7个短枝木麻黄种源耐寒评价及综合分析
2015-12-20姜清彬马英姿仲崇禄
刘 芬,姜清彬,马英姿,仲崇禄,张 勇,陈 羽,陈 珍
(1.中南林业科技大学,湖南 长沙 410004;2.中国林科院热带林业研究所,广东 广州 510520)
7个短枝木麻黄种源耐寒评价及综合分析
刘 芬1,2,姜清彬2,马英姿1,仲崇禄2,张 勇2,陈 羽2,陈 珍2
(1.中南林业科技大学,湖南 长沙 410004;2.中国林科院热带林业研究所,广东 广州 510520)
对7个短枝木麻黄种源的离体小枝在人工气候箱中进行梯度低温处理, 在25 ℃,5 ℃,0 ℃,-5 ℃ 和-10 ℃温度下处理24 h,重复3次,每个种源的每个处理采集10株苗的枝条,测定了渗透率、丙二醛含量、脯氨酸含量、可溶性蛋白含量,可溶性糖含量等9个耐寒指标。通过半致死温度评价了7个种源的耐寒能力,并通过耐寒指标的特征结合相关分析对耐寒指标进行筛选。运用主成分分析、回归分析和聚类分析对耐寒指标进一步综合分析。结果表明:利用短枝木麻黄的半致死温度对7个种源进行排序,耐寒能力按种源号依次为18402>18268>18119>18267>18128>18015>18244。用逐步回归的方式建立短枝木麻黄的耐寒回归方程为Y=9.462-0.119Chl4-0.026SP1-0.039MDA1+0.051Chla3-0.008Chla2,(R2=1.000),估计精度大于98%。
短枝木麻黄;种源耐寒评价;低温胁迫;主成分分析;线性回归
短枝木麻黄Casuarina equisetifolia耐盐碱、抗风害、沙埋能力强,树干较通直,主根深长,有根瘤菌固氮,适应性强,能在滨海有机质低的沙土上迅速生长[1],是我国华南沿海防护林的重要树种,也是防风固沙农田防护林的先锋树种之一[2]。但是由于短枝木麻黄不耐寒,严重限制了其栽种范围,且近年来的极端气温频繁出现,对耐寒的短枝木麻黄的选育得尤为重要。木麻黄的耐寒研究,主要集中在耐寒性比较及对耐寒独立指标的分析[3-5],未对耐寒指标进行综合评价。本文通过结合主成分分析、线性回归分析、及聚类分析,旨在建立短枝木麻黄在寒冷胁迫下的耐寒评价模型。
1 材料与方法
1.1 材料
供试的7个种源的短枝木麻黄种子由澳大利亚林木种子中心(CSIRO)提供,种源信息见表1。2012年4月播种,6月移栽于试验地苗圃。试验地设在广州市中国林业科学研究院热带林业研究所苗圃,位于东经113°15′,北纬23°6′,海拔43 m。属于南亚热带季风气候,热量充足,年平均气温22℃,极端最高温39.3℃,极端最低温0℃。雨量充沛,年均降雨量1 800 mm,主要集中在4~9月,年平均相对湿度74%。
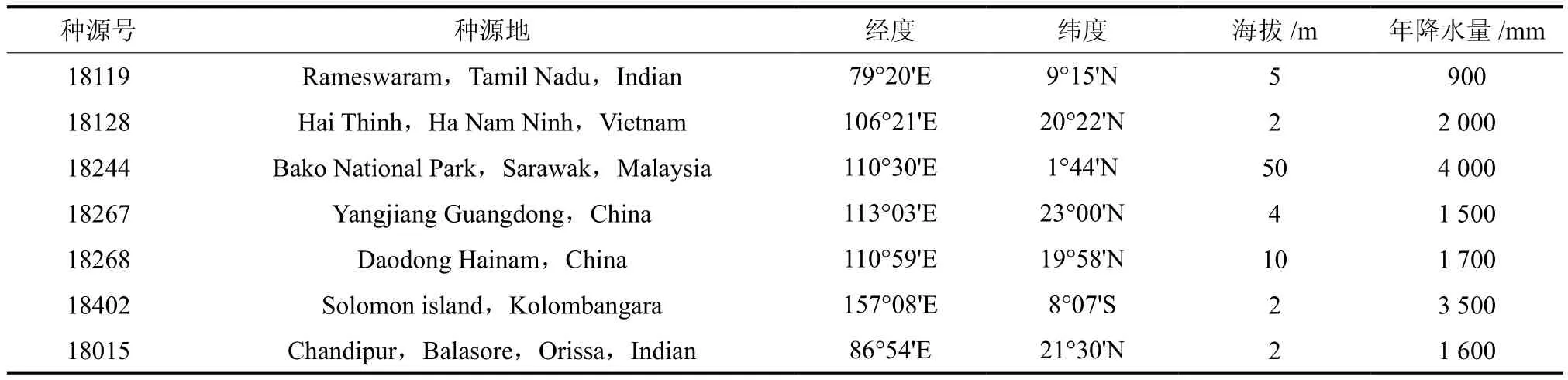
表1 7个参试种源的种源信息Table 1 Information of 7 experimental provenances
1.2 方法
1.2.1 低温处理
人工气候箱中,采用双因素随机区组设计,参试种源7个,苗龄为2a,以每种源随机选取10株苗的30个离体小枝条作为生理指标测试材料,采用25 ℃,5 ℃,0 ℃,-5 ℃,-10 ℃五个温度梯度处理24 h,其中25℃为对照温度组,每个温度处理重复3次,合计处理苗木数为1 050株,即7种源×5温度处理×10株×3重复。
1.2.2 生理指标测定
小枝的电解质渗透率(REC)采用电导率法[6:203-204];丙二醛含量(MDA)用硫代巴比妥酸法[7];蒽酮比色法[8:144-148]测定可溶性糖含量(SS);磺基水杨酸法[8:228-231]测定脯氨酸含量(Pro);考马斯亮蓝G-250法[8:142-143]测定可溶性蛋白(SP);超氧化物歧化酶活性(SOD)采用NBT光化还原法[6:191-193],叶绿素a含量(Chla)、叶绿素b含量(Chlb)、总叶绿素含量(Chl)采用丙酮-乙醇混合液浸提比色法[8:74-77]。
1.2.3 数据统计与分析
数据整理与分析采用Microsoft Excel 2010,用皮尔逊积差进行相关分析,用SPSS 19.0对数据进行主成分分析、回归分析和聚类分析。以处理温度和电解质渗透率拟合Logistic曲线计算半致死温度LT50,具体计算方法参考时朝等[9]的方法。
除电解质渗透率外的8个耐寒指标,为消除不同量纲数据处理带来的影响,通过公式计算成耐寒系数[10]:各指标耐寒系数(α)=处理测定值/对照测定值
2 结果与分析
2.1 寒冷胁迫对抗寒生理指标的影响及耐寒指标筛选
2.1.1 寒冷胁迫对细胞膜系统指标的影响
由表2可知,小枝的电解质相对渗透率均随着温度的降低逐渐上升,在5 ℃,0 ℃渗透率变化幅度小,说明该温度对细胞膜的破坏性不大,继续降温到在-5 ℃,-10 ℃处相对电导率大幅度,膜透性增加,细胞膜遭到破坏。
植物在遭受伤害时与膜脂的过氧化作用密切相关,MDA反映植物在遭受伤害的过程中膜过脂氧化物的程度。总体的MDA含量均是处理组大于对照组,但在5 ℃~0 ℃过程中有两个种源的MDA 含量低于对照组,该温度处的MDA含量可能与耐寒性相关,记为MDA1。在5个处理温度处理下,MDA 的含量总体的趋势呈M型或N型(表2),说明在5 ℃附近植物有一个明显的自我修复的过程,但在-5 ℃~10 ℃处,MDA的变化趋势不一,记为MDA2。
2.1.2 寒冷胁迫对渗透性调节物质的影响
游离脯氨酸、可溶性蛋白和可溶性糖,是植物细胞中重要的渗透物质。在低温锻炼过程中,可溶性蛋白含量、可溶性糖和游离脯氨酸都呈现出两种变化,一种为逐渐上升,可能是在温度降低以后,可溶性蛋白、可溶性糖和游离脯氨酸含量的增加,可以有效提高细胞内的束缚水的能力,降低细胞的冰点,说明这些种源的抗性仍有继续扩大的可能性[11]。另一种为先升后降,可能是过低的温度使得植物细胞的机体运行能力下降,渗透调节物质开始降解[12:16-19]。不同种源的细胞渗透调节能力体现植物的耐寒能力。对照~5 ℃和0 ℃~-5 ℃的可溶性蛋白呈显著正相关(表3),分别记为SP1和SP2。-5 ℃~-10 ℃的可溶性糖变化趋势不一,记为SS1。
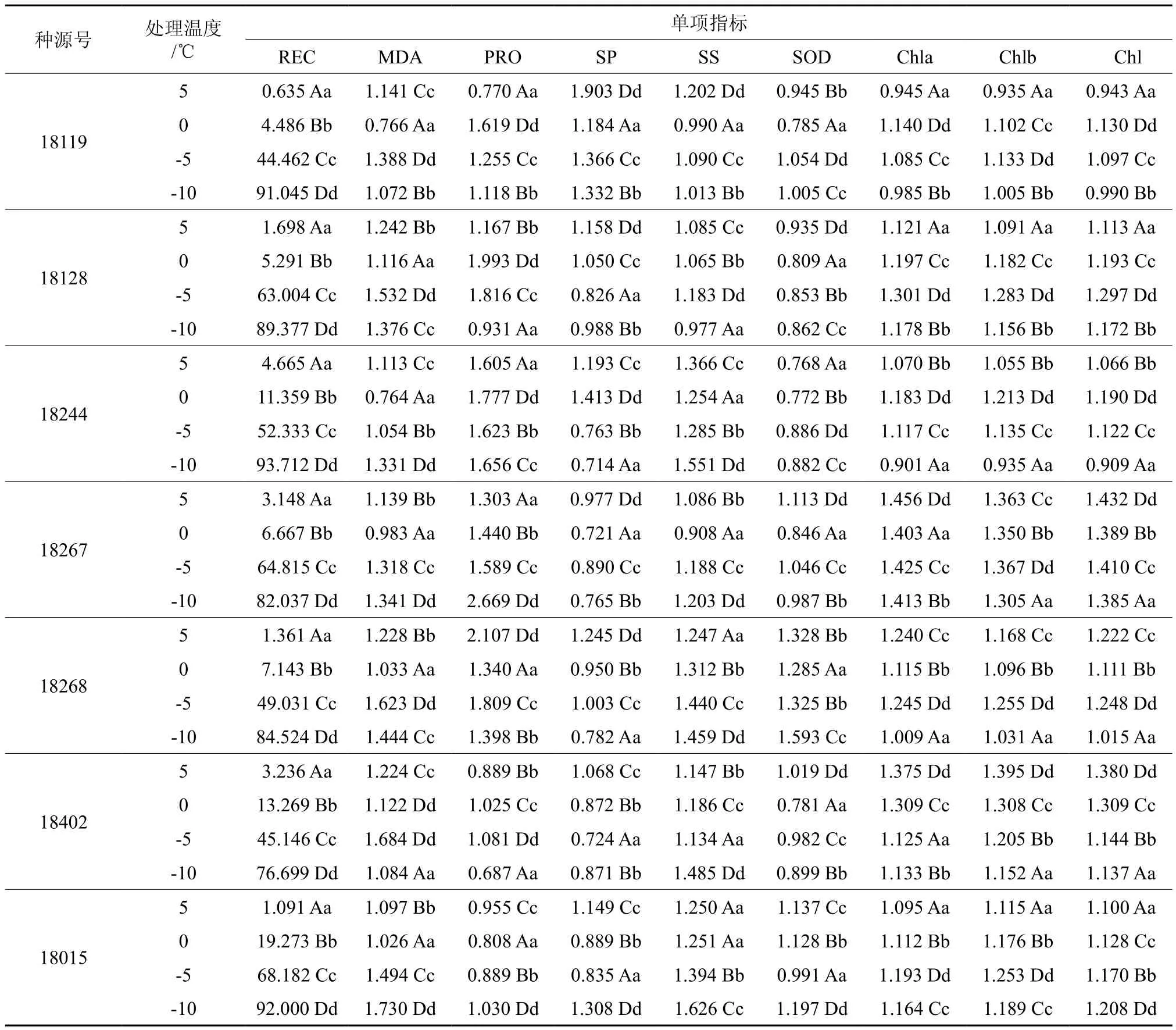
表2 不同温度处理对参试种源生理指标的影响†Table 2 Effects of different concentrations frost stress on physiological indicators of tested provenances
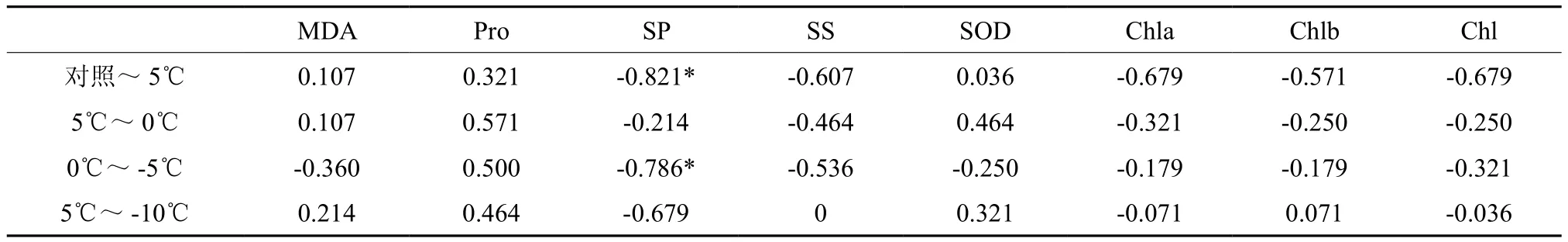
表3 LT50与不同温度下耐寒指标的相关分析†Table 3 Correlation analyze between LT50 and cold tolerance index at different temperature
2.1.3 寒冷胁迫对保护性酶和光合物质的影响
在植物体遭受寒冷胁迫时,植物体内的自由基增加,SOD是植物体内清除自由基的重要酶之一,在本实验中,SOD出现反复的波动,体现了SOD在植物体内保持修复平衡。在对照~5 ℃和-5 ℃到-10 ℃处的变化趋势出现分歧,分别记为SOD1,SOD2。
类囊体膜的组分、透性和流动性、及其叶绿素会响应植物体的低温胁迫[13]。随着温度的降低,7个种源的短枝木麻黄的总叶绿素含量,叶绿素a/b含量都呈现先增加后减少或是平稳的趋势,这与柏科的结果一致[12:19-20],随着温度的降低,细胞开始脱水,而叶绿体还保持其光合的能力,因此叶绿素的相对含量升高。温度继续降低,叶绿体的结构遭到破坏,叶绿素的活性受到抑制甚至叶绿素本身开始降解,使得叶绿素含量降低,这与木麻黄在低温下的枝条变红褐色的形态结果一致。叶绿素a的变化趋势呈现出两个大的方向,可能与耐寒性相关,4个温度区间的叶绿素含量分别记为 Chl1,Chl2,Chl3和Chl4;叶绿素 b的含量在5 ℃~-0 ℃和0 ℃~-5 ℃出现三个明显不同的变化趋势,分别记为Chlb1和Chlb2。在0~-5℃区间的总叶绿素记为Chl1。
2.1.4 半致死温度LT50的确定
LT50能有效的反映植物的耐寒性能,通过相对渗透率拟合Logistic曲线[14]求出各种源半致死温度LT50。半致死温度均在-5℃附近,依据半致死温度对短枝木麻黄耐寒性进行排序依次为18402(-5.956 ℃)> 18268(-5.930 ℃)> 18119(-5.494℃)> 18267(-5.220℃) > 18128(-4.940℃) >18015(-3.669℃)>18244(-3.475℃)。
2.2 短枝木麻黄抗寒性综合评价体系的建立
根据上述上生理指标的分析,筛选出的15个与短枝木麻黄抗寒性密切相关的单项生理指标。即 LT50,MDA1,MDA2,SP1,SP2,SS1,SOD1,SOD2,Chla1,Chla2,Chla3,Chla4,Chlb1,Chlb2,Chl1。
2.2.1 主成分分析
以15个单项指标为变量进行主成分分析,累计贡献率达到96.314%,说明该15个指标已经包含了短枝木麻黄耐寒的大部分信息。主成分1包含的信息主要为叶绿素a,保护酶和可溶性蛋白。主成分2主要携带膜系统和SS1的信息。主成分3携带叶绿素b和SP的信息,主成分4携带的信息为总叶绿素,最后一个主成分携带SOD的信息。从各种源在各主成分的得分情况看,在各种源的得分最高的主成分不尽相同,在4个主成分中都有分布。因此难以通过种源在主成分的得分情况揭示短枝木麻黄耐寒的特性。还需借助回归进一步分析。
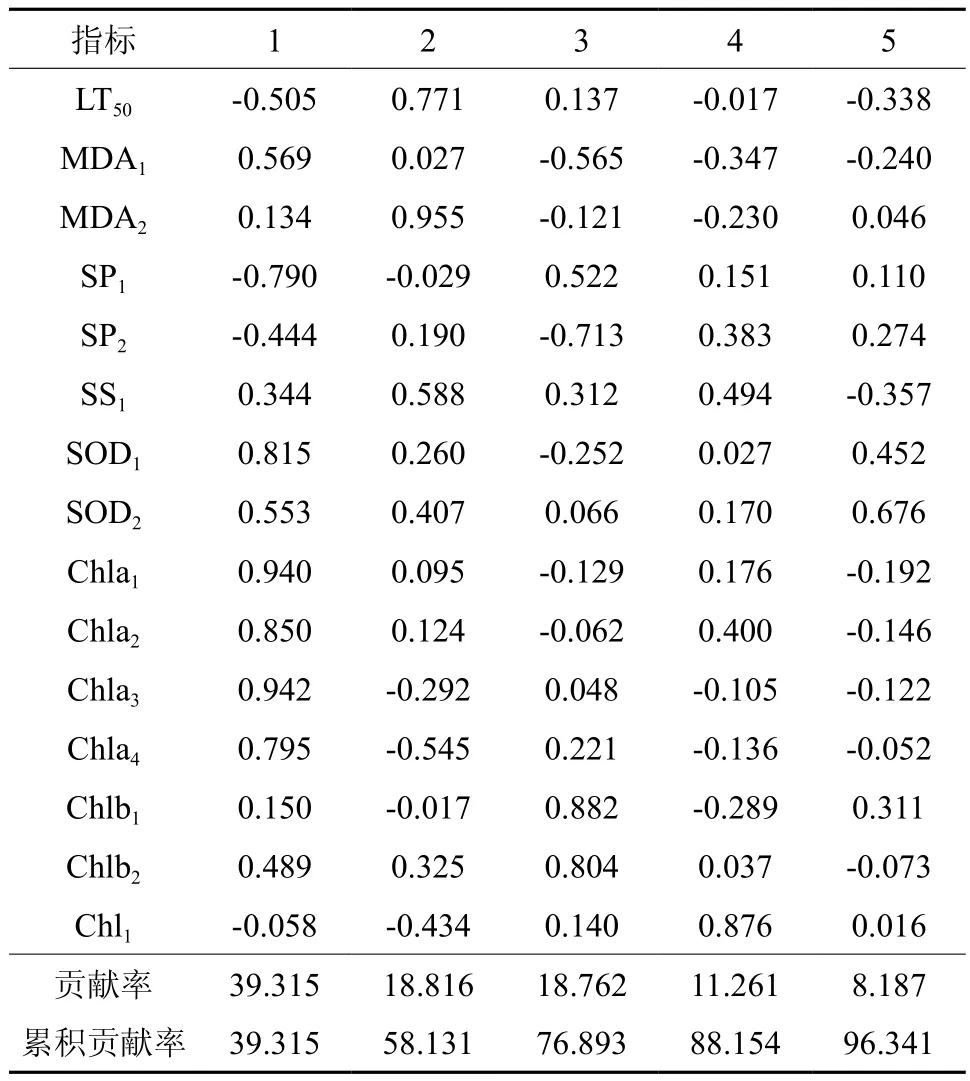
表4 短枝木麻黄耐寒综合评价主成分分析Table 4 Principal component evaluation based on cold tolerance of C. equisetifolia
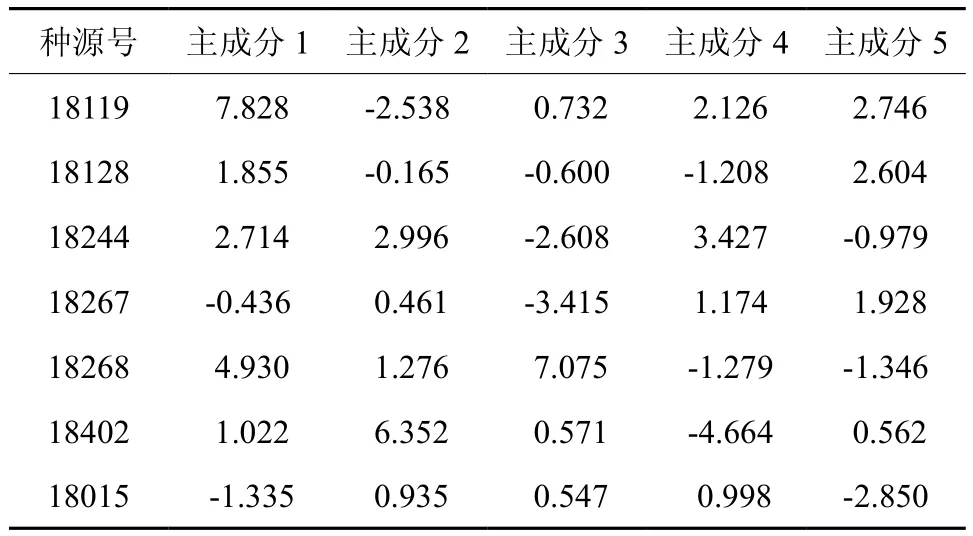
表5 参试种源主成分得分Table 5 Principal component factor scores of tested provenances
2.2.2 回归分析
以LT50为因变量,剩下的14个指标作为自变量,以回归方程中F检验的相伴概率值小于0.05的选入方程,大于0.1的移出方程进行逐步回归分 析。14个 指 标 中 Chla2,Chla3,Chla4,SP1,MDA1被引入方程,回归方程为:
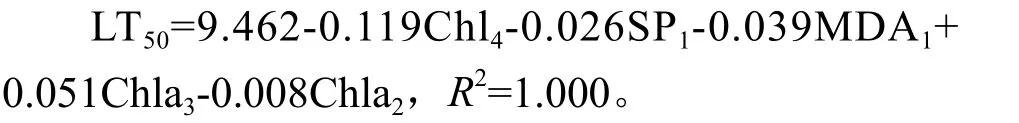
从回归方程可以看出短枝木麻黄的耐寒性主要与叶绿素a在-5 ℃~-10 ℃的变化呈负相关,也与叶绿素a在0 ℃~5 ℃呈一定的正相关。将数据代入方程对方程精度进行评价,由表6 可知方程的估计精度均在98%以上,说明该方程符合对短枝木麻黄耐寒性评价的要求。
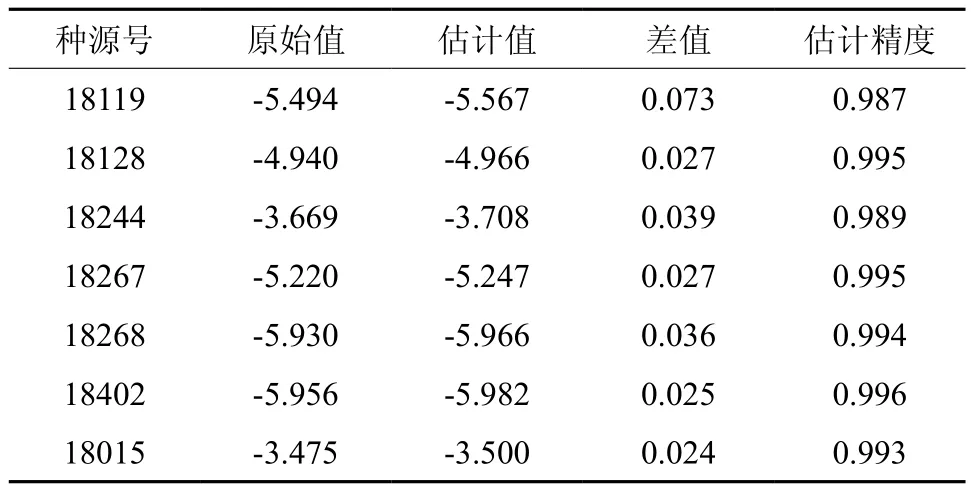
表6 回归方程的估计精度分析Table 6 Estimated accuracy analysis of regression equation
2.2.3 聚类分析
通过回归得出影响短枝木麻黄耐寒性的5个指标,以LT50和5个指标的耐寒系数进行聚类分析(表7),第一类中,半致死温度低,光合物质的平均值在4个类别中都是最大值。第二类中,半致死温度较低,光合物质的平均值都处于中等水平,但可溶性蛋白在对照~5 ℃处处于4类中的最高水平。第三类的半致死温度明显低于第二类,Chla4在4类中最低,且MDA的含量在5 ℃~0 ℃温度变化过程中偏高SP1也比第二类低很多。第四类半致死温度最高,最不耐寒,光合物质相比前三大类别都偏低。
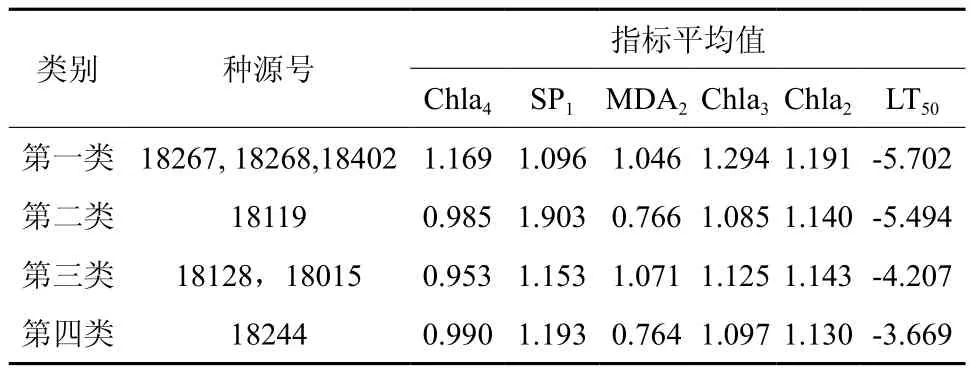
表7 聚类分析结果Table 7 Results of cluster analysis
3 结论与讨论
寒冷胁迫是植物重要的环境胁迫之一,植物的抗寒性与自然对植物的选择和植物对自然的适应过程中,微效并特异的抗寒基因进调控有关。低温半致死温度是植物在遭受寒冷胁迫后能否恢复生长的一个临界值,能较准确反映植物的抗寒能力[15-17]。本试验通过短枝木麻黄小枝在低温胁迫下的渗透率拟合Logistic曲线得出半致死温度,根据半致死温度对7个种源的耐寒顺序排序,各种源号的耐寒力从大到小依次为18402>18268>18119>18267>18128>18015>18244。
主成分分析可以简化数据集,保持对方差贡献大的数据。本文通过单项指标分析结合相关分析对耐寒指标进行简化选出与耐寒性密切相关的15个指标。通过主成分分析,15个指标的贡献率达到96.341%,说明该15个指标包含短枝木麻黄耐寒的大部分信息。通过线性回归对指标进一步筛选,并得出回归方程LT50=9.462-0.119Chl4-0.026SP1-0.039MDA1+0.051Chla3-0.008Chla2,通过半致死温度的精度分析和聚类分析进行验证。
在植物中,耐寒能力强的植物保持光合作用的能力要比耐寒性差的强[18-19]。植物在低温逆境下叶绿素降低,光和机能的破坏导致植物死亡[20-21]。在本文中叶绿素a的含量在回归方程中占三个指标,体现了其对短枝木麻黄耐寒性的影响的重要性。经过聚类分析也表明,叶绿素a的含量是影响聚类结果的重要的指标。
[1] 仲崇禄,白嘉雨,张 勇.我国木麻黄种质资源引种与保存[J].林业科学研究,2005,18(3):345-350.
[2] Midglev S J, Turnbull J W, Johnston R D,et al. Casuarina ecology management and utilization [C]. International Workshop Casuarina Ecology Management and Utilization. Canberra(Australia), 1981, 17-21.
[3] 邬 金,温国胜,王电杰,等.低温胁迫下7 种木麻黄变异类型抗寒性的比较[J].福建林学院报,2013,33(1):34-37.
[4] 武 冲,张 勇,马 妮,等.接种菌根菌短枝木麻黄对低温胁迫的响应特征[J].西北植物学报,2012,32(10):2068-2074.
[5] 何贵平,卓仁英,陈雨春,等.低温处理对耐寒粗枝木麻黄无性系生理指标的影响[J].林业科学研究,2011,24(4):523-526.
[6] 张蜀秋.植物生理学实验技术教程[M].北京:科学出版社,2011.
[7] 赵世杰,许长城,邹 琦,等.植物组织中丙二醛测定方法的改进[J].植物生理学通讯,1994,30(3):207-210.
[8] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[9] 时 朝,王亚芝,刘国杰.应用Logistic方程确定五种苹果枝条的半致死温度的研究[J].北方园艺,2013,(2):36-38.
[10] 周广生,梅方竹,周竹青,等.小麦不同品种耐湿性生理指标评价及预测[J].中国农业科学,2003,36(11):1378-1382.
[11] 高志红,章镇,韩振海.果树种质枝条抗寒性鉴定[J].果树学报,2005,22(6):709-712.
[12] 姚 红.四个柏科植物品种抗寒性生理生化研究[D].呼和浩特:内蒙古农业大学,2012.
[13] 徐 俊.银丝竹等四种城市观赏竹抗寒性生理研究[D].雅安:四川农业大学,2011.49-50.
[14] 许 瑛,陈发棣.菊花8个品种的低温半致死温度及抗寒适应性[J].园艺学报,2008,35(4):559-564.
[15] 朱根海,刘祖祺,朱培仁. 应用Logistic方程确定植物组织低温半致死温度的研究[J].南京农业大学学报,1986,(3):11-16.
[16] 王荣富.植物抗寒指标的种类及其应用[J].植物生理学通讯,1987, (03):49-55.
[17] 刘 建,项东云,陈健波,等.应用Logistic方程确定三种桉树的低温半致死温度[J].广西林业科学,2009,38(2):75-78.
[18] Kimball S L, Sallsbury F B. Ultrastructural changes of plants exposed to low temperature [J]. Aln J Bot,1973,60(10):1028-1037.
[19] Runich-Bayer S, Krause H G. Freezing damage and frost tolerance of the photosynthetic apparatus studies with isolated mesophyll protoplasts of Valerianella locusta [J]. Photosyn Res,1986, 8: 161-174.
[20] 王 毅,方秀娟,徐 欣,等.黄瓜幼苗低温锻炼对叶片细胞叶绿体结构的影响[J].园艺学报,1995,22(3):299-300.
[21] 林秀琴,黄金水,蔡守平,等. 木麻黄内含物含量与对星天牛抗性关系的研究[J].中南林业科技大学学报,2014,34(5):37-41.
Evaluation and Comprehensive Analysis of Cold Tolerance on SevenCasuarina equisetifoliaProvenances
LIU Fen1,2, JIANG Qin-bin2, MA Ying-zi1, ZHONG Chong-lu2, ZHANG Yong2, CHEN Yu2, CHEN Zhen2
(1.Central South University of Forestry and Technology, Changsha 410004, Hunan, China; 2. Research Institute of Tropical Forestry,CAF, Guangzhou 510520, Guangdong, China)
The 9 physiological traits were measured with seedlings of sevenCasuarina equisetifoliaprovenances, which treated under fi ve low temperature gradients (25℃ , 5℃ , 0℃ , -5℃ and -10℃ ) for 24 hours, the test was repeated 3 times. Ten seedlings’ branches on per treatment per provenance were collected, and theirs nine cold resistance indexes such as permeability, MDA, proline content,soluble protein content, soluble sugar content, etc. were determined in artif i cial climate chamber. The cold resistance capability were evaluated by semilethal temperature(LT50). Combined correlation analysis and characteristics analysis, the cold-resistant indexes were screened. Principal component, regression and cluster analysis were used for further comprehensive analysis on the selected coldresistant indexes. The results show that sevenC. equisetifoliaprovenances were sequenced in freeze resistance capability according to their semi-lethal temperature from big to small as follows: 18402> 18268> 18119> 18267> 18128 > 18015 > 18244; The regression equation was built by stepwise regression method: LT50=9.462-0.119Chl4-0.026SP1-0.039MDA1+0.051Chla3-0.008Chla2,(R2=1.000), the estimation accuracy of the model was higher than 98%.
Casuarina equisetifolia; provenance cold tolerance evaluation; low temperature stress; principal component analysis; linear regression
S718.43;S792.93
A
1673-923X(2015)06-0040-05
10.14067/j.cnki.1673-923x.2015.06.008
2014-11-05
十二五科技支撑计划任务(2012BAD01B0603);广东省林业科技创新招标项目(201401);中央级公益性科研院所基本科研业务费专项基金(RITFYWZX201203)
刘 芬,硕士研究生 通讯作者:马英姿,博士,教授
刘 芬,姜清彬,马英姿,等. 7个短枝木麻黄种源耐寒评价及综合分析[J].中南林业科技大学学报,2015,35(6):40-44,62.
[本文编校:吴 彬]
