猪胃肠道谷氨酸代谢及其对机体健康的影响
2016-01-07
猪胃肠道谷氨酸代谢及其对机体健康的影响
黄 博1,2胡佳宇1,2吴苗苗1,2肖 昊1,2张 军1,2谭碧娥1∗印遇龙1∗
(1.中国科学院亚热带农业生态研究所,中国科学院亚热带农业生态过程重点实验室,湖南省畜禽健康养殖工程技术中心,农业部中南动物营养与饲料科学观测实验站,长沙410125;
2.中国科学院大学,北京100049)
E⁃mail:664135003@qq.com
摘 要:谷氨酸是组成蛋白质或肽的结构氨基酸之一。胃和小肠能吸收游离谷氨酸,结肠上皮细胞及结肠微生物也能利用肠腔内的谷氨酸。食物中的谷氨酸在胃肠道被大量代谢,为肠道完整性和功能维持提供了大量三磷酸腺苷(ATP),是胃肠道的主要能量来源。此外,谷氨酸是谷氨酰胺等其他氨基酸以及一些生物活性分子的前体,发挥着营养、免疫调节和抗氧化应激等生理作用。本文综述谷氨酸在猪胃肠道的代谢机制及其对机体健康的影响。
关键词:猪;谷氨酸;代谢;胃肠道;健康
谷氨酸是一种酸性氨基酸,约占膳食蛋白的10%~20%,在生物体内的蛋白质代谢过程中占有重要地位。谷氨酸在小肠细胞中被大量代谢,只有小部分的谷氨酸可从肠腔进入肠系膜静脉,是肠细胞的主要能源物质。谷氨酸、谷氨酰胺和天冬氨酸的代谢为肠道完整性和功能维持提供了大量的三磷酸腺苷(adenosine triphosphate,ATP)[1],且与甘氨酸一起调节神经功能[2]。由于血浆谷氨酸、谷氨酰胺和天冬氨酸浓度过高会引起神经毒性[3],它们在小肠的大量代谢对于组织生长也是必需的。因此,谷氨酸的代谢对于维持机体健康具有重要意义。
1 谷氨酸在胃内的转运和代谢
越来越多的研究表明,有小部分谷氨酸能够在胃内被吸收代谢[4]。Dougls等通过往仔猪胃和十二指肠灌注13C标记测定谷氨酸的代谢率,发现胃内13C标记的谷氨酸主要被氧化成二氧化碳(CO2),然而,当灌注3倍正常谷氨酸时,与十二指肠灌注相比,通过胃灌注方式谷氨酸在胃肠道的吸收率显著提高表明谷氨酸以游离形式并在高剂量添加的情况下,一部分谷氨酸可以在胃内消化代谢。除了进行谷氨酸的消化代谢外,胃上皮细胞有许多谷氨酸转运载体表达,Zhang等[5]研究发现在饲粮中补偿谷氨酸可以提高胃肠道谷氨酸信号受体和谷氨酸转运体的表达量。由此可见,胃部也是谷氨酸感应和调节消化功能的重要位点。一些研究发现谷氨酸可以通过胆碱型神经促进胃底部收缩和血液流动,也可以通过谷氨酸受体促进胃酸分泌,从而控制胃肠道的消化功能[6]。上述研究表明表明胃黏膜具有转运谷氨酸进入血液循环的能力,同时也是也是谷氨酸感应和调节消化功能的位点[7]。
2 谷氨酸在小肠内的转运和代谢
尽管一直认为氨基酸主要在肝脏中代谢,但肠道也是许多氨基酸如谷氨酰胺、谷氨酸、天冬氨酸代谢的重要场所。饲粮中仅有5%谷氨酸和天冬氨酸、30%~33%的谷氨酰胺及60%~64%的脯氨酸进入门脉循环[8]。谷氨酸在小肠的代谢大多发生在肠上皮细胞。谷氨酸主要通过高亲和力的X⁃AG系统和低亲和力的B0系统从肠腔转运进入肠上皮细胞。X⁃AG系统转运谷氨酸和天冬氨酸,包括谷氨酸—天冬氨酸转运载体-1(glutamate/as⁃partate transporter⁃1,GLAST⁃1)、谷氨酸转运载体-1(glutamate transporter⁃1,GLT⁃1)、兴奋性氨基酸载体(excitatory amino⁃acid carrier,EAAC)⁃1、EAAC⁃4和EAAC⁃5[9-11]。肠道内主要的谷氨酸转运载体为EAAC⁃1,在整个小肠膜的刷状缘顶端表达。但GLAST⁃1和GLT⁃1在不同胃细胞中表达,而在小肠表达量较少。最新研究表明,饲粮中补充谷氨酸可以显著提高空肠EAAT⁃1、EAAT⁃2、EAAT⁃3和EAAT⁃4表达量,从而最大化吸收利用肠腔中的谷氨酸[5]。
氧化是谷氨酸在肠道上皮细胞的主要代谢方式。谷氨酸在肠上皮细胞内被氧化,首先与草酰乙酸通过转氨基作用产生α-酮戊二酸和天冬氨酸,α-酮戊二酸进入线粒体通过三羧酸循环产生还原型辅酶烟酰胺腺嘌呤二核苷酸(nicotinamide⁃adenine dinucleotide,reduced,NADH)和黄素腺嘌呤二核苷酸(flavin adenine dinucleotide,reduced,FADH2),用于线粒体ATP的合成(图1)[12]。同时,谷氨酸转氨基作用产生的天冬氨酸也能进入线粒体通过三羧酸循环被氧化成为肠上皮细胞的另一种能源物质[13]。

图1 谷氨酸在小肠内的代谢途径Fig.1 Glutamate metabolism pathways in the small intestine[12]
大量的谷氨酰胺在体内代谢,包括水解并通过谷氨酰胺酶作用生成谷氨酸和氨,小肠肠腔内细菌和肠上皮细胞利用谷氨酸进一步生成谷胱甘肽、脯氨酸、鸟氨酸和精氨酸[11,14-15],或经过代谢生成CO2或葡萄糖和以尿素或氨排泄出体外(图2)[1,3]。当谷氨酸和谷氨酰胺同时存在于上皮细胞中时,谷氨酸能够抑制谷氨酰胺的利用和氧化[5]。同时,谷氨酰胺降解生成的谷氨酸,经吡咯啉-5-羧酸合成酶(pyrroline⁃5⁃carboxylate syn⁃thase,P5CS)催化可转化为谷氨酸-γ-半醛,产生吡咯啉羧酸(pyrroline⁃5⁃carboxylate,P5C),降解生成脯氨酸。绝大多数脯氨酸在体内的合成是通过谷氨酸/P5CS途径(图3)[16],P5CS仅在肠道表达,所以脯氨酸净合成部位是小肠[17]。
3 谷氨酸在大肠内的代谢
众所周知,小肠和大肠的肠腔环境有很大的不同,大肠的肠腔中有大量的细菌和细菌代谢物,其中有一部分微生物代谢产物是能源物质。此外,在刚出生的小段时间内,大肠中几乎没有氨基酸从肠腔转运到血液中。在这种情况下,氨基酸必须由动脉血进入结肠细胞,结肠上皮细胞利用血液中的谷氨酰胺作为氧化底物,谷氨酰胺首先被线粒体中的谷氨酰胺酶转化成谷氨酸和氨,然后通过转氨作用转化成α-酮戊二酸,进入三羧酸循环[12]。尽管食物中的蛋白质在小肠中充分消化水解成氨基酸和寡肽被吸收,但也有大量的含氮混合物可以通过回盲连接进入大肠,被结肠微生物代谢产物水解成氨基酸和多肽被吸收[14]。在大肠肠腔,蛋白质和多肽水解释放的谷氨酸及氨,在谷氨酰胺合成酶的作用下转化生成谷氨酰胺,大肠黏膜有着较小肠黏膜更高的谷氨酰胺合成酶活性,将结肠肠腔中的谷氨酸和氨转化成谷氨酰胺,以防止结肠肠腔中较高的氨浓度抑制短链脂肪酸在结肠上皮细胞的氧化[15]。由此可见,谷氨酸在结肠上皮细胞的供能过程中具有极其重要的作用。然而,结肠细胞对谷氨酸的利用并不仅仅局限在能量代谢上。结肠细胞中存在丰富的还原型谷胱甘肽合成和转运活性,作为合成谷胱甘肽的前体物质,谷氨酸对于维持结肠上皮细胞还原性、抵抗各种氧化应激具有重要的作用[18]。此外,还有研究发现,在体外分离的结肠细胞氨基酸代谢过程中,谷氨酸可以被转化成天冬氨酸、丙氨酸等其他氨基酸,从而提高饲粮中氨基酸的利用率[19]。
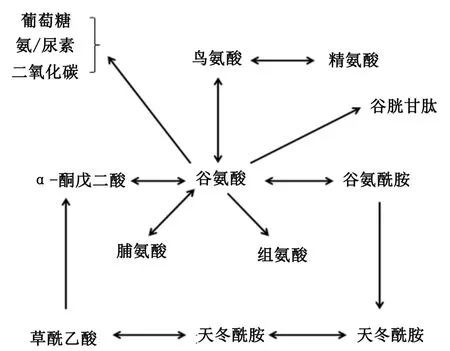
图2 谷氨酸代谢简图Fig.2 Sketch of metabolism of glutamate[1,3]
4 谷氨酸代谢对猪肠道和机体健康的影响
近年来,越来越多的研究表明,胃肠道组织主要通过氨基酸代谢来获取能量而不是葡萄糖或脂肪酸。谷氨酰胺能抑制葡萄糖在肠上皮细胞中的氧化,尽管葡萄糖是一种重要的能源物质,但是在胃肠道中葡萄糖氧化比例小于谷氨酸或谷氨酰胺。Stoll等[20]以猪为试验对象发现饲粮中90%谷氨酸在肠道中被代谢,其中50%转化成CO2,是消化道主要的能源物质。这说明谷氨酰胺和谷氨酸在消化道首先被被线粒体氧化为肠细胞增殖、分化以及维持胃肠道完整性和胃肠道功能提供能量,而食物中的大部分的葡萄糖被用于机体其他组织器官代谢,保障猪机体健康生长所需要的足够能源物质。
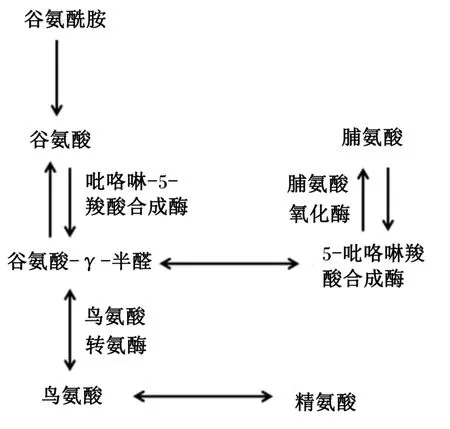
图3 脯氨酸合成和利用途径Fig.3 Pathways of proline synthesis and utilization[17]
谷氨酸尤其是谷氨酸钠的长期食用一直以来被认为对幼畜及婴儿具有神经毒性的风险,还会引起肥胖等疾病[21-22]。但是,也有研究发现,过量的谷氨酸(4倍正常量)摄入对血液中谷氨酸浓度没有显著影响[23-24],只有极端高剂量的谷氨酸才会带来神经毒性[25]。正常情况下,谷氨酸进入消化道被氧化成CO2或者转化成其他非必需氨基酸。研究发现,当饲粮中谷氨酸摄入量增加到3倍时,肠内谷氨酰胺、天冬氨酸和鸟氨酸分别显著增加4.8、4.0、2.7倍;小肠吸收的脯氨酸、精氨酸和其他支链氨基酸等也显著增加[26]。Feng等[27]研究发现在饲粮中添加谷氨酸钠可以通过增加消化道氨基酸转运载体的表达量从而提高空肠氨基酸的吸收量。这些研究说明,提高饲粮中谷氨酸的水平能够减少或节省消化道对其他氨基酸的代谢,从而提高饲粮中氨基酸的利用率。因此,在养猪生产中,我们可以通过在饲粮中适当补充谷氨酸以达到节约氨基酸的效益,以提高营养物质的利用率。此外,谷氨酸是合成谷胱甘肽的前体物质,而谷胱甘肽是肠道上皮细胞内主要的抗氧化物质,在氧化应激情况下对于维持肠道健康和稳态极其重要[28]。Wu等[29]、吴苗苗等[30]研究发
现,在饲粮中补充谷氨酸可显著降低仔猪氧化应激水平,缓解由呕吐毒素引起的肠道损伤,从而维持机体健康。这表明谷氨酸在肠道中的代谢对于肠道健康和缓解肠道疾病具有重要作用。新生儿和幼畜容易出现胃和十二指肠运动功能障碍,由于胃排空慢而出现饲喂不耐受症状,在养猪生产上常表现为断奶仔猪腹泻,在仔猪断奶前后饲粮中添加谷氨酸可以缓解断奶应激导致的肠绒毛萎缩和仔猪腹泻[31]。Akiba等[32]研究发现十二指肠肠上皮细胞内的pH和黏液层厚度与肠腔中L-谷氨酸浓度(0.1~10 mmol/L)成正比,L-谷氨酸的这种影响受十二指肠中辣椒素和消炎药敏感通路的调节,辣椒素敏感传入神经和环氧化酶活性在其中的发挥重要作用。Akiba等[33]还研究发现,L-谷氨酸对细胞内pH和黏液层厚度的影响受代谢型谷氨酸受体4(metabotropic glutamate recep⁃tor4,mGluR4)拮抗剂抑制,而mGluR4激动剂具有L-谷氨酸类似的作用效果,钙传感受体(calci⁃um sensing receptor,CaSR)激动剂可以通过增加黏液层厚度和十二指肠碳酸氢盐分泌酸化细胞。由此可见,L-谷氨酸通过传入神经和激活多种谷氨酸受体如mGluR4、CaSR等调节肠上皮细胞pH和肠腔黏液层厚度,通过提高肠道黏膜防御能力来保护肠黏膜,防止进入十二指肠中的胃酸造成的黏膜损伤。
5 小 结
目前研究已证实饲粮中的谷氨酸绝大部分在胃肠道消化代谢,谷氨酸不仅是肠道重要的能源物质,也是合成其他功能性氨基酸前体,在饲粮中添加谷氨酸可以促进其他氨基酸的合成,达到节约氨基酸效应,从而提高营养利用率。近年来谷氨酸在维持机体健康中发挥的作用受到了越来越多的关注。但是,不同生长阶段或不同生理状态下谷氨酸的安全添加剂量以及谷氨酸的营养生理作用及其机制需要进一步研究。以期明确胃肠道谷氨酸代谢更多的生理作用,为谷氨酸在猪生产中的应用提供理论依据。
参考文献:
[1]WATFORD M.Glutamine metabolism and function in relation to proline synthesis and the safety of gluta⁃mine and proline supplementation[J].Journal of Nu⁃trition,2008,138(10):2003S-2007S.
[2]WU G Y.Functional Amino acids in growth,reproduc⁃tion,and health[J].Advances in Nutrition,2010,1(1):31-37.
[3]WU G Y.Amino acids:metabolism,functions,and nu⁃trition[J].Amino Acids,2009,37(1):1-17.
[4]TORII K,UNEYAMA H,NAKAMURA E.Physiolog⁃ical roles of dietary glutamate signaling via gut⁃brain axis due to efficient digestion and absorption[J].Jour⁃nal of Gastroenterology,2013,48(4):442-451.
[5]ZHANG J,YIN Y L,SHU X G,et al.Oral administra⁃tion of MSG increases expression of glutamate recep⁃tors and transporters in the gastrointestinal tract of young piglets[J].Amino Acids,2013,45(5):1169-1177.
[6]BURRIN D G,STOLL B.Metabolic fate and function of dietary glutamate in the gut[J].American Journal of Clinical Nutrition,2009,90(3):850S-856S.
[7]HOWELL J A,MATTHEWS A D,WELBOURNE T C,et al.Content of ileal EAAC1 and hepatic GLT⁃1 high⁃affinity glutamate transporters is increased in growing vs.nongrowing lambs,paralleling increased tissue D⁃and L⁃glutamate,plasma glutamine,and ala⁃nine concentrations[J].Journal of Animal Science,2003,81(4):1030-1039.
[8]WU G,BAZER F W,BURGHARDT R C,et al.Im⁃pacts of amino acid nutrition on pregnancy outcome in pigs:mechanisms and implications for swine produc⁃tion[J].Journal of Animal Science,2010,88(13S):E195-E204.
[9]FAN M Z,MATTHEWS J C,ETIENNE N M P,et al.Expression of apical membrane L⁃glutamate transport⁃ers in neonatal porcine epithelial cells along the small intestinal crypt⁃villus axis[J].American Journal of Physiology:Gastrointestinal and Liver Physiology,2004,287(2):G385-G398.
[10]BEART P M,O’SHEA R D.Transporters for L⁃gluta⁃mate:an update on their molecular pharmacology and pathological involvement[J].British Journal of Phar⁃macology,2007,150(1):5-17.
[11]KANAI Y,HEDIGER M A.The glutamate and neutral amino acid transporter family:physiological and phar⁃macological implications[J].European Journal of Pharmacology,2003,479(1/2/3):237-247.
[12]BLACHIER F,BOUTRY C,COS C,et al.Metabolism and functions of L⁃glutamate in the epithelial cells of the small and large intestines[J].American Journal of Clinical Nutrition,2009,90(3):814S-821S.
[13]BLACHIER F,GUIHOT⁃JOUBREL G,VAUGELADE P,et al.Portal hyper glutamate mia after dietary supple⁃mentation with monosodium glutamate in pigs[J].Di⁃gestion,1999,60(4):349-357.
[14]BLACHIER F,MARIOTTI F,HUNEAU J F,et al.Effects of amino acid⁃derived luminal metabolites on the colonic epithelium and physiopathological conse⁃quences[J].Amino acids,2007,33(4):547-562.
[15]CREMIN J D JR,FITCH M D,FLEMING S E.Glu⁃cose alleviates ammonia⁃induced inhibition of short⁃chain fatty acid metabolism in rat colonic epithelial cells[J].American Journal of Physiology:Gastrointes⁃tinal and Liver Physiology,2003,285(1):G105-G114.
[16]BERTOLO R F P,BRUNTON J A,PENCHARZ P B,et al.Arginine,ornithine,and proline interconver⁃sion is dependent on small intestinal metabolism in neonatal pigs[J].American Journal of Physiology:En⁃docrinology and Metabolism,2003,284(5):E915-E922.
[17]FUJITA T,YANAGA K.Association between gluta⁃mine extraction and release of citrulline and glycine by the human small intestine[J].Life Sciences,2007,80(20):1846-1850.
[18]ROEDIGER W E W,BABIDGE W.Human colono⁃cyte detoxification[J].Gut,1997,41(6):731-734.
[19]DARCY⁃VRILLON B,MOREL M T,CHERBUY C,et al.Metabolic characteristics of pig colonocytes after adaptation to a high fiber diet[J].Journal of Nutri⁃tion,1993,123(2):234-243.
[20]STOLL B,BURRIN D G,HENRY J,et al.Substrate oxidation by the portal drained viscera of fed piglets[J].American Journal of Physiology:Endocrinology and Metabolism,1999,277(1):E168-E175.
[21]HERMANUSSEN M,TRESGUERRES J A F.How much glutamate is toxic in paediatric parenteral nutri⁃tion?[J].Acta Paediatrica,2005,94(1):16-19.
[22]HERMANUSSEN M,TRESGUERRES J A F.Does the thrifty phenotype result from chronic glutamate in⁃toxication?A hypothesis[J].Journal of Perinatal Medicine,2003,31(6):489-495.
[23]HERMANUSSEN M,GARCÍA A P,SUNDER M,et al.Obesity,voracity,and short stature:the impact of glutamate on the regulation of appetite[J].European Journal of Clinical Nutrition,2006,60(1):25-31.
[24]HAŸS S P,ORDONEZ J M,BURRIN D G,et al.Di⁃etary glutamate is almost entirely removed in its first pass through the splanchnic bed in premature infants[J].Pediatric Research,2007,62(3):353-356.
[25]JANECZKO M J,STOLL B,CHANG X Y,et al.Ex⁃tensive gut metabolism limits the intestinal absorption of excessive supplemental dietary glutamate loads in infant pigs[J].Journal of Nutrition,2007,137(11):2384-2390.
[26]LAMBERT B D,FILIP R,STOLL B,et al.First⁃pass metabolism limits the intestinal absorption of enteral alpha⁃ketoglutarate in young pigs[J].Journal of Nutri⁃tion,2006,136(11):2779-2784.
[27]FENG Z M,ZHOU X L,WU F,et al.Both dietary supplementation with monosodium L⁃glutamate and fat modify circulating and tissue amino acid pools in growing pigs,but with little interactive effect[J].PLoS One,2014,9(1):e84533.
[28]REEDS P J,BURRIN DG,STOLL B,et al.Enteral glutamate is the preferential source for mucosal gluta⁃thione synthesis in fed piglets[J].American Journal of Physiology:Endocrinology and Metabolism,1997,273(2):E408-E415.
[29]WU M M,XIAO H,REN W K,et al.Therapeutic effects of glutamic acid in piglets challenged with de⁃oxynivalenol[J].PLoS One,2014,9(7):e100591.
[30]吴苗苗,肖昊,印遇龙,等.谷氨酸对脱氧雪腐镰刀菌烯醇刺激下的断奶仔猪生长性能、血常规及血清生化指标变化的干预作用[J].动物营养学报,2013,25(7):1587-1594.
[31]CABRERA R A,USRY J L,ARRELLANO C,et al.Effects of creep feeding and supplemental glutamine or glutamine plus glutamate(aminogut)on pre⁃and post⁃weaning growth performance and intestinal health of piglets[J].Journal of Animal Science and Biotech⁃nology,2013,4(1):29.
[32]AKIBA Y,WATANABE C,MIZUMORI M,et al.Lu⁃minal L⁃glutamate enhances duodenal mucosal defense mechanisms via multiple glutamate receptors in rats[J].American Journal of Physiology:Gastrointestinal and Liver Physiology,2009,297(4):G781-G791.
Metabolism of Glutamic Acid in Gastrointestinal Tract and Its Effects on Health of Pigs
HUANG Bo1,2HU Jiayu1,2WU Miaomiao1,2XIAO Hao1,2ZHANG Jun1,2TAN Bi’e1∗YIN Yulong1∗
(1.Scientific Observing and Experimental Station of Animal Nutrition and Feed Science in South⁃Central,Ministry of Agriculture,Hunan Provincial Engineering Research Center of Healthy Livestock and Poultry,Key Laboratory of Agro⁃Ecological Processes in Subtropical Region,Institute of Subtropical Agriculture,Chinese Academy of Sciences,Changsha 410125,China;
2.University of the Chinese Academy of Sciences,Beijing 100049,China)
(责任编辑 陈 燕)
E⁃mail:bietan@isa.ac.cn;YIN Yulong,professor,
E⁃mail:yinyulong@isa.ac.cn
Abstract:Glutamic acid is one of constituents of protein or peptide.The free glutamic acid can be absorbed by the stomach and small intestine and also can be utilized by colonic epithelial cells and its microbes.The most of glutamic acid in diets is metabolized in the gastrointestinal tract,which provides plenty of adenosine triphos⁃phate(ATP)to maintain the gut integrity and functions and is used as the main energy source of the gastroin⁃testinal tract.In addition,as the principal precursor of other amino acids and bioactive molecules,glutamic plays nutritional,immunoregulatory and anti⁃oxidative functions.In the present review,the metabolism of glu⁃tamic acid in the gastrointestinal tract and its effects on the health of pigs were summarized.[Chinese Journal of Animal Nutrition,2015,27(11):3326⁃3331]
Key words:pigs;glutamic acid;metabolism;gastrointestinal tract;health
Corresponding author∗s:TAN Bi’e,associate professor,
doi:10.3969/j.issn.1006⁃267x.2015.11.002
文章编号:1006⁃267X(2015)11⁃3326⁃06
文献标识码:A
中图分类号:S852.2;S828
作者简介:黄 博(1991—),女,湖南汩罗人,硕士研究生,从事单胃动物营养研究。
基金项目:国家自然科学基金(31110103909,31330075,31372326);中国科学院青年创新促进会(2014336)
收稿日期:2015-06-12
通信作者:∗谭碧娥,副研究员,
E⁃mail:bietan@isa.ac.cn;印遇龙,研究员,博士生导师,
E⁃mail:yinyulong@isa.ac.cn
