。对照组细胞不施加处理因素。
1.3免疫荧光观察饥饿诱导后自噬体的形成MKN1细胞培养于24孔板内,孔内事先置入无菌盖玻片。取正常对照细胞和EBSS培养12 h的细胞,PBS洗2次,4%多聚甲醛固定,10%BSA(0.2% TritonX-100)封闭,加入一抗(LC3B,滴度为1∶1 000),常温下孵育2 h,PBS清洗3次,加入荧光标记二抗 Alexa488(Invitrogen 公司,1∶800)常温下孵育1 h,甘油封片。于荧光显微镜下观察。
1.4实时荧光定量RT-PCR 检测LC3B和Beclin-1 mRNA表达以Trizol法提取实验和对照组细胞总RNA,并用紫外分光光度计检测RNA浓度及纯度(A260/A280>1.9)。根据浓度取1 μg总RNA按照逆转录试剂盒操作说明将RNA逆转录为cDNA,进行PCR扩增。LC3B上游引物5′-CAACATGAGCGAGTTGGTCAAGA-3′,下游引物5′-ACTCACCATGCTGTGCTGGTTC-3′;Beclin-1上游引物5′-ATGCAGGTGAGCTTCGTGTG-3′,下游引物5′-CTGGGCTGTGCTAAGTAATGGA-3′;GAPDH上游引物5′-AACGGATTTGGTCGTATTGGG-3′,下游引物5′-TCGCTCCTGGAAGATGGTGAT-3′。以上引物由上海生工生物技术有限公司合成。在无菌去酶的Eppendorf 管中按照说明书将配置好的real-time PCR反应液(反应总体积为20 μl)离心数秒,混匀后迅速置入ABI7500荧光定量PCR仪中,反应条件:95℃ 30 s,95℃ 5 s,60℃ 34 s,34个循环,加溶解曲线。获得各样本待测基因的Ct值。采用2-ΔΔCt法进行试验数据处理,其结果代表各目的基因表达的相对定量,以对照组作为矫正样本。
1.5Western印迹检测LC3B和Beclin-1表达收集对照组、饥饿诱导组细胞总蛋白,将蛋白煮沸5 min 使其变性,取处理后的蛋白样品进行SDS-PAGE电泳并电转移至PVDF膜,5%脱脂奶粉室温封闭1 h,羊抗LC3B多克隆抗体(1∶500),兔抗Beclin-1单克隆抗体(1∶500)或兔抗β-actin抗体(1∶1 000)4℃孵育过夜,TBST洗膜,驴抗羊(1∶2 000)或羊抗兔(1∶2 000)辣根过氧化物酶IgG室温孵育1 h,TBST洗膜,暗室中加入ECL液孵育后,利用凝胶电泳成像系统进行读片分析。
2结果
2.1免疫荧光显示饥饿诱导后细胞内自噬体形成荧光显微镜下可见,对照组与实验组中细胞均显现绿色荧光。MKN1细胞经饥饿处理12 h后,多数细胞出现点状聚集的自噬体,而对照组无此现象(图1)。
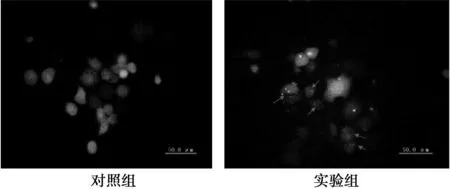
图1 饥饿诱导MKN1细胞后自噬体的形成(×40)
2.2实时定量RT-PCR检测LC3B和Beclin-1 mRNA表达LC3B mRNA 及Beclin-1 mRNA在饥饿处理12 h后表达(0.982±0.027,0.678±0.031)较对照组明显增高(0.463±0.019,0.135±0.056,P<0.05)。
2.3Western印迹检测LC3B和Beclin-1蛋白表达饥饿处理12 h后,与对照组LSC3BⅡ/LC3BⅠ灰度比值(0.13±0.32)比较,实验组LC3BⅡ/LC3BⅠ灰度比值(0.68±0.14)明显增加(P<0.05),Beclin-1蛋白表达在饥饿处理后显著增高〔(0.56±0.24)vs(1.67±0.15),P<0.05〕,见图2。
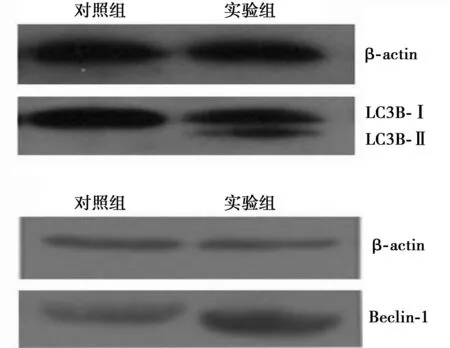
图2 饥饿处理后MKN1细胞LC3B和Beclin-1蛋白表达
3讨论
自噬作用参与多种生理过程中,在应激或饥饿状态下,可通过分解胞质内生物大分子为自身生存提供所需营养,也是清除细胞内长半衰期蛋白、衰老细胞器和受损结构的主要分子过程,还是细胞对不良环境的一种防御机制。自噬或调解自噬的某一环节出现问题都可能导致疾病发生。自噬作用的调节异常与肿瘤的发生、发展密切相关。实验研究发现,自噬作用是一种肿瘤抑制机制。有报道鼠的胰腺细胞经致癌物质诱导后,癌变前期细胞的自噬及降解能力增加,但当形成癌细胞后自噬能力下降〔4〕。多种肿瘤存在Beclin-1等自噬基因的缺陷,能够逃避自噬性死亡〔6〕。细胞自噬研究可能为恶性肿瘤的治疗提供更新的途径。
在哺乳动物细胞中,LC3B的修饰过程对自噬泡的形成起关键作用。LC3B前体形成后,在细胞质内通过加工生成胞质可溶性蛋白LC3B-I。当自噬发生的时候,自噬相关基因Atg7活化LC3B-Ⅰ,活化的LC3B-Ⅰ被转运至E2样酶Atg3,经其泛素化修饰后能够与自噬泡膜表面的磷脂酰乙醇胺(PE)结合成膜结合形式LC3B-Ⅱ〔7〕。由于LC3B-Ⅱ定位于自噬前体和自噬体,因此,目前认为LC3B-Ⅱ是自噬体的标志分子〔8〕。饥饿法是诱导细胞自噬的一种经典方法,在肝癌〔9〕、前列腺癌〔10〕等肿瘤细胞的自噬研究中均有应用。本研究显示细胞出现多个明亮的绿色荧光斑点,即自噬体形成,证实饥饿可以诱导胃癌细胞自噬发生。
另外,哺乳动物细胞发生自噬时,细胞内LC3B-Ⅰ向LC3B-Ⅱ的转化明显增加,LC3B-Ⅱ的含量与自噬泡的数量成正比关系〔11〕。因此,通过Western印迹法来检测LC3-Ⅰ到LC3-Ⅱ的转化,可以方便地判断自噬是被诱导还是被抑制,从而成为一种量化分析的依据。本研究发现,胃癌细胞经饥饿诱导后,除了可以观察到大量点状聚集的自噬体的形成,同时LC3-Ⅰ向LC3-Ⅱ的转化明显,这和Mizushima等〔12〕对GFP-LC3转基因鼠的研究结果相一致。
Beclin-1是哺乳动物参与自噬的特异性基因,其机制主要是与Ⅲ型磷脂酰肌醇激酶(PI3K)形成复合物参与募集胞质中含FYVE或PX基序的蛋白质,用于自噬体膜的形成,并引导其它自噬蛋白定位于自噬体膜〔5〕。细胞过度表达Beclin-1可以诱导细胞自噬。Beclin-1在人类卵巢癌中下降了75%,在乳腺癌中下降了50%〔13〕。有报道上调Beclin-1的表达能够增强顺铂诱导的胃癌细胞系MKN28凋亡,抑制Beclin-1的表达则削弱了顺铂的细胞毒作用〔14〕,提示Beclin-1的低表达在胃癌的发生、发展中起着一定的作用。本研究表明在胃癌细胞MKN1发生自噬的同时,能够检测到自噬相关蛋白Beclin-1表达的上调,说明Beclin-1表达上调与胃癌细胞自噬关系密切。
本研究成功采用饥饿法诱导胃癌细胞MKN1产生自噬,检测了自噬的标记蛋白LC3B的变化,建立了胃癌自噬的细胞模型。同时检测了自噬相关蛋白Beclin-1的表达变化,结果显示Beclin-1与胃癌细胞的自噬相关。这为进一步研究自噬对胃癌细胞活性的影响及自噬的作用机制奠定了基础。
参考文献4
1Xie B,Zhou J,Shu G,etal.Restoration of klotho gene expression induces apoptosis and autophagy in gastric cancer cells:tumor suppressive role of klotho in gastric cancer〔J〕.Cancer Cell Int,2013;13(1):2867-73.
2Kuma A,Hatano M,Matsui M,etal.The role of autophagy during the early neonatal starvation period〔J〕.Nature,2004;432(7020):1032-6.
3Yan CH,Liang ZQ,Gu ZL,etal.Contributions of autophagic and apoptotic mechanisms to CrTX2 induced death of K562 cells〔J〕.Toxicon,2006;47(5):521-30.
4Zheng HY,Zhang XY,Wang XF,etal.Autophagy enhances the aggressiveness of human colorectal cancer cells and their ability to adapt to apoptotic stimulus〔J〕.Cancer Biol Med,2012;9(2):105-10.
5Chen Z,Li Y,Zhang C,etal.Downregulation of Beclin 1 and impairment of autophagy in a small population of colorectal cancer〔J〕.Dig Dis Sci,2013;58(10):2887-94.
6Graf MR,Jia W,Johnson RS,etal.Autophagy and the functional roles of Atg5 and beclin-1 in the anti-tumor effects of 3beta and rostene 17alpha diol neurosteroid on malignant glioma cells〔J〕.Steroid Biochem Mol Biol,2009;115(35):137-45.
7Kroemer G,Marino G,Levine B.Autophagy and the integrated stress response〔J〕.Mol Cell,2010;40(2):280-93.
8Bellu AR,Kiel JA.Selective degradation of peroxisomes in yeasts〔J〕.Microsc Res Tech,2003;61(2):161-70.
9Tang H,Da L,Mao Y,etal.Hepatitis B virus X protein sensitizes cells to starvation-induced autophagy via up-regulation of beclin-1 expression〔J〕.Hepatology,2009;49(1):60-71.
10DiPaola RS,Dvorzhinski D,Thalasila A,etal.Therapeutic starvation and autophagy in prostate cancer:a new paradigm for targeting metabolism in cancer therapy〔J〕.Prostate,2008;68(16):1743-52.
11Tanida I,Ueno T,Kominami E.LC3 conjugation system in mammalian autophagy〔J〕.Int J Biochem Cell Biol,2004;36(12):2503-18.
12Mizushima N,Yamamoto A,Matsui M,etal.In vivo analysis of autophagy in response to nutrient starvation using transgenic mice expressing a fluorescent autophagosome marker〔J〕.Mol Biol Cell,2004;15(3):1101-11.
13Liang C,Feng P,Ku B,etal.Autophagic and tumor suppressor activity of a novel Beclin1-binding protein〔J〕.Nat Cell Biol,2006;8(7):688-99.
14Furuya D,Tsuji N,Yagihashi A,etal.Beclin-1 augmented cis-diamminedichloroplatinum induced apoptosis via enhancing Caspase-9 activity〔J〕.Exp Cell Res,2005;307(1):26-40.
〔2014-08-11修回〕
(编辑赵慧玲/曹梦园)
基金项目:辽宁省教育厅杰出青年学者成长计划(No.LJQ2012091);沈阳医学院优秀人才启动基金(No.20123041)
中图分类号〔〕R730.261〔
文献标识码〕A〔
文章编号〕1005-9202(2015)24-6994-03;doi:10.3969/j.issn.1005-9202.2015.24.012
