家兔小腿屈伸趾肌群肌纤维型增龄变化研究
2015-12-16罗怀香
罗怀香 薛 黔
1(遵义医学院 基础医学教育教研室,遵义563099)
2(遵医学院 人体解剖教研室,遵义563099)
人类衰老过程中,肌肉是人体最先衰老的器官之一。自然增龄过程中,人体会出现的退行性改变主要表现为肌肉萎缩和力量水平的下降,导致人体肌肉功能结构减少并且增加慢性代谢性疾病发展的危险。骨骼肌质量下降的现象始于50岁左右,但是更明显的减少是在60岁以后[1]。对于衰老过程中骨骼肌的改变,主要集中表现在肌纤维类型、肌纤维数目和肌纤维横切面积的变化等方面[2]。本实验采用NADH-TR\染色方法对生后4周龄、12月龄、24月龄和36月龄家兔屈伸趾肌群进行研究,观察自然增龄过程中肌纤维型的变化,为人类老龄化骨骼肌的研究提供比较解剖学的资料。
1 材料与方法
1.1 实验动物与分组
各年龄段家兔共24只,分别是出生后4周、12月、24月、36月龄,雌雄不拘,均购自遵义医学院实验动物中心。本实验选用的动物模型是自然增龄的家兔,按动物实验计算好的生理能量,在一定的饲养条件下,让其自然生长,一般不通过药物干预的装置,从而使家兔逐渐达到该时间段的实验要求。
1.2 实验方法
1.2.1 实验材料 家兔空气栓塞处死,取双侧趾长伸肌、 长伸肌、趾浅屈肌和趾深屈肌,剔除肌肉表面筋膜,切取肌肉腹中部(0.5cm×0.2cm)组织块,锡纸包裹,置入液氮(-196℃)冷冻。
1.2.2 组织切片准备 液氮内取出组织块,置于横冷切片箱,在-22℃条件下作冰冻切片,切片厚度为10μm,室温下干燥30~60min。
1.2.3 组织切片的 NADH-TR酶染色 选用Tris-HCL缓冲液配置孵育液并染色,切片置37℃条件下孵育30~40min,脱水,透明,中性树胶封片。1.2.4 NADH-TR染色后,取趾长伸肌、 长伸肌、趾浅屈肌和趾深屈肌各年龄段样本切片,图像分析系统下每例测数10张切片,每张切片随机测数10个视野,每个视野面积为500×500μm2。
1.3 统计学方法
应用SPSS 17.0软件。实验数据用均数加减标准差(±s)来表示,用单因素方差进行分析,检验水准α=0.05。
2 结果
2.1 染色所见
经NADH-TR染色后,显微镜下观察,在4周龄组至36月龄组家兔屈伸趾肌群的切片上,能明显区分出三型肌纤维,即Ⅰ型、ⅡA型和ⅡB型。其中,Ⅰ型肌纤维横切面积最小,染色最深,呈深蓝紫色,胞浆内蓝紫色颗粒粗大,分布较均匀;ⅡB型肌纤维横切面积最大,染色最浅,呈淡蓝色,胞浆内的染色颗粒分布均匀;ⅡA型肌纤维染色中等,呈淡蓝紫色,胞浆内染色颗粒粗大,肌膜下分布多于中央(见图1),其中在4周龄组、12月龄组24月龄组肌染色切片上,镜下可见呈深蓝紫色、淡蓝紫色和淡蓝色的三型肌纤维,即Ⅰ型、ⅡA型和ⅡB型。三型肌纤维呈棋盘状镶嵌分布,肌纤维形态多呈椭圆形和圆形(见图1A、B、C);在36月龄组肌纤维的横切面多呈三角形,可见小角化肌纤维及10余条同一型肌纤维聚集在一起,以Ⅰ型肌纤维为主(见图1D)。
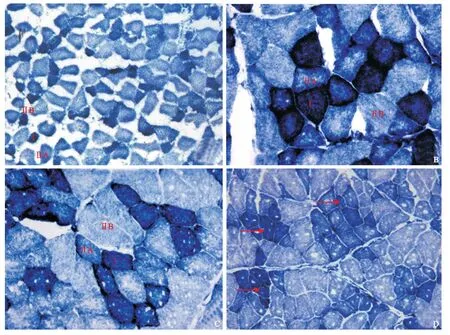
图1 各年龄组家兔趾浅屈肌型增龄变化(NADH-TR染色,×200)
2.2 肌纤维型的构成比变化
屈伸趾肌群各实验组三型肌纤维的构成比(见表1,2,3,4)。伸趾肌群中,趾长伸肌和 长伸肌Ⅰ型肌纤维构成比随增龄均呈梯度下降,到24月龄降到最低,36月龄时比例上升,在24月龄组的基础上,36月龄组Ⅰ型肌纤维分别增加了0.66%、16.07%,长伸肌增加有统计学差异,P<0.05;ⅡB型肌纤维构成比随增龄均逐渐增加,到24月龄时增加到最大值,36月龄时比例下降,在24月龄组的基础上,36月龄组分别下降了16.94%、33.50%,而ⅡA型则分别增加了25.86%、25.98%,伸趾肌群ⅡA型和ⅡB型变化均有统计学差异,P<0.05。屈趾肌群中,趾浅屈趾和趾深屈肌4周龄组至24月龄组Ⅰ型肌纤维构成比随增龄呈梯度下降,到24月龄降到最低,在36月龄时比例上升。ⅡB型肌纤维构成比的变化恰好相反。在24月龄组的基础上,36月龄组Ⅰ型肌纤维分别增加了1.17%、18.39%,ⅡB型分别下降了0.47%、17.27%,趾浅屈肌ⅡA型下降了0.43%,趾深屈肌增加了15.70%,其中趾深屈肌三型肌纤维变化均有统计学差异,P<0.05。
表1 生后发育和增龄过程中家兔趾趾长伸肌纤维构成比(n=6,±s,%)

表1 生后发育和增龄过程中家兔趾趾长伸肌纤维构成比(n=6,±s,%)
注:★与24月龄组比较P<0.05
年龄 趾长伸肌Ⅰ型(%) ⅡA型(%) ⅡB型(%)4周 47.83±3.74★ 23.74±3.45 28.43±1.63★12月 31.52±0.35 32.58±1.45★ 35.90±1.71★24月 29.97±1.37 22.50±1.51 47.53±1.72 36月 30.17±1.66 30.35±2.38★ 39.48±2.53★
表2 生后发育和增龄过程中家兔 长伸肌纤维构成比(n=6,±s,%)

表2 生后发育和增龄过程中家兔 长伸肌纤维构成比(n=6,±s,%)
注:★与24月龄组比较P<0.05
年龄 长伸肌Ⅰ型(%) ⅡA型(%) ⅡB型(%)4周 47.70±3.26★ 25.28±3.19★ 27.02±2.04★12月 27.17±0.31★ 29.46±0.59 43.37±0.65 24月 21.68±1.22 32.20±2.84 46.12±4.13 36月 25.83±2.39★ 43.50±2.05★ 30.67±1.55★
表3 生后发育和增龄过程中家兔趾浅屈肌纤维构成比(n=6,±s,%)

表3 生后发育和增龄过程中家兔趾浅屈肌纤维构成比(n=6,±s,%)
注:★与24月龄组比较P<0.05
年龄 趾浅屈肌Ⅰ型(%) ⅡA型(%) ⅡB型(%)4周 46.25±0.38★ 27.42±1.24★ 26.33±1.09★12月 26.18±2.19 26.52±1.90★ 47.30±1.33★24月 27.97±1.49 21.11±1.37 50.92±1.50 36月28.30±0.83 21.02±1.17 50.68±1.82
3 讨论
生后发育早期(前几周)是哺乳动物骨骼肌肌纤维生长和分化的主要阶段[3]。在生后发育过程中,同一动物不同肌肉各型肌纤维分化发育规律不同[4]。本研究通过NADH-TR染色发现:生后4周龄时,可清晰观察到Ⅰ型及Ⅱ型纤维的亚型纤维,即ⅡA和ⅡB型。三型肌纤维中,Ⅰ型比例最大。家兔屈伸趾肌群肌纤维自4周至24月龄组Ⅰ型纤维均逐渐减少,至24月龄组时下降到最低。ⅡA型在各个年龄段变化幅度小。ⅡB型纤维构成比的变化与Ⅰ型纤维恰好相反,即4周至24月比例逐渐增加,至24月龄时达到最高,与朱道立等人研究结果一致[5]。
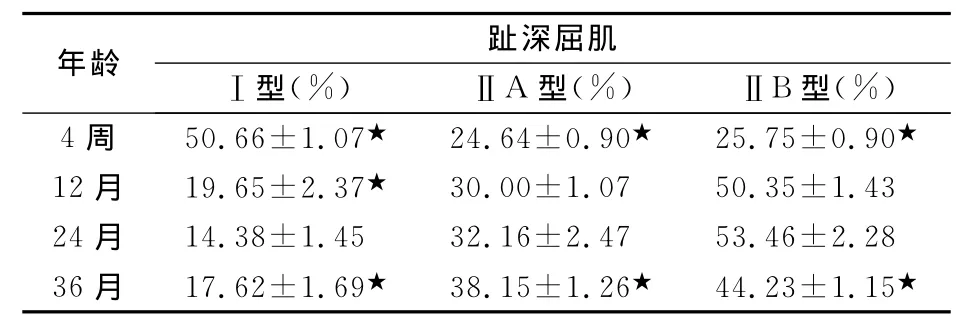
表4 生后发育和增龄过程中家兔趾深屈肌纤维构成比(n=6x±s,%)
Aniansson等[6]对老年男性的股外侧肌和肱二头肌的纵向研究表明:与青年组相比,股外侧肌和肱二头肌的ⅡB型肌纤维的比例都下降了50%。Lexell等[7],与年轻人相比,在老年人的股外侧肌中,肌纤维数量下降40%,Ⅱ型肌纤维减少25%~30%。Caccia通过研究比较3月龄和36月龄的大鼠趾长伸肌和比目鱼肌得出,比目鱼肌从3~4月龄到30月龄,Ⅰ型肌纤维比例从73.5%减少到40.5%,ⅡA型肌纤维比例增加,而趾长伸肌Ⅰ型肌纤维的百分比是增加的,ⅡA和ⅡB型是减少的[8]。本研究得出,36月龄老年组家兔与12月龄青年组相比,趾长伸肌和趾浅屈肌ⅡA型肌纤维分别下降了6.84%、20.59%, 长伸肌和趾深屈肌ⅡB型肌纤维分别下降29.28%、12.15%。与24月龄组相比,36月龄组屈伸趾肌群Ⅰ型肌纤维比例均有增加,分别为趾长伸肌0.66%、 长伸肌16.07%、趾浅屈肌1.17%、趾深屈肌18.39%;ⅡB型肌纤维呈下降趋势,下降比例分别为:趾长伸肌16.94%, 长伸肌33.50%,趾浅屈肌0.47%,趾深屈肌17.27%。与以上研究结果一致,随着增龄改变,Ⅰ型肌纤维比例增加,Ⅱ型肌纤维比例下降。
据肌纤维型研究结果显示,在青年人的肌肉,不同类型肌纤维呈马赛克状分布,在老年人,表现为相同类型的肌纤维型聚集在一起,即肌纤维类聚(“Fiber-type grouping”)[7]。Kung等[9]使用 ATP酶染色分别对成年组大鼠(11~13月龄)和老年组大鼠(36~37月龄)的趾长伸肌进行肌纤维型的观察,染色表明,成年组大鼠三型肌纤维镶嵌分布呈棋盘状,老年组则观察到3~4个Ⅰ型肌纤维呈肌纤维类聚。Andersen等[10]分别对22岁和87岁两个年龄组肌肉使用ATP酶染色得出,22岁年龄组肌纤维呈现棋盘状分布,而87岁年龄组肌纤维明显出现了肌纤维类聚现象。这种随着年龄增加肌纤维型组成改变的现象是失神经和神经再支配的过程[11]。本实验4周至24月龄组军观察到三型肌纤维呈棋盘状分布,36月龄组老年家兔屈伸趾肌群均可见10余条肌纤维聚集在一起,且Ⅰ型纤维较明显,我们推测Ⅱ型肌纤维运动单位开始向Ⅰ型肌纤维转换。骨骼肌肌纤维萎缩的组织学表现为:肌纤维小角化、圆形化、肌纤维直径的差别增大等[12]。Andersen使用ATP酶染色观察到的老年人肌纤维形态多样,多呈扁平或者“煎薄饼”、“香蕉”状肌纤维[10],黄丽敏等[13]使用Gomori染色研究得出,老年组(17月龄)和高龄老年组(30月龄)大鼠腓肠肌肌纤维开始萎缩甚至坏死,多呈现小角化纤维。本实验中36月龄组各型肌纤维的大小和形态出现多样性改变,多呈三角形或不规则形,还可观察到小角化肌纤维,可能是肌细胞凋亡的早期迹象,但仍需用高龄动物肌研究进行进一步观察和验证。
[1] Valeria Z,Renato G,Luisa C,et al.Interventions Against Sarcopenia In Older Persons [J].Current Pharm Aceutical Design,2014,3(16).
[2] Koopman R.Dietary protein and exercise training in ageing[J].Proceedings of the Nutrition Society,2011,70(01):104-113.
[3] White RB,Biérinx AS,Gnocchi VF,et al.Dynamics of muscle fibre growth during postnatal mouse development[J].BMC Developmental Biology,2010,10(1):21.
[4] Punkt K,Naupert A,Asmussen G.Differentiation of rat skeletal muscle fibres during development and ageing[J].Acta Histochemica,2004,106(2):145-154.
[5] 朱道立,陈佩林,叶辉,等.家兔生后快慢肌纤维 MyHCs的转化比较[J].江苏农业科学.2009,1(2)208-212.
[6] Aniansson A,Grimby G,Hedberg M.Compensatory muscle fiber hypertrophy in elderly men [J].Journal of Applied Physiology,1992,73(3):812-816.
[7] Lexell J,Taylor C C,Sjöström M.What is the cause of the ageing atrophy?:Total number,size and proportion of different fiber types studied in whole vastus lateralis muscle from 15-to 83-year-old men [J].Journal of the Neurological Sciences,1988,84(2):275-294.
[8] Caccia M R,Harris J B,Johnson M A.Morphology and physiology of skeletal muscle in aging rodents[J].Muscle &Nerve,1979,2(3):202-212
[9] Kung T A,Cederna P S,van der Meulen J H,et al.Motor Unit Changes Seen With Skeletal Muscle Sarcopenia in Oldest Old Rats[J].The Journals of Gerontology Series A:Biological Sciences and Medical Sciences,2013,4:1-9.
[10] Andersen J L.Muscle fibre type adaptation in the elderly human muscle[J].Scandinavian Journal of Medicine & Science in sports,2003,13(1):40-47.
[11] Aniansson A,Grimby G,Hedberg M.Compensatory muscle fiber hypertrophy in elderly men [J].Journal of Applied Physiology,1992,73(3):812-816.
[12] 壄中征哉(日)著,吴士文 马维娅 译.临床肌肉病理学(第三版修行版)[M].北京:人民军医出版社,2007:10.
[13] 黄丽敏.负重跑台训练对衰老大鼠骨骼肌的作用效果及MGF基因表达的研究[D].河北师范大学,2010.