基于构件特征的内蒙古典型草原植物羊草个体地上生物量估算
2016-01-27胡静侯向阳萨茹拉郭丰辉丁勇
胡静,侯向阳,萨茹拉,郭丰辉,丁勇*
(1.中国农业科学院草原研究所,内蒙古 呼和浩特 010010;2.中国农业科学院研究生院,北京 100081;
3.农业部牧草资源与利用重点实验室,内蒙古 呼和浩特 010010)
基于构件特征的内蒙古典型草原植物羊草个体地上生物量估算
胡静1,2,侯向阳1,3,萨茹拉1,郭丰辉1,2,丁勇1,3*
(1.中国农业科学院草原研究所,内蒙古 呼和浩特 010010;2.中国农业科学院研究生院,北京 100081;
3.农业部牧草资源与利用重点实验室,内蒙古 呼和浩特 010010)
摘要:生物量是评价草原生态系统生产力的重要指标,草原主要植物个体地上生物量估算模型的建立,可为采用非刈割方法获取较准确的草原地面数据资料提供新方法。通过分析内蒙古典型草原植物羊草的主要构件特征(叶片数、叶长宽积、茎粗、茎高和株高)与其个体地上生物量之间的关系,比较并筛选出能较好预测羊草个体生物量变化的构件指标,采用Minitab软件的回归分析方法,分别利用6月、7月和8月采集的90株和总体270株羊草植物样本,建立羊草不同生长阶段(生长初期、生长旺盛期和生长末期)以及总体样本的个体地上生物量估算方程,并对实测值与模拟值进行t检验分析。结果表明,叶长宽积和株高相对于其他构件信息能够更好地描述羊草个体地上生物量,方程Y=-0.193+0.009X2+0.011X5(X2为叶长宽积,X5为株高,P<0.01,R2=0.854)为最优羊草个体生物量估算模型。
关键词:羊草;构件特征;个体地上生物量;典型草原
DOI:10.11686/cyxb2015053http://cyxb.lzu.edu.cn
胡静,侯向阳,萨茹拉,郭丰辉,丁勇. 基于构件特征的内蒙古典型草原植物羊草个体地上生物量估算. 草业学报, 2015, 24(8): 211-217.
Hu J, Hou X Y, Sa R L, Guo F H, Ding Y. Estimating individual above-ground biomass ofLeymuschinensisplants. Acta Prataculturae Sinica, 2015, 24(8): 211-217.
收稿日期:2015-01-27;改回日期:2015-03-18
基金项目:国家重点基础研究发展计划(973计划)项目(2014CB138805),国家自然科学基金项目(71103185,71311120089),中央公益性科研院所基本科研业务费项目(1610332015001),国家科技支撑计划项目(2012BAD13B07)和中国农业科学院科技创新工程(CAAS-ASTIP-IGR 2015-05)资助。
作者简介:胡静(1986-),女,内蒙古呼和浩特人,在读博士。E-mail:hu1129@sina.com
通讯作者*Corresponding author. E-mail: dingyong228@126.com
Estimating individual above-ground biomass ofLeymuschinensisplants
HU Jing1,2, HOU Xiang-Yang1,3, SA Ru-La1, GUO Feng-Hui1,2, DING Yong1,3*
1.InstituteofGrasslandResearchofChineseAcademyofAgriculturalScience,Hohhot010010,China; 2.GraduateSchoolofChineseAcademyofAgriculturalScience,Beijing100081,China; 3.KeyLaboratoryofGrasslandResourcesandUtilization,MinistryofAgriculture,Hohhot010010,China
Abstract:Biomass is an important component of grassland ecosystem productivity. Models able to describe the productivity of the main plant species in grassland offer the potential to estimate aboveground biomass without physical cutting. In this research the relationships among plant characteristics (leaf number, leaf length×leaf width, stem diameter, stem height and plant height) and individual above-ground biomass was analyzed, subsequently those characteristics able to best predict changes of individual above-ground biomass were selected for modelling. Ninety Leymus chinensis plants were sampled in June (early growing season), July (middle growing season) and August (late growing season), respectively, and total of 270 observations were used to build models able to estimate individual plant above-ground biomass. Equations were obtained using regression analysis using Minitab 16.0, and T-Test analysis was used to compare observed and predicted values. The results showed that leaf length×leaf width and plant height could describe individual above-ground biomass of L. chinensis plants; model Y=-0.193+0.009X2+0.011X5(X2indicated leaf length×leaf width,X5indicated plant height, P<0.01, R2=0.854).
Key words:Leymus chinensis; module characteristics; individual above-ground biomass; typical steppe
羊草(Leymuschinensis)草原是欧亚大陆草原的特有草地类型,集中分布于中国东北平原和内蒙古高原东部,是内蒙古典型草原植物群落的建群种或优势种[1]。羊草为赖草属根茎型多年生禾草,茎秆细嫩,叶量丰富,营养价值高,具有较好的耐盐碱、耐旱、耐践踏性,是一种优良的牧草资源[2]。生物量是评价草原生态系统生产力的重要指标,准确的测定草地植物生物量不仅是监测草原环境变化的依据,也是深入开展生态学、生理学及相关学科研究的基础[3]。羊草生物量变化是羊草草原生态系统群落结构优劣和功能高低的最直接体现,在大量的放牧[4-5]、刈割[6-7]、气候变化[8-9]、土壤营养元素变化[10-11]等相关研究中均有涉及。个体生物量是构成种群生物量和群落生产力的基础,个体地上生物量常与其主要构件的形态特征密切相关。与20世纪80年代相比,草原生产力持续衰减,严重地区可以下降60%~80%[12]。安渊等[13]研究发现草地退化后植被发生明显的矮化,同时伴随着生物量的减少。2000年之后,随着我国草原退化形势日趋严峻,很多研究以过度放牧为背景,阐述了羊草构件主要形态指标的变化,并明确指出,植物形态变化是生物量降低的主要原因,如王炜等[14-15]、Gallacher和Hill[16]的研究均认为在过度放牧下,草原植物性状整体表现为“小型化”(individual miniaturization),植株高度、构件生物量较未退化样地正常植株下降30%~80%,随着放牧胁迫增强,植物首先采取高度和生物量降低的适应策略。钟梦莹等[17]研究发现,在过度放牧干扰下羊草发生矮化现象,具体表现为株高明显降低,且叶片、根长、根节长减少,这也是导致羊草个体生物量明显下降的主要原因。李西良等[18]在呼伦贝尔草原的研究也表明,羊草茎高、株高、叶面积等对放牧响应极为敏感,放牧干扰会造成羊草株高和个体地上生物量显著降低。
可见,羊草植物形态特征与羊草地上生物量之间存在着明显的相关关系,但这种关系多为定性描述,仍缺乏对植物个体地上生物量与其主要易测形态指标的定量关系的研究。在科学研究和生产实践中,常以破坏性较大的样方法来进行草原生物量或生产力的地面调查和动态监测。草原主要植物个体地上生物量估算模型的建立,可为采用非刈割方法获取较准确的草原地面数据资料提供重要的方法和途径。本研究作为探索性研究,以内蒙古典型草原优势植物羊草作为研究对象,通过对植物叶片数、叶长宽积、茎高、茎粗、株高5个主要构件与个体地上生物量的分析,构建一系列多元线性回归模型,并进行模型优化和验证,希冀该研究能够为草原植物个体生物量和群落生产力的模拟研究提供新的思路和启示。
1材料与方法
1.1 研究区域概况
研究区域位于内蒙古锡林郭勒盟锡林浩特市朝克乌拉苏木中国农业科学院草原研究所草原生态系统保护和可持续利用研究与示范基地,地理位置系43°26′-44°08′ N,116°04′-117°05′ E,平均海拔1100 m。该区域属温带半干旱草原气候,年平均温度0.5~1℃,年平均降水350 mm左右,其中60%~80%的降水量集中在5-8月,年蒸发量1600~1800 mm,约为降水量的4~5倍。2014年生长季前半段较为湿润,但温度较低,对植物生长产生了一定影响,自7月下旬开始,发生较为严重的干旱(图1),对植物的生长产生较为严重的影响,致使部分植物提前枯黄。试验区自2007年开始禁牧,只作为割草利用,采样区植被类型为地带性典型草原植被,优势植物为羊草,主要植物包括大针茅(Stipagrandis)、糙隐子草(Cleistogenessquarrosa)、米氏冰草(Agropyronmichnoi)和黄囊苔草(Carexkorshinskyi)等,土壤为钙栗土。

图1 2014年研究区4月至10月降雨量与温度月动态(锡林浩特气象站)Fig.1 The dynamics of precipitation and temperature from April to October of study area in 2014
1.2 研究方法
试验设置3个采样区作为重复,分别在2014年6月12日(生长初期)、7月12日(生长旺盛期)和8月12日(生长末期)进行取样,随机选取单株羊草齐地刈割,每个采样区每月采集羊草样品40株,合计120株,全年共采集样品360株。采集后的样品带回实验室对株高、齐地刈割部位的茎粗、叶片数、叶长、叶宽和茎高等指标进行测量和计数,后将样品放入烘箱,65℃烘48 h至恒重,称重后获取地上个体生物量数据。
随机选取270株(6月、7月、8月各90株)羊草作为个体生物量估算模型的基础数据,通过Minitab软件建立个体生物量与各构件(叶片数、叶长宽积、茎粗、茎高、株高)数量特征的多元线性回归模型,并对羊草构件特征进行筛选以确定最优模型。本文中叶长宽积为每株羊草总叶片的累加值。通过剩余90株(6月、7月、8月各30株)羊草对模型进行验证,模型验证采用Independent-Samples T-Text法对模拟值和实测值进行比较。
2结果与分析
2.1 羊草地上个体生物量与主要构件的相关性分析
选取常规易测且具有相似变化规律的羊草主要构件作为估算模型的基础变量,研究假定各构件对羊草个体地上生物量的贡献值相同,由此可建立羊草个体地上生物量的多元线性估算模型为:
Y=β0+β1X1+β2X2+β3X3+β4X4+β5X5+μ
式中,X1为叶片数,X2为叶长宽积,X3为茎粗,X4为茎高,X5为株高,μ为随机变量,β为变量系数。
利用Minitab软件的相关分析功能,对羊草地上个体生物量、叶长宽积、茎粗、茎高和株高进行相关性分析,结果表明,羊草叶长宽积与羊草个体地上生物量的相关系数最高,是较适合的预测变量,二者间的R2(判定系数)为0.935,表明羊草地上个体生物量的变异性有93.50%可以通过叶生物量的增加或减少来解释。茎高和株高对羊草地上个体生物量的影响也不可忽视,相关系数也较高。通常习惯将相关系数较小的变量予以剔除,以确保模型的准确性。本研究中茎粗与羊草地上个体生物量的相关系数较低,而叶片数为离散变量,为了更好的拟合模型,将茎粗和叶片数这两个变量剔除(图2)。
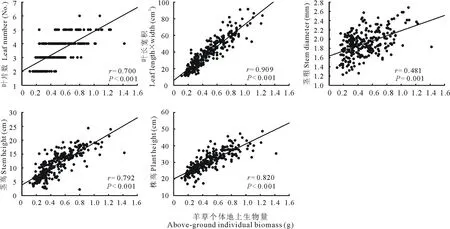
图2 羊草地上个体生物量与
因此羊草地上个体生物量估算模型修正为:
Y=β0+β2X2+β4X4+β5X5+μ
式中,X2为叶长宽积,X4为茎高,X5为株高,μ为随机变量,β为变量系数。
2.2 参数估算
通过Minitab中的回归分析进行参数估算,其中T是统计量的值,即描述样本特征的值,T值的绝对值越大,表示羊草个体地上生物量和各构件的关系越密切。P值为t检验(假设系数为0时,自变量和因变量无关)的判定值,P值小于0.05时,该假设不成立。因此T值越大,P值越小,则方程中的参数值越好。S表示残差的标准差,其值越小越好。R2是回归模型误差占总误差的百分比,取值越接近1,表明回归模型与数据吻合的越好。R2(adj)是经过Minitab回归分析后的调整值,R2和R2(adj)越接近,表明回归模型越可靠。
从表1可以看出,羊草的叶长宽积、茎高和株高能够较好的描述个体地上生物量,特别是在样本量较大(3月总样本)的情况下,方程参数的P值均为显著水平,S较小,R2较大,且R2和R2(adj)相接近,表明羊草个体生物量估算模型具有较好的拟合性(表1)。
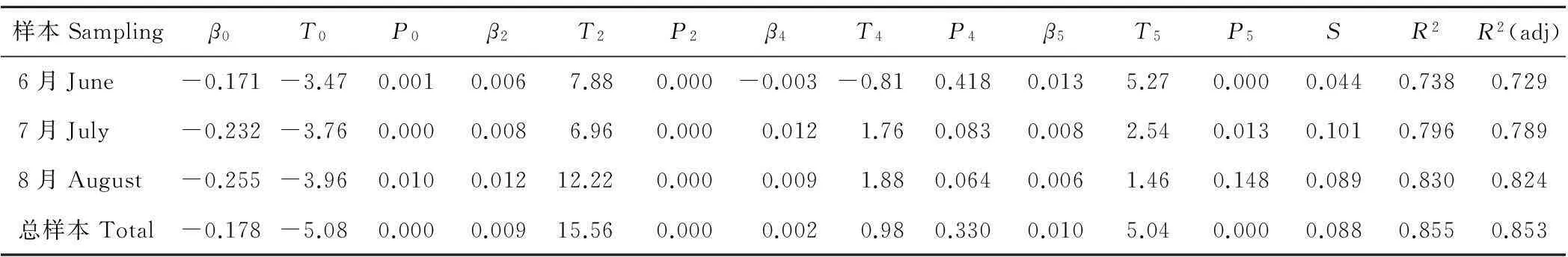
表1 羊草个体地上生物量估算模型
2.3 模型优化
方程参数的估算结果表明,6月、7月、8月和总样本的多元回归模型中,茎高参数β4的P值均大于0.05,t检验结果不显著,可以考虑剔除该项。模型中茎高和株高均是与高度有关的构件特征,相比较来说,羊草株高与个体地上生物量的相关关系要好于茎高,且在试验操作时,株高的测量较方便,因此剔除茎高对羊草个体地上生物量估算模型进行进一步优化。
优化后的羊草地上个体生物量估算模型修正为:
Y=β0+β2X2+β5X5+μ
式中,X2为叶长宽积,X5为株高,μ为随机变量,β为变量系数。
方程参数如表2所示。优化后各估算模型中所有参数的P值均达到显著水平,S和R2未有明显的波动,说明茎高的剔除有助于提高方程自变量系数的准确性。因此在本研究中,选择羊草叶长宽积和株高这两个构件特征值作为地上个体生物量的预测因子。6月至8月羊草个体地上生物量估算模型的R2呈逐渐增加趋势,这是由于羊草各构件生长趋于稳定,数据变异性逐渐减小所致,8月的构件特征数据能够更好的描述羊草个体地上生物量。同时加大样本量也有助于提高模型的准确性(表2)。

表2 羊草个体地上生物量估算模型优化
2.4 模型验证
通过剩余的90株羊草样品(6月、7月、8月各30株)对以上模型进行验证,以t检验分析对羊草个体地上生物量的实测值和模拟值进行比较,如表3所示,模拟值与实测值之间t检验结果均大于0.05,差异不显著,表明羊草个体地上生物量估算模型较可靠,可以对实际情况进行模拟(表3)。
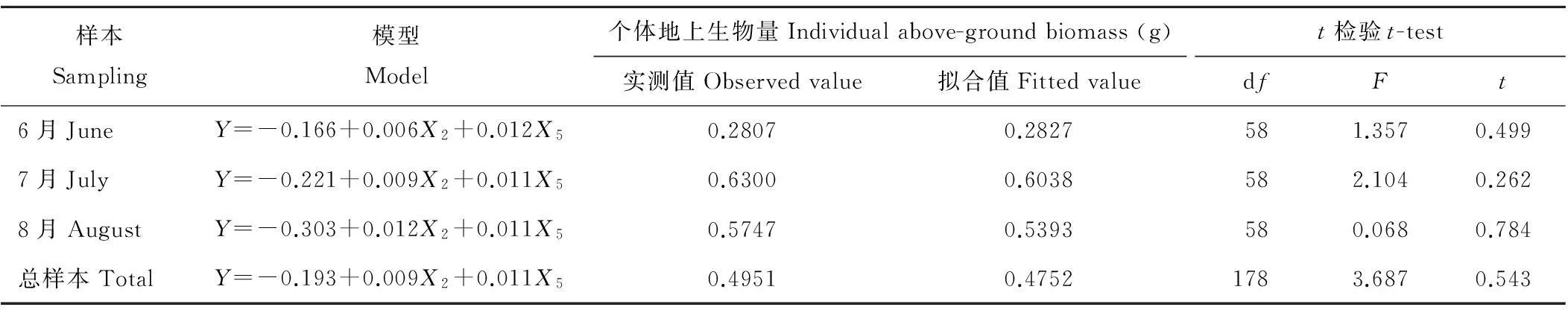
表3 羊草个体地上生物量估算模型验证
3讨论
本研究所获取的羊草主要构件数量特征在不同的生长阶段均呈现出了相同的变化趋势,表现为6月生长初期<7月生长旺盛期>8月生长末期,这与植物体内含水量随着气候发生波动有关[19-20],同时也可能是因为羊草在抽穗后,将更多的营养投入到生殖构件中,营养构件中的投入就会相应的减少所致[21]。因此在建立生物量估算模型时,要根据实际情况选取适当的取样时间。对比优化后羊草地上个体生物量估算模型的判定系数R2,可知8月的样品可以得到较好的地上个体生物量估算模型。如果为连续取样,则样品量较大时方程的拟合性好。通过本研究,我们认为应该进行多年的监测取样,通过对大量样本的研究,能够更清晰地刻画出羊草植物主要构件特征在生长季不同阶段的变化趋势和规律,以避免极端气候事件对研究结果的影响。
对羊草构件特征的研究表明,植物构件与其生物量之间存在着一定的相关关系。从羊草地上个体生物量估算模型建立的过程中可以看出,并不是构件叠加越多,得到的方程拟合性越好,有的构件对于个体生物量的作用存在冗余现象,剔除不但不影响模型的准确性,反而会使模型变得简化和优化。另外,从植物构件特征可以继续对模型进行扩展,如羊草叶长宽积可以用来计算叶面积,李亚军等[22]通过分析羊草叶面积和叶长宽积之间的相关关系,并与数字图像法相比较,得出羊草叶面积的计算公式为:A=k(L×W),k=0.6555,式中,A代表叶面积,k为叶长和叶宽乘积的修正系数,L为叶长,W为叶宽。在此次取样中虽未对叶面积进行测定,但在未来的研究中可以将叶面积作为变量参与模型的构建,并结合光谱技术[23],将单株的植物模型扩展到群落水平。本研究为多元线性回归模型的建立过程提供了一种筛选适合因子方式,不仅适用于生物量模型,也适用于其他多元线性回归模型的优化。
研究植物个体地上生物量模型模拟,具有重要的实践价值。草原植物多样性丰富,每种植物主要构件形态特征与个体生物量间是否存在普遍的相关性仍值得深入研究。目前植株的形态研究在农作物方面较多,并建立了多种作物生长模型[24],而关于草原主要植物的相关研究还较少。植物模型的研究已趋于多维化和可视化,但在未知领域基础数据的研究仍必不可少。在以往的草地监测和测产中,传统的样方法具有一定的破坏性,加之草原生态系统的异质性,往往使动态监测的结果存在着一定的误差,即使通过多点重复,也难以使误差消除。通过模型对植物生物量进行估算,一方面可以实现保护性测定,另一方面,通过定点、定样和定株监测估产,可有效提高测定结果的准确性。
4结论
内蒙古典型草原植物羊草叶长宽积和株高相对于叶片数、茎高和茎粗能够更好地描述其个体地上生物量的变化;通过Minitab软件的建模和模型优化处理,方程Y=-0.193+0.009X2+0.011X5(X2为叶长宽积,X5为株高,P<0.01,R2=0.854)为最优羊草个体生物量估算模型。
References:
[1]Pan Q M, Bai Y F, Han X G,etal. Effects of nitrogen additionson aLeymuschinensispopulation in typical steppe of Inner Mongolia. Acta Phytoecologica Sinica, 2005, 29(2):311-317.
[2]Li L H, Wang Q B, Bai Y F,etal. Soil respiration of aLeymuschinensisgrassland stand in the Xilin river basin as affected by over-grazing and mate. Acta Phytoecologica Sinica, 2000, 24(6):680-686.
[3]Dong X Y, Fu H, Li X D,etal. Effects on plant biomass and CNP contents of plants in grazed and fenced steppe grasslands of the Loess Plateau. Acta Prataculturae Sinica, 2010, 19(2):175-182.
[4]Wang R Z, Ripley E A. Effects of grazing on aLeymuschinensisgrassland on the Songnen plain of north-eastern China. Journal of Arid Environments, 1997, 36(2):307-318.
[5]Wang Y H, He X Y, Zhou G S. Study on the responses ofLeymuschinensissteppe to grazing in Songnen plain. Acta Agrestia Sinica, 2002, 10(1):45-49.
[6]Baoyin T G T, Li Y H, Bao Q H,etal. Effects of mowing regimes and climate variability on hay production ofLeymuschinensisgrassland in northern China. The Rangeland Journal, 2014, 36(6):593-600.
[7]He F, Li X L, Wan L Q. Influence of rainfall in growing season and elipping intensity on aboveground biomass ofLeymuschinensissteppe. Pratacultural Science, 2009, 26(4):28-32.
[8]Xu Z Z, Zhou G S. Combined effects of water stress and high temperature on photosynthesis, nitrogen metabolism and lipid peroxidation of a perennial grassLeymuschinensis. Planta, 2006, 224(5):1080-1090.
[9]Wang R Z, Gao Q. Climate-driven changes in shoot density and shoot biomass inLeymuschinensis(Poaceae) on the North-east China Transect (NECT). Global Ecology and Biogeography, 2003, 12(3):249-259.
[10]Ma L, Yuan S, Guo C,etal. Carbon and nitrogen dynamics of nativeLeymuschinensisgrasslands along a 1000 km longitudinal precipitation gradient in northeastern China. Biogeosciences, 2014, 11(24):7097-7106.
[11]He N P, Yu Q, Wu L,etal. Carbon and nitrogen store and storage potential as affected by land-use in aLeymuschinensisgrassland of northern China. Soil Biology and Biochemistry, 2008, 40(12):2952-2959.
[12]Han G D, Jiao S Y, Ao D,etal. Effects of plant species diversity and productivity under different stocking rates in theStipabrevifloraGriseb. desert steppe. Acta Ecologica Sinica, 2007, 27(1):182-188.
[13]An Y, Li B, Yang C,etal. Influence of grazing rate on population structure ofStipagrandis. Acta Phytoecologica Sinica, 2002, 26(2):163-169.
[14]Wang W, Liang C Z, Liu Z L,etal. Analysis of the plant individual, behaviouring the degradation and restoring succession in steppe community. Acta Phytoecologica Sinica, 2000, 24(3):268-274.
[15]Wang W, Liang C Z. Mechanism of degradation succession inLeymuschinensis+Stipagrandissteppe community. Acta Phytoecologica Sinica, 2000, 24(4):468-472.
[16]Gallacher D, Hill J. Effects of camel grazing on the ecology of small perennial plants in the Dubai (UAE) inland desert. Journal of Arid Environments, 2006, 66(4):738-750.
[17]Zhong M Y, Fan Q L, Zhang Y J,etal. Morphological plasticity and the biomass allocation models of miniaturizedLeymuschinensis. Acta Agrestia Sinica, 2013, 21(2):260-264.
[18]Li X L, Hou X Y, Wu X H,etal. Plastic responses of stem and leaf functional traits inLeymuschinensisto long-term grazing in a meadow steppe. Acta Phytoecologica Sinica, 2014, 38(5):440-451.
[19]Wullschleger S D, Hanson P J, Tschaplinski T J. Whole-plant water flux in understory red maple exposed to altered precipitation regimes. Tree Physiology, 1998, 18(2):71-79.
[20]Savi T, Andri S, Nardini A. Impact of different green roof layering on plant water status and drought survival. Ecological Engineering, 2013, 57(8):188-196.
[21]Liu P Y, Zhang Q L, Yang Y F. Biomass structure and quantitative relationship models of modules in clonal population ofPuccinilliachinampoensisin Songnen plain. Chinese Journal of Applied Ecology, 2004, 15(4):543-548.
[22]Li Y J, Ji S Q, Zhang S W,etal. The determination method of leaf area onLeymuschinensis. Journal of Anhui Agricultural Sciences, 2009, 37(15):6819-6851.
[23]Lv X D, Wang J G, Sun Q Z,etal. Research on the hyperspectral remote sensing estimation models for the fresh yield of alfalfa grassland. Acta Prataculturae Sinica, 2014, 23(1):84-91.
[24]Lai Q, Li Q F, Mo R G,etal. A study on factors affecting moisture content measurement and ratio of forage fresh weight to dry weight. Chinese Journal of Grassland, 2008, 30(4):73-77.
参考文献:
[1]潘庆民, 白永飞, 韩兴国, 等. 氮素对内蒙古典型草原羊草种群的影响. 植物生态学报, 2005, 29(2):311-317.
[2]李凌浩, 王其兵, 白永飞, 等. 锡林河流域羊草草原群落土壤呼吸及其影响因子的研究. 植物生态学报, 2000, 24(6):680-686.
[3]董晓玉, 傅华, 李旭东, 等. 放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响. 草业学报, 2010, 19(2):175-182.
[5]王玉辉, 何兴元, 周广胜. 放牧强度对羊草草原的影响. 草地学报, 2002, 10(1):45-49.
[7]何峰, 李向林, 万里强. 生长季降水量和刈割强度对羊草群落地上生物量的影响. 草业科学, 2009, 26(4):28-32.
[12]韩国栋, 焦树英, 敖登, 等. 短花针茅草原不同载畜率对植物多样性和草地生产力的影响. 生态学报, 2007, 27(1):182-188.
[13]安渊, 李博, 杨持, 等. 不同放牧率对大针茅种群结构的影响. 植物生态学报, 2002, 26(2):163-169.
[14]王炜, 梁存柱, 刘钟龄, 等. 草原群落退化与恢复演替中的植物个体行为分析. 植物生态学报, 2000, 24(3):268-274.
[15]王炜, 梁存柱. 羊草+大针茅草原群落退化演替机理的研究. 植物生态学报, 2000, 24(4):468-472.
[17]钟梦莹, 樊青丽, 张亚军, 等. 矮化羊草形态可塑性及生物量分配模式研究. 草地学报, 2013, 21(2):260-264.
[18]李西良, 侯向阳, 吴新宏, 等. 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应. 植物生态学报, 2014, 38(5):440-451.
[21]刘佩勇, 张庆灵, 杨允菲. 松嫩平原朝鲜碱茅无性系种群构件生物量结构及相关模型分析. 应用生态学报, 2004, 15(4):543-548.
[22]李亚军, 纪澍琴, 张守伟, 等. 羊草叶面积的测定方法. 安徽农业科学, 2009, 37(15):6819-6851.
[23]吕小东, 王建光, 孙启忠, 等. 苜蓿人工草地高光谱遥感估产模型的研究. 草业学报, 2014, 23(1):84-91.
[24]来强, 李青丰, 莫日根, 等. 影响牧草含水量测定以及牧草干鲜比的主要因素. 中国草地学报, 2008, 30(4):73-77.
