蛇床子素对转染APP595/596基因的SH-SY5Y细胞的保护作用
2016-01-11教亚男,姚璎珈,孔亮等
蛇床子素对转染APP595/596基因的SH-SY5Y细胞的保护作用*
教亚男,姚璎珈,孔亮,李少恒,陶震宇,闫宇辉,杨静娴△
(辽宁中医药大学,辽宁 大连116600)
[摘要]目的: 研究蛇床子素对转染APP595/596基因的SH-SY5Y细胞的作用,以探讨其可能的作用机制。方法: 体外培养SH-SY5Y细胞,并转染APP595/596基因,体外构建研究Aβ致病作用的细胞模型。采用CCK-8法检测细胞存活率;检测LDH评价细胞的损伤程度;流式细胞术检测细胞凋亡;用RT-PCR和Western blot法检测β-分泌酶(又称β位点APP裂解酶1,β-site APP cleaving enzyme 1,BACE 1)的mRNA及蛋白的表达;采用免疫荧光细胞化学和Western blot法检测Aβ的表达。结果: 蛇床子素对转染APP595/596基因的SH-SY5Y神经细胞有保护作用,可增加细胞的存活率,降低LDH的释放,抑制细胞的凋亡,减少BACE1的mRNA与蛋白的表达,抑制Aβ的生成。结论: 蛇床子素可保护转染APP595/596基因的SH-SY5Y神经细胞,其保护机制可能与减少BACE l的mRNA与蛋白表达有关。
[关键词]蛇床子素; APP595/596基因; β位点APP裂解酶1
[中图分类号]R329.2+5; R363[文献标志码]A
doi:10.3969/j.issn.1000-4718.2015.11.021
[文章编号]1000-4718(2015)11-2059-06
[收稿日期]2015-04-13[修回日期] 2015-08-28
[基金项目]*国家自然科学基金资助项目(No. 81260241)
通讯作者△Tel: 0453-7916015; E-mail: 1257067540@qq.com
Protective effect of osthole on SH-SY5Y cells transfected withAPP595/596 geneJIAO Ya-nan, YAO Ying-jia, KONG Liang, LI Shao-heng, TAO Zhen-yu, YAN Yu-hui, YANG Jing-xian
(LiaoningUniversityofTraditionalChineseMedicine,Dalian116600,China.E-mail:jingxianyang@yahoo.com)
ABSTRACT[]AIM: To explore the protective effect of osthole on the SH-SY5Y cells transfected with APP595/596 gene, and to investigate the molecular mechanism. METHODS: The SH-SY5Y cells were transfected with APP595/596 gene in vitro for establishing a cell model to study the pathogenic role of amyloid β-protein (Aβ). The cell viability was detected by CCK-8 assay. The release of lactate dehydrogenase (LDH) was determined by the colour reaction of diaphorase-INT. The cell apoptotic rate was analyzed by flow cytometry. The expression of β-site APP cleaving enzyme 1 (BACE1) at mRNA and protein levels was detected by RT-PCR and Western blot. The expression of Aβ was measured by the technique of immunofluorescence cytochemistry and Western blot. RESULTS: Treatment with osthole inhibited the LDH release, and increased the viability of the cells. The percentage of apoptotic cells was also significantly decreased. Osthole also inhibited the expression of BACE1 at mRNA and protein levels and the protein expression of Aβ. CONCLUSION: Osthole has protective effect on SH-SY5Y cells transfected with APP595/596 gene. The mechanism may be association with inhibiting the mRNA and protein expression of BACE1.
[KEY WORDS]Osthole;APP595/596 gene; β-site APP cleaving enzyme 1
阿尔茨海默病(Alzheimer’s disease,AD)是一种常见的致死性神经退行性疾病,多发于65岁以上的老人,是威胁人类健康和发展的最严峻的社会问题之一。然而却没有较好的治疗方案,所以找到有效治疗AD的药物对临床治疗AD,提高患者生存率有重要意义。AD的显著特征是淀粉样β-蛋白(amyloid β-protein,Aβ)的沉积[1]。越来越多的研究显示Aβ是AD形成与发展的关键因素,Aβ的异常聚集和沉积具有神经毒性[2],可引起神经细胞的凋亡,促进了AD的发生[3-4]。
蛇床子素(osthole)又名甲氧基欧芹素,是从多种伞形科植物如中药蛇床子、独活等提取分离出来的天然香豆素类化合物[5-6]。目前国内外研究表明蛇床子素有多种药理作用,包括抗炎、抗肿瘤、抗癌[7]、改善记忆等功效。本实验利用第595和596位氨基酸密码子发生并列突变(G→T和A→C)的淀粉样前体蛋白595/596(amyloid precursor protein 595/596,APP595/596)基因转染SH-SY5Y细胞,旨在探究蛇床子素对转染细胞的神经保护作用,进而探讨AD发病的机制及治疗方法。
材料和方法
1材料和仪器
SH-SY5Y细胞购于首都医科大学;DMEM/F12培养基、DMEM高糖培养基、胎牛血清(fetal bovine serum,FBS)、胰蛋白酶-EDTA消化液和青霉素/链霉素(penicillin and streptomycin,P/S)购于Gibco; APP595/596质粒、GFP质粒DNA以及慢病毒包装系统pLP1、pLP2、pLP/VSV-G(ViraPowerTMLentiviral Expression Systems)委托天津医科大学闫亚平教授前期构建;LipofectamineTM2000 购于Invitrogen;293T细胞由天津医科大学闫亚平教授惠赠;大肠杆菌DH5α感受态细胞和无内毒素质粒小提试剂盒购于Omega;蛇床子素购自中国药品生物制品鉴定所(分子量244.29,纯度>99.0%);细胞增殖与活性检测试剂盒CCK-8购于Dojindo;Annexin V-Light 650/PI细胞凋亡试剂盒购于沈阳万类生物公司;二甲基亚砜(dimethyl sulfoxide,DMSO) 购于Amresco;LDH试剂盒购于南京建成公司;TRIzol购于Invitrogen;cDNA合成试剂盒和PCR Master Mix试剂均购于Fermentas;兔抗人Aβ1-42单克隆抗体(I 抗)购于Bioss;Cy3 标记羊抗兔免疫球蛋白G(IgG II 抗)购于Jackson;其它试剂皆为国产分析纯。
荧光生物显微镜(Nikon);超低温冰箱(青岛海尔);CO2培养箱(NUAIRE);酶标仪(深圳迈瑞);流式细胞仪(BD);紫外-可见光分光光度计(上海元析);PCR仪(杭州朗基);凝胶成像系统(UVP)。
2方法
2.1SH-SY5Y细胞与293细胞的培养SH-SY5Y细胞与293细胞分别常规培养于DMEM/F12与高糖DMEM完全培养基(10% FBS、1% P/S)中,于37 ℃、5%的CO2培养箱中培养,3 d换1次液,待细胞达到80%~90%融合度时进行传代,选取对数生长期的细胞进行实验。
2.2慢病毒包装转染SH-SY5Y细胞建立AD的细胞模型将APP595/596质粒转化大肠杆菌DH5α感受态细胞,并接种在Ampr抗性平板上,16 h后挑取阳性克隆,置于LB 液体培养基中37 ℃振荡培养18 h,采用无内毒素质粒小提试剂盒进行提取质粒,再将APP595/596质粒/GFP质粒与慢病毒系统pLP1、pLP2、pLP/VSV-G以15 μg:6.5 μg:2.5 μg:3.5 μg比例用Lipofectamine 2000介导转染293T包装细胞,对照组转染GFP[8]。培养293T细胞,24 h后荧光显微镜下观察2组细胞都呈现绿色荧光。分别收集转染后72 h的上述293T细胞培养液上清(含病毒),3 000 r/min离心20 min去除细胞碎片,0.22 μm滤膜过滤除菌,100 kD超滤管5 000 r/min离心1 h得到浓缩液并检测病毒滴度;用浓缩的病毒上清转染SH-SY5Y细胞;转染3 d后用RT-PCR与Western blot检测APP的 mRNA及蛋白的表达量[9]。
2.3细胞分组对照组为稳定转染GFP的SH-SY5Y细胞;模型组为稳定转染APP595/596基因的SH-SY5Y细胞。给药组将APP595/596基因稳定转染的SH-SY5Y细胞分别给予终浓度10 μmol/L、50 μmol/L、100 μmol/L的蛇床子素处理24 h后进行实验。
2.4CCK-8法检测细胞活力将细胞以1×108/L的密度接种于96孔板中,按上述分组每组3个复孔,培养终点时每孔加入10 μL的CCK-8溶液,将培养板在培养箱内孵育4 h。用酶标仪在450 nm处检测吸光度(A)值。
2.5乳酸脱氢酶(lactate dehydrogenase,LDH)释放量的检测细胞以1×108/L接种于96孔板,进行分组,每组取3复孔,培养至终点取上清液,根据试剂盒说明书进行操作,用紫外-可见光分光光度计于510 nm下测定吸光度值,检测各组细胞LDH释放量[10]。
2.6流式细胞术检测细胞凋亡细胞接种于6孔板,分组处理后,用不含EDTA的胰酶消化,PBS清洗2遍后,加入500 μL的binding buffer重悬细胞;加入5 μL Annexin V-Light 650混匀后,加入5 μL PI混匀,室温避光反应15 min,用流式细胞仪检测细胞凋亡率[11]。
2.7RT-PCR实验细胞接种于6孔板内,分组后,弃去培养液,用PBS洗3次,向每孔加入1 mL的 TRIzol,吹打数次,转移到Eppendorf管中,冰浴5 min,再加入0.2 mL氯仿,盖紧后剧烈振摇15 s,冰浴5 min,12 000 r/min、4 ℃离心10 min;取上层水相,加入0.5 mL异丙醇,轻微混合,冰浴10 min,12 000 r/min、4 ℃离心10 min;弃去上清,用75%乙醇混悬,再7 500 r/min、4 ℃离心5 min;弃去上清将沉淀自然风干后溶于0.01% DEPC水,吹打,提取出总RNA。用紫外分光光度计检测260 nm和280 nm处吸光度,A260/A280在1.8~2.0范围内样品质量合格;浓度=A260×稀释倍数×40 mg/L。逆转录合成cDNA(25 μL体系): 取各组RNA 3 μg样品,按Revert AidTMFirst Strand cDNA Synthesis Kit所示比例配制成RT反应体系,65 ℃ 5 min;42 ℃ 60 min,70 ℃ 5 min。合成cDNA,-80 ℃冻存备用。进行PCR反应(50 μL体系):按PCR Master Mix Kit所示比例配制成PCR反应体系,样品DNA取2.5 μL。各引物序列见表 1。反应条件:95 ℃ 2 min;95 ℃ 30 s,5 ℃ 30 s,72 ℃ 1 min,72 ℃ 10 min, 35个循环,再进行琼脂糖凝胶电泳,用ImageJ进行光密度扫描分析。
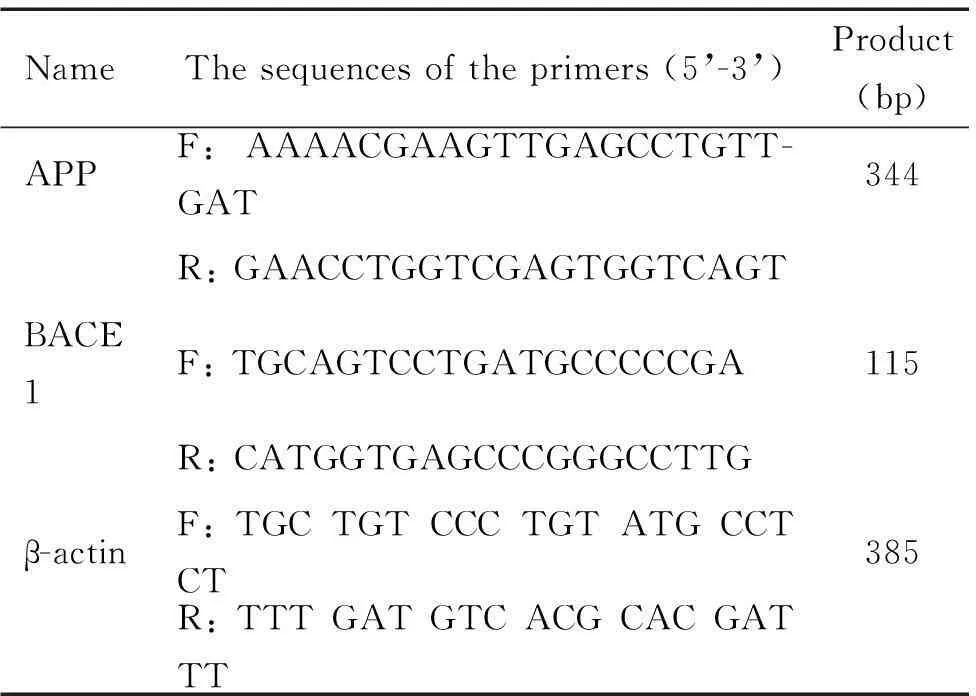
表1 APP、BACE1和β-actin的引物序列
F: forward; R: reverse.
2.8免疫荧光细胞化学法检测Aβ的表达将细胞接种于96孔板中,每组取3复孔,以4%多聚甲醛固定30 min,再用0.1% Triton X-100透化20 min,加入 I 抗(1∶100),于4 ℃过夜,次日用PBS洗后,加入Cy3标记特异 II 抗(1∶200),室温放置1 h后再用PBS洗涤后,加入DAPI染色15 min后,PBS洗涤后,用倒置荧光显微镜观察并用ImageJ软件对各组细胞中Aβ的荧光强度进行扫描和定量,平行独立实验3次。
2.9Western blot 检测β位点APP裂解酶1(β-site APP cleaving enzyme 1, BACE1)及Aβ蛋白的表达细胞裂解液提取总蛋白,蛋白变性,SDS-PAGE分离蛋白,转膜和封闭。分别与兔抗人BACE1抗体(1∶200)、兔抗人Aβ抗体(1∶200)和羊抗兔过氧化物酶标记 II抗(1∶500)作用。化学发光剂反应,X线片曝光,显影,定影。进行扫描分析。
3统计学处理
采用GraphPad Prism 5软件完成实验数据的统计处理,数据以均数±标准差(mean±SD)表示,两组间比较采用双侧t检验,多组间比较采用单因素方差分析,而后用Bonferroni校正的t检验进行多组均数间的两两比较。以P<0.05为差异有统计学意义。
结果
1获得转染APP595/596基因稳定表达APP的SH-SY5Y细胞
SH-SY5Y细胞转染APP595/596基因后,可在荧光显微镜下观察GFP的表达情况,用RT-PCR法与Western bolt检测APP的表达情况,结果显示模型组与对照组相对比APP呈高表达,证明成功获得稳定表达APP的SH-SY5Y细胞,见图1。
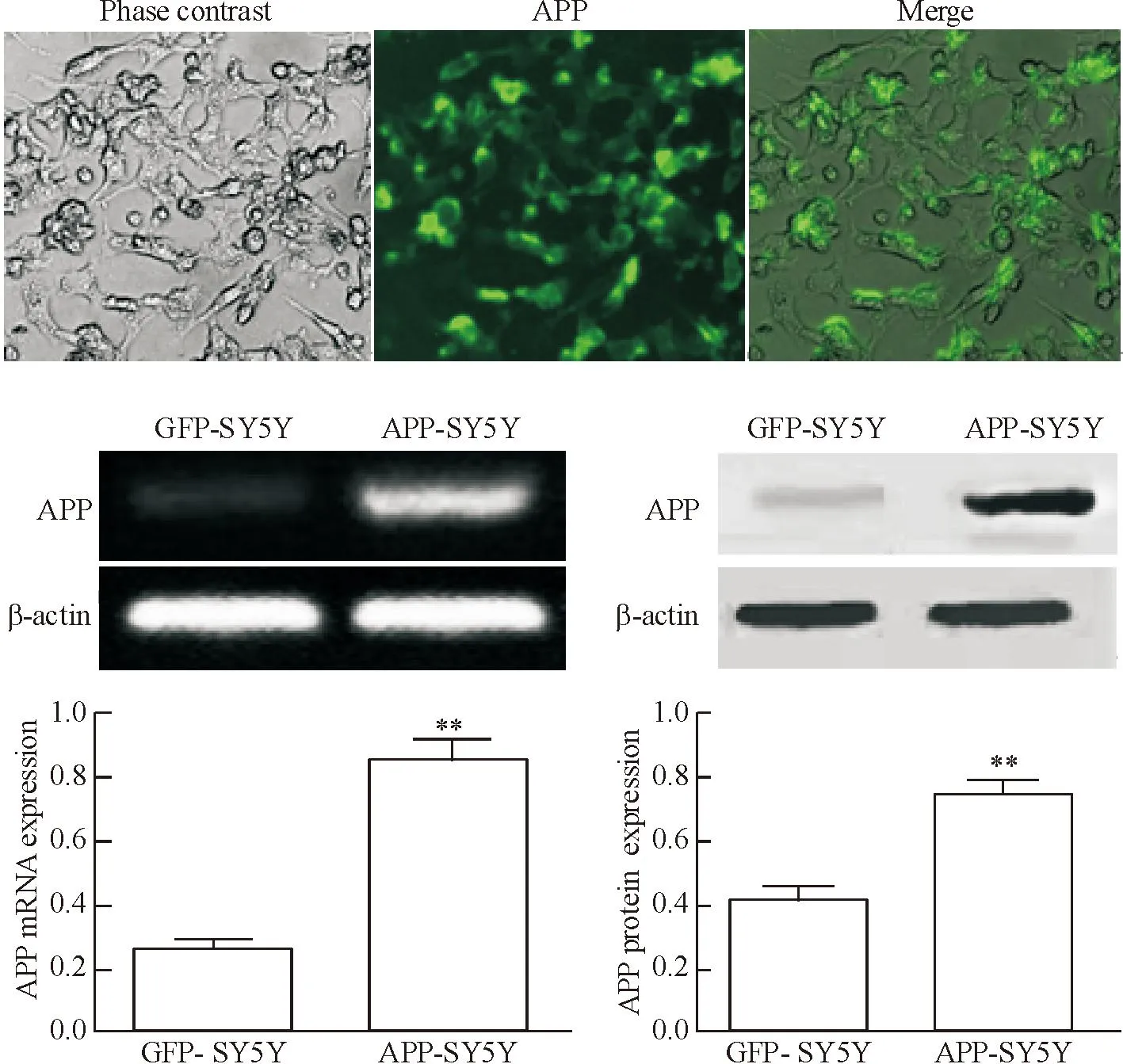
Figure 1. APP was highly expressed inAPP595/596-transfected SH-SY5Y cells. The scale bar=25 μm. Mean±SD.n=3.**P<0.01vsGFP group.
图1APP在SH-SY5Y细胞中高表达
2蛇床子素对转染APP595/596基因的SH-SY5Y细胞的保护作用
CCK-8法检测结果显示,与对照组相比,转染APP595/596基因的模型组细胞存活率显著下降(P<0.01),而不同浓度的蛇床子素可使得细胞活力上升,其中经50 μmol/L蛇床子素处理的细胞活力最高。上述结果显示50 μmol/L蛇床子素为保护转染APP595/596基因的SH-SY5Y细胞的最佳浓度。故以下各项实验中给药组所用的蛇床子素浓度均为50 μmol/L,见图2。
3蛇床子素对转染APP595/596基因的SH-SY5Y细胞中LDH释放量的影响
模型组细胞的LDH释放量明显增加,其释放量几近对照组的2倍,差异具有统计学意义(P<0.01)。与模型组相比,给药组细胞的LDH释放量明显减少 (P<0.01),见图2。
4蛇床子素抑制转染APP595/596基因的SH-SY5Y细胞凋亡
流式细胞术结果显示,模型组凋亡细胞的百分比为27.9%,与对照组的5.2% 相比差异有统计学意义;而给药组的凋亡细胞百分比为16.4%,显著低于模型组,说明蛇床子素能有效抑制转染APP595/596基因的SH-SY5Y细胞的凋亡,见图2。

Figure 2.The effect of osthole (Ost, 50 μmol/L) on the SH-SY5Y cells transfected withAPP595/596 gene. Mean±SD.n=3.**P<0.01vsGFP group;##P<0.01vsAPP group.
图2蛇床子素对转染APP595/596基因的SH-SY5Y细胞的作用
5蛇床子素下调转染APP595/596基因的SH-SY5Y细胞的Aβ表达
Aβ免疫荧光细胞化学法检测结果可见,各组细胞浆中均有Aβ的表达,而模型组细胞的红色荧光强度较对照组明显减弱,给予蛇床子素后可显著减弱模型组细胞的荧光强度。Western blot的结果也可见模型组细胞中Aβ蛋白的表达量明显高于对照组,而给药组则可显著降低其表达量,说明蛇床子素可显著抑制Aβ蛋白的表达水平,见图3。
6蛇床子素可抑制转染APP595/596基因的SH-SY5Y细胞中BACE1的 mRNA与蛋白表达
RT-PCR结果显示,模型组细胞BACE1的mRNA表达量显著上升,给予蛇床子素后BACE1的mRNA表达量降低,与模型组相比差异有统计学意义(P<0.01),说明蛇床子素可抑制BACE1的mRNA表达。Western blot法的检测结果可见,模型组细胞BACE1蛋白表达量与对照组细胞相比显著上升,给药组可降低BACE1蛋白的的表达,说明蛇床子素能抑制BACE1蛋白的表达,见图4。
讨论
阿尔茨海默病是严重威胁人类晚年生活质量的主要疾病之一,其淀粉样致病假说已被广泛接受,研究人员认为Aβ的异常聚集和沉积是AD患者脑内病变的重要原因之一,其脑内水平升高可引起神经毒性,包括直接毒性作用和增强、放大各种伤害性毒性作用[12],进而诱导神经细胞的凋亡,最终导致AD的发生。现而今已有不少治疗AD的方式开始研究,但是目前仍缺乏有效的防治措施。

Figure 3.Osthole (Ost, 50 μmol/L) inhibited Aβ levels inAPP595/596-transfected SH-SY5Y cells(×40). Mean±SD.n=3.**P<0.01vsGFP group;##P<0.01vsAPP group.
图3蛇床子素可抑制转染APP595/596基因的SH-SY5Y细胞中Aβ的表达
蛇床子素是独活、蛇床子等中药中的活性成分,为天然香豆素衍生物,作为一种脂溶性物质,解离度小,易通过血脑屏障,几乎没有毒性作用。已有研究发现蛇床子素能通过调控MAPK蛋白表达对大鼠皮层神经元内氧和葡萄糖剥夺有作用[13],更有研究发现蛇床子素能改善大鼠记忆功能[14]。这些研究都提示蛇床子素可能具有治疗和改善AD相关的临床症状及病理变化的潜在功效。
1992年Citron等[15]发现APP基因的第595和第596位氨基酸密码子发生并列突变(G-T、A-C),致使Aβ的分泌量显著升高。因此本实验采用APP595/596基因转染SH-SY5Y细胞成功在体外构建研究Aβ致病作用的细胞模型,Aβ的表达显著升高,SH-SY5Y细胞呈现与Aβ损伤有关的特征:CCK-8法显示细胞的存活率降低,LDH表达量增多,流式细胞术结果发现细胞凋亡率升高。在本研究中我们发现给予蛇床子素后对以上损伤变化具有显著改善。为了进一步研究蛇床子素对神经细胞保护作用的机制,本实验又针对Aβ形成途径进行研究。采用RT-PCR法与Western blot法,检测BACE1的mRNA与蛋白的表达。BACE 1是一种膜结合的天冬氨酸蛋白酶,是裂解APP产生Aβ的限速酶,更有研究人员发现在体内敲除BACE1基因的小鼠,其Aβ的产生会减少[16],为BACE1作为AD治疗靶点的可能性提供了实验依据。而本实验发现蛇床子素可降低BACE1的 mRNA及蛋白水平,说明蛇床子素可作为BACE1抑制剂,能减少APP的淀粉样代谢途径,进而抑制了Aβ蛋白的表达,而关于蛇床子素抑制BACE1的机制,仍需要在今后的研究中做进一步探讨。
综上所述,蛇床子素具有神经保护作用,能通过减少BACE1表达进而抑制Aβ的产生,提高细胞的存活率,降低损伤程度,减少细胞凋亡,在治疗AD相关新药的开发中具有重要的应用价值。
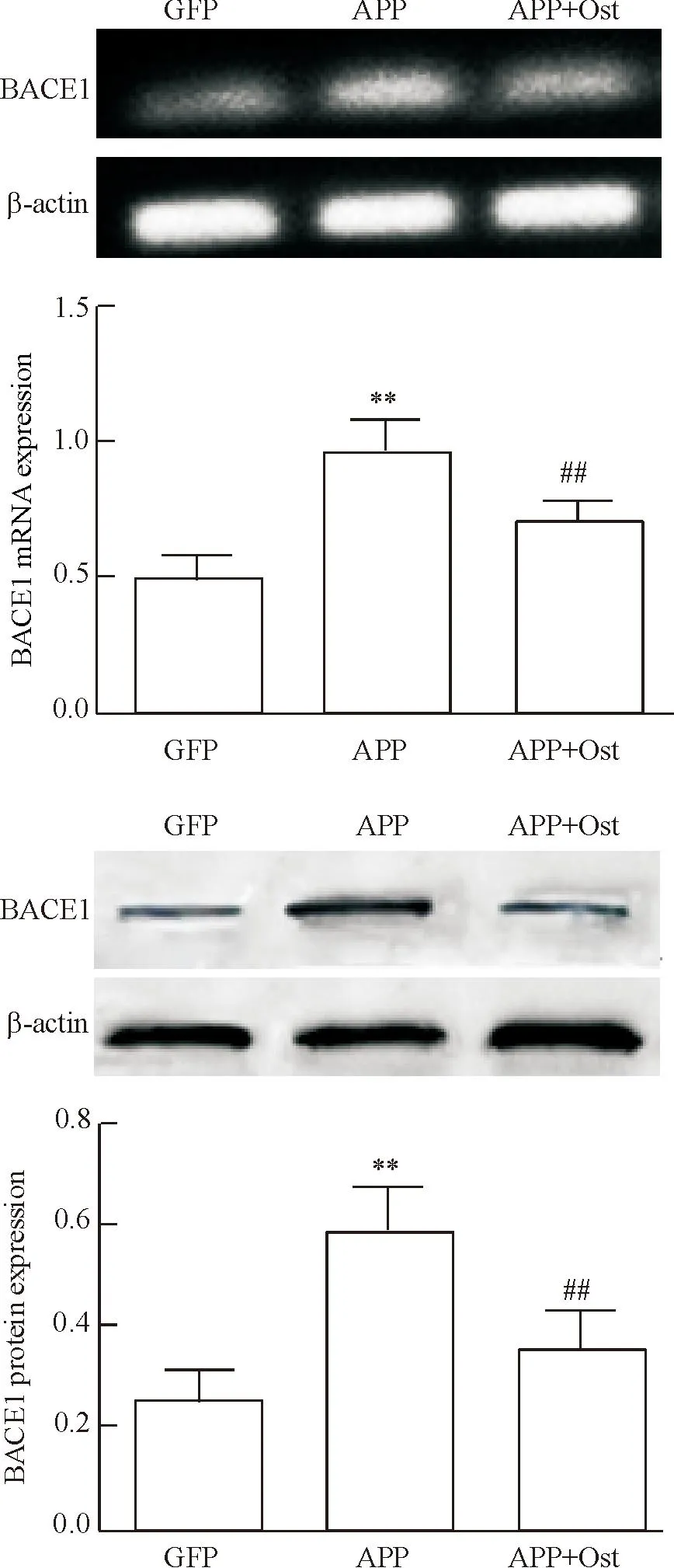
Figure 4.Osthole (Ost, 50 μmol/L) decreased the expression of BACE1 at mRNA and protein levels in the SH-SY5Y cells transfected withAPP595/596. Mean±SD.n=3.**P<0.01vsGFP group;##P<0.01vsAPP group.
图4蛇床子素能降低转染APP595/596的SH-SY5Y细胞中BACE1的mRNA和蛋白表达
[参考文献]
[1]Kukreja L, Kujoth GC, Prolla TA, et al. Increased mtDNA mutations with aging promotes amyloid accumulation and brain atrophy in the APP/Ld transgenic mouse model of Alzheimer’s disease[J]. Mol Neurodegener, 2014, 9:16.
[2]Kowall NW, Beal MF, Busciglio J, et al. Aninvivomodel for the neurodegenerative effects of β-amyloid and protection by substance P[J]. Proc Natl Acad Sci U S A, 1991, 88(16):7247-7251.
[3]Butterfield DA. Amyloid β-peptide (1-42)-induced oxidative stress and neurotoxicity: implications for neurodegeneration in Alzheimer’s disease brain. A review[J]. Free Radical Res, 2002, 36(12):1307-1313.
[4]Zhu X, Chen C, Ye D, et al. Diammonium glycyrrhi-zinate upregulates PGC-1α and protects against Aβ1-42-induced neurotoxicity[J]. PLoS One, 2012,7(4):e35823.
[5]周则卫,沈秀. 蛇床子素药理活性的研究概况[J]. 中国新药杂志, 2006, 15(20):1726-1730.
[6]毛佳蕾,钱晓萍,刘宝瑞. 蛇床子素的提取及其体外抗肿瘤活性的研究进展[J]. 现代肿瘤医学, 2014, 22(3):726-728.
[7]Lin YC, Lin JC, Hung CM, et al. Osthole inhibits insulin-like growth factor-1-induced epithelial to mesenchymal transition via the inhibition of PI3K/Akt signaling pathway in human brain cancer cells[J]. Agric Food Chem, 2014, 62(22):5061-5071.
[8]Yan Y, Ding X, Li K, et al. CNS-specific therapy for ongoing EAE by silencing IL-17 pathway in astrocytes[J]. Mol Ther, 2012, 20(7):1338-1348.
[9]Yang J, Yan Y, Xia Y, et al. Neurotrophin 3 transduction augments remyelinating and immunomodulatory capacity of neural stem cells[J]. Mol Ther, 2014, 22(2):440-450.
[10]Cao BY, Yang YP, Luo WF, et al. Paeoniflorin, a potent natural compound, protects PC12 cells from MPP+and acidic damage via autophagic pathway[J]. J Ethnopharmacol, 2010, 131(1):122-129.
[11]王梦瑶, 王苏美, 左应林, 等. 姜黄素衍生物 T83 对同源不同辐射抗性鼻咽癌细胞凋亡的作用[J]. 中国病理生理杂志, 2013, 29(4): 654-659.
[12]刘洪玲,郭素,王辉明,等. 海康灵复方对Aβ(1-40)诱导SH-SY5Y细胞损伤的保护作用[J]. 中国医院药学杂志,2013, 33(17):1405-1408.
[13]Chen T, Liu W, Chao X, et al. Neuroprotective effect of osthole against oxygen and glucose deprivation in rat cortical neurons: involvement of 49 mitogen-activated protein kinase pathway[J]. Neuroscience, 2011, 183(6):203-211.
[14]沈丽霞,张丹参,张力,等. 蛇床子素对学习记忆的影响及其机制分析[J]. 药学学报,1999, 34(6):405-409.
[15]Citron M, Oltersdorf T, Haass C, et al. Mutation of the β-amyloid precursor protein in familial Alzheimer’s disease increases β-protein production[J]. Nature, 1992, 360(6405):672-674.
[16]谢兆宏. 长非编码RNA通过调节BACE1表达介导β淀粉样蛋白生成的机制研究[D]. 济南:山东大学, 2012.
(责任编辑: 陈妙玲, 罗森)

