孑遗植物四合木(Tetraena mongolica)迁地保护中的光合作用日变化特征与生理生态适应性
2015-12-05智颖飙李红丽崔艳路战远刘珮叶学华张荷亮杨持刘钟龄王云飞华宇鹏红鸽赵凯魏玲玲王强
智颖飙,李红丽,崔艳,路战远,刘珮,叶学华,张荷亮*,杨持,刘钟龄,王云飞,华宇鹏,红鸽,赵凯,魏玲玲,王强
1. 内蒙古大学环境与资源学院,内蒙古 呼和浩特 010021;2. 内蒙古大学生命科学学院,内蒙古 呼和浩特 010021;3. 内蒙古大学鄂尔多斯学院,内蒙古 鄂尔多斯 017000;4. 北京林业大学自然保护区学院,北京 100083;5. 内蒙古农牧业科学院,内蒙古 呼和浩特 010021;6. 植被与环境变化国家重点实验室,中国科学院植物研究所,北京 100093;7. 南京师范大学地理科学学院,江苏 南京 210039;8. 西尔多斯国家级自然保护区管理局,内蒙古 乌海 016000
孑遗植物四合木(Tetraena mongolica)迁地保护中的光合作用日变化特征与生理生态适应性
智颖飙1,2,3,李红丽4,崔艳5,路战远5,刘珮1,叶学华6,张荷亮3*,杨持2,刘钟龄2,王云飞3,华宇鹏3,红鸽3,赵凯7,魏玲玲6,王强8
1. 内蒙古大学环境与资源学院,内蒙古 呼和浩特 010021;2. 内蒙古大学生命科学学院,内蒙古 呼和浩特 010021;3. 内蒙古大学鄂尔多斯学院,内蒙古 鄂尔多斯 017000;4. 北京林业大学自然保护区学院,北京 100083;5. 内蒙古农牧业科学院,内蒙古 呼和浩特 010021;6. 植被与环境变化国家重点实验室,中国科学院植物研究所,北京 100093;7. 南京师范大学地理科学学院,江苏 南京 210039;8. 西尔多斯国家级自然保护区管理局,内蒙古 乌海 016000
为了进一步廓清迁地保护条件下孑遗植物四合木(Tetraena mongolica)的光合生理生态适应性,在分析了瞬时光合效率的基础上,应用LI-6400光合作用测定系统测定了迁地保护试验区的四合木以及原生境伴生种白刺(Nitraria tangutorum)的光合作用日变化,并测定了其生长量。结果表明:四合木实生苗的生长南北冠幅大小依次为乌海四合木核心区实生苗(26.48 cm×27.26 cm)>鄂尔多斯实生苗(21.27 cm×21.75 cm)>阿拉善实生苗(19.25 cm×18.27 cm)。在原生境地乌海四合木核心区种植的实生苗与阿拉善实生苗之间的生长量存在显著差异(P≤0.01),与鄂尔多斯实生苗之间的生长量存在显著差异(P≤0.05)。迁地保护条件下四合木生实生苗植株叶片光合速率Pn日变化均呈“双峰”曲线。不同试验区四合木光合作用日变化(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间 CO2浓度(Ci)均表现出明显的分异。迁地保护四合木条件下原生境地栽培的四合木实生苗的光合速率>鄂尔多斯栽培的四合木实生苗的光合速率>阿拉善栽培的四合木实生苗。鉴此可以作进一步推论,孑遗濒危植物四合木从原生境地西鄂尔多斯核心区(乌海)东移进行迁地保护具有更高的生理生态适应性和生境适宜性。但完成“从种子到种子”进而实现“保存性和代表性”,最终实现四合木迁地保护的“保持性和防止性”,保持其遗传多样性和遗传稳定性,最终成功实现四合木的迁地保护仍有待作进一步探索和深入研究。
迁地保护;生境适宜性;光合作用;保持力;多基因库;四合木
四合木(Tetraena mongolica)是蒺藜科(Zygophyllaceae)四合木属的单种属的古地中海孑遗植物(徐朗然和黄成就,1998;马毓泉,1989)。被誉为“活化石”和植物中的“大熊猫”(黄会青和钱荣,2006)。全世界仅在亚洲中部荒漠区的鄂尔多斯高原西北部及阿拉善东缘有极为有限的分布(分布幅为 106°35′~107°25′E,39°20′~40°15′N)(刘钟龄等,1985476-477)。目前优势种面积仅为461.67 km2(张韬等,2005)。其形态与主产南美洲的金虎尾科Malpighiaceae近缘,反映出与古地中海植物区系成分的密切联系以及在系统演化占据的重要地位(张韬等,2005),在维护草原生态系统平衡上也具有巨大价值(黄会青和钱荣,2006;刘钟龄等,1985684-686;张韬等,2005)。
为了拓展濒危植物的生存空间,植物迁地保护作为植物多样性就地保护的重要辅助措施已得到广泛的应用并取得了系列成果(Brütting等,2013;Moir等,2012;Daniel等,2012;Huang Hongwen,2011;Jason等,2010;傅立国和金鉴明,1991;国家林业局野生动植物与自然保护区管理司,2013;谷安琳和王宗礼,2009;许再富等,2008;许再富,199826-35)。但四合木的迁地保护却异常艰难(智颖飙,2004;智颖飙等,2008)。而光作为影响植物迁地保护十分复杂的环境因子,其太阳辐射的强度、质量以及周期性变化对濒危植物的生长发育、地理分布都产生着深刻的影响(周云龙,2011)。尽管对四合木就地保护和异地保护的生态学过程有所研究(智颖飙,2004;王峰等,2003),但四合木迁地保护生物学中光合生理生态的研究鲜见报道(智颖飙等,2014)。鉴于此,从 2010年伊始,我们从生境适宜性、生态适应性特征和“气候相似论”的角度(许再富等,2008;许再富,199872-81),探索从四合木原生境地乌海向在同一纬度区西移(阿拉善)、东移(鄂尔多斯)进行迁地保护的可能性(智颖飙等,2014;智颖飙,2004)。在分析了气候生物学特征和瞬时光合效率的基础上,2013年测定了迁地保护条件下的四合木的光合作用日变化,目的是为了进一步廓清迁地保护条件下孑遗植物四合木的光合生理生态适应性,以期为四合木的迁地保护提供实践和理论依据,并我国西北干旱草原区其它濒危植物的迁地保护提供案例借鉴。
1 试验区特征与研究方法
1.1 试验区自然特征
研究区包括四合木自然分布区(106°35′~107°25′E,39°20′~40°15′N)(黄会青,2006;智颖飙等,2014)的海勃湾区东郊千里山(39°41′N,106°49′E)、迁地保护试验区的鄂尔多斯市的康巴什新区和阿拉善盟巴彦浩特市东郊的沙生植物园(图1)。四合木分别异地西移184 km,种植在阿拉善盟巴彦浩特市东郊的沙生植物园(38°50′N,105°40′E)。东移397.7 km,种植在的内蒙古大学多斯学院校园绿化区(39°50′N,109°59′E)。
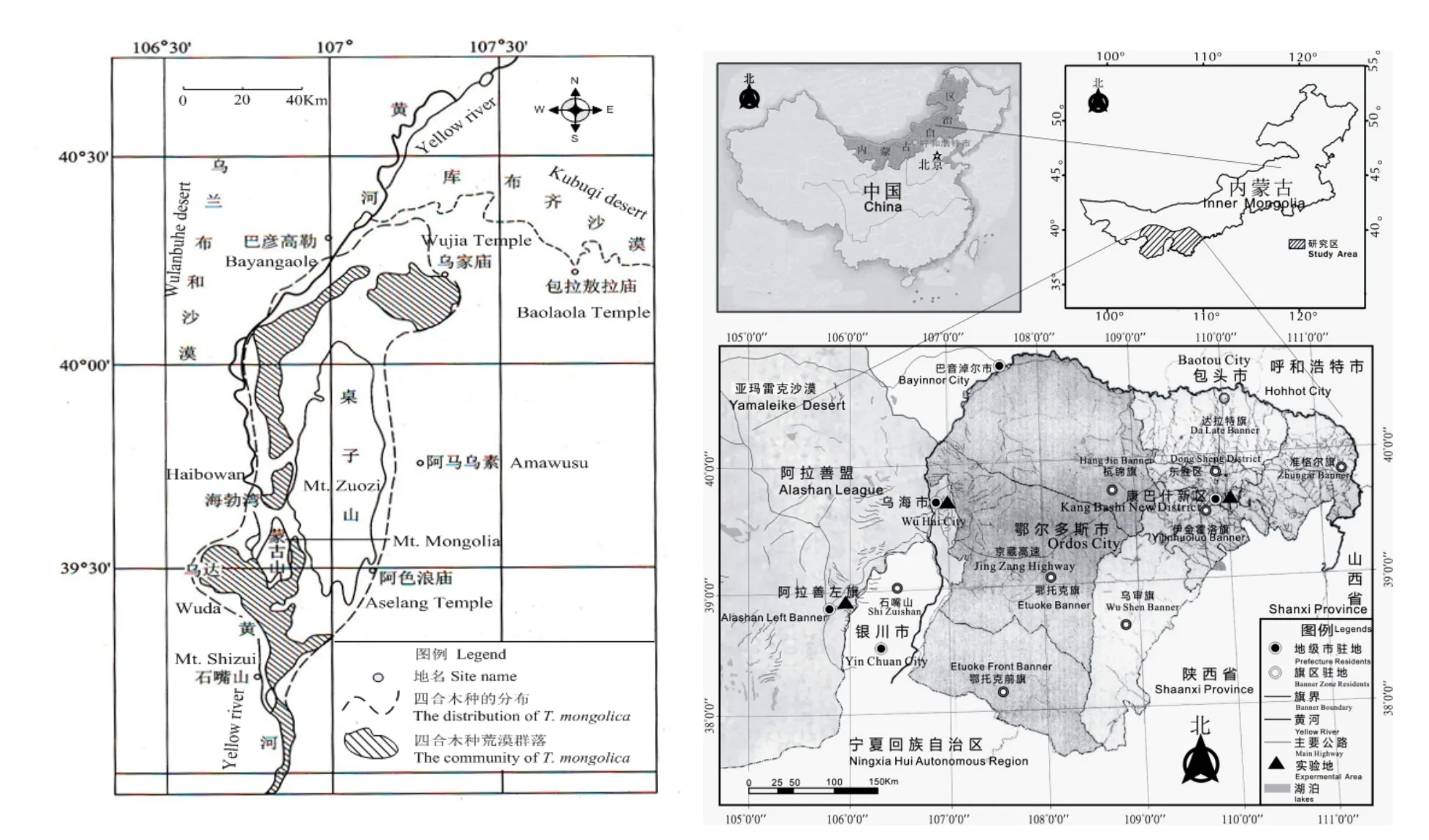
图1 四合木自然分布区与迁地保护试验区Fig. 1 The distributive area and its ex-situ conservation area of T. mongolica
四合木自然核心分布区海勃湾区(地理坐标39°41′N,106°49′E,海拔1091.6 m;土壤类型为棕钙土;年平均气温9.3 ℃,无霜期161 d,有效积温4144 ℃。降雨量162.4 mm,蒸发量3481.1 mm。日照时数3120.7 h,光合有效辐射3138.5 MJ/m2,总辐射量6471.1 MJ/m2)和位于其西边184 km的阿拉善盟巴彦浩特市沙生植物园(地理坐标38°50′N,105°40′E,海拔1561 m。土壤类型为灰褐土。年平均气温7.7 ℃,无霜期147 d,有效积温3064 ℃。降雨量213.1 mm,蒸发量2349.2 mm。日照时数3098.6 h,光合有效辐射2930.8 MJ/m2,总辐射量6042.8 MJ/m2),以及位于其的东边397.7 km的内蒙古大学鄂尔多斯学院校园绿化区(地理坐标39°50′N,109°59′E,海拔1460 m。土壤类型为栗钙土。年平均气温6.3 ℃,无霜期126 d,有效积温 2795 ℃。降雨量 357.42535 mm,蒸发量2349.2 mm。日照时数3019 h,光合有效辐射2871.6 MJ/m2,总辐射量5860.3 MJ/m2)。四合木原生境与迁地保护实验区的阿拉善左旗、鄂尔多斯康巴什新区之间在海拔高度、环境因子(水、肥、气、热和光)、土壤和植被类型均有明显差异(智颖飙等,2014)。
1.2 实验设计
1.2.1 客土栽培
2001年 11月,将鄂尔多斯市东胜区塔垃壕(110°05′46″E,39°47′20″N)的土壤拉运至四合木核心分布区乌海市海渤湾区千里山北麓(106°50′20″E,39°49′34″N)的试验区(将0~20和20~50 cm土壤分别装袋后用汽车拉运,取土和填土时保持原有土壤层次)进行小区设计,每个小区为长×宽×深=500 cm×200 cm×50 cm,每个小区种植220株,株距45 cm。3个重复。在2002年5月30日进行播种。
1.2.2 实生苗栽培
2010年 9月在东阿拉善-西鄂尔多斯分布的 8个四合木种群(乌家庙种群、桃司兔种群、千里山种群、乌达种群、海南种群、低山种群、石嘴山种群和东阿拉善种群)进行多基因库采样法(multiple genepool sampling)进行四合木种子采集,并在自然晾晒中予以充分混合,以保持其遗传多样性。于2011年各地区5月份终霜期过后,分别在乌海海勃湾区(以下简称乌海)、鄂尔多斯市康巴什新区(简称鄂尔多斯)和阿拉善盟的巴彦浩特(简称阿拉善)进行实生苗播种,采用拉丁方区组试验设计。
1.2.3 成株移植
在2010年2月,把苗龄为16年左右四合木成株 36株从其原分布区带冻土移植到东阿拉善的巴彦浩特镇的沙生植物园,株距×行距=65 cm×95 cm。
1.3 指标测定
1.3.1 植株高度与冠幅
在每一实验小区,随机选取植株,对在不同生育期、不同生境、不同方式条件下种植的四合木及原生境生长的白刺(Nitraria tangutorum),用2 m钢卷尺测定进行植株高度与南北冠幅的测定。
1.3.2 光合作用参数测定
2013年 8月 18─27日晴朗无风天气的07:00-19:00进行光合指标的测定,以乌海(原生境四合木成株和原生境的伴生土著种白刺成株、客土栽培四合木成株、四合木实生苗)、阿拉善(带土迁地保护四合木成株、实生苗)和鄂尔多斯栽培的四合木实生苗作为供试材料,每株选取向阳中部正常生长的成熟叶片,叶位一致,利用便携式气体交换系统(LI-6400XT,LiCor Inc. Nebraska,USA)从07:00-19:00,每隔1 h测定1次叶片的气体交换特性,自动记录光合有效辐射(Par)、叶片温度等,计算得到净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,μmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)等生理指标测定;叶温(Ta,℃)、空气相对湿度(RH%),光合有效辐射(PAR,μmol·m-2·s-1)、大气相对湿度(RH%)、大气CO2浓度(Ca,μmol·moL-1)大气水汽压亏缺(Vpd,KPa)等微气象参数由该仪器同时记录。测定中以空气 CO2浓度作为气源,变化范围为370~400 μmol·moL-1,温度 24~31 ℃,湿度16%~44%。每 1 h测定 1次,中午加密(在12:00-14:00时,每隔30 min测定1次)。每一小区测定3株,测定3个重复。
1.4 数据处理
对冠幅和净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等生理指标数据主要用 SPSS19.0软件对各光合作用参数进行处理,采用方差分析(ANOVA)进行差异显著性比较和LSD检验以及Duncan’s新复极差法多重比较。
2 结果与分析
分析了原生境四合木及其伴生种白刺、不同迁地试验区四合木植株生长量、光合作用日变化动态、气孔导度、胞间CO2浓度和蒸腾速率。
2.1 不同迁地试验区植株生长指标的差异性
在原生境土壤条件下生长的四合木成株的植株高度(73.75±3.17) cm和冠幅67.35 cm×65.45 cm明显高于客土栽培的四合木高度(49.77±2.58) cm和冠幅7.39 cm×47.65 cm。同时,四合木伴生种白刺(Nitraria tangutorum)的植株高度(98.55±7.55) cm和冠幅105.57 cm×108.73 cm明显高于在原生境土壤条件下生长的四合木成株。而四合木实生苗的生长高度大小依次为乌海四合木核心区实生苗(27.15±2.65) cm>鄂尔多斯实生苗(22.66±1.75) cm>阿拉善实生苗(19.05±1.24) cm。四合木实生苗的生长冠幅大小依次为乌海四合木核心区实生苗(26.48 cm×27.26 cm)>鄂尔多斯实生苗(21.27 cm×21.75 cm)>阿拉善实生苗(19.25 cm×18.27 cm)。四合木实生苗的分枝数大小依次为乌海四合木核心区实生苗(37.54±1.25)>鄂尔多斯实生苗(26.45±1.65)>阿拉善实生苗(24.25±1.45)。而且在原生境地乌海四合木核心区种植的实生苗与阿拉善实生苗之间的生长量存在显著差异(P≤0.01),与鄂尔多斯实生苗之间的生长量存在显著差异(P≤0.05)(表1)。
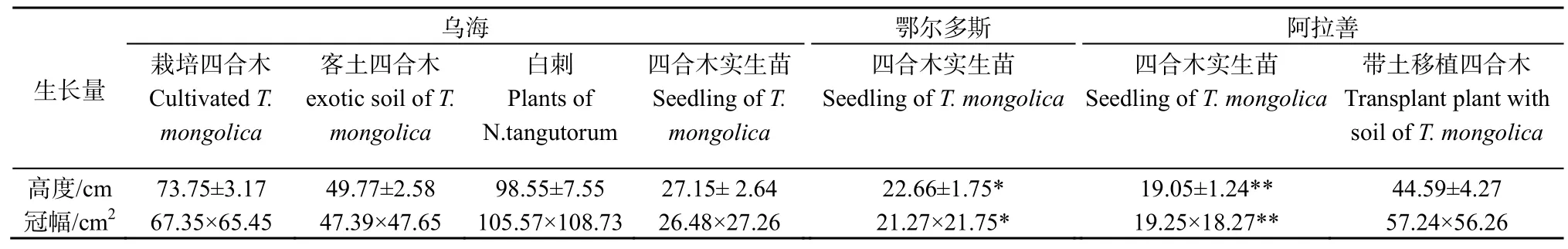
表1 不同生境条件下迁地试验区植株生长指标的比较Table 1 The comparison of growth characteristic value among the experiment blocks for relict plant Tetraena mongolica in ex-situ conservation
2.2 迁地保护条件下的四合木实生苗日光合作用特性的差异性
测定结果显示,迁地保护条件下四合木生实生苗植株叶片光合速率Pn日变化均呈“双峰”曲线,“午休”现象明显。早晨07:00时开始净光合速率均逐步增加。其中原生境四合木实生苗的主峰值分别出现在09:00时(14.7676 μmol·m-2·s-1)和12:00左右(14.4388 μmol·m-2·s-1),鄂尔多斯和阿拉善四合木实生苗的主峰值分别出现在10:00时(6.25795、3.43522 μmol·m-2·s-1)和 13:00左右(6.05886、11.8632 μmol·m-2·s-1),其中鄂尔多斯实生苗上午的光合速率明显高于下午,而阿拉善四合木实生苗的光合速率明显低于下午。光合速率表现出明显的分异(图2A)。
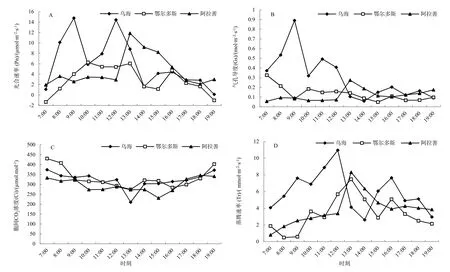
图2 不同迁地保护试验区四合木光合日变化特征的差异性Fig. 2 The photosynthetic characteristic differences among the original distributive area and different ex-situ conservation test area of T. monglica
气孔是气体进出植物体的主要通道,光合作用需要的CO2和蒸腾作用放出的水蒸汽都是通过气孔实现的。因此,气孔开度即气孔导度(Gs)和气孔限制值(Ls)的高低,对于Pn和Tr都有一定制约,进而影响WUE气孔导度是反映气孔开度的一个重要指标(赵丽丽等,2014)。气孔导度受叶片蒸腾作用影响,气孔导度的变化又对叶片蒸腾作用产生反馈调节作用(张金政等,2014)。从图2B可以看出,四合木气孔导度的日变化曲线均表现为明显的“双峰”曲线,与Pn与Tr的变化趋势基本相似,其中原生境四合木实生苗的主峰值分别出现在09:00时(0.8909 mol·m-2·s-1)和11:00左右(0.4923 mol·m-2·s-1),鄂尔多斯四合木实生苗的主峰值分别出现在10:00时(0.18406 mol·m-2·s-1)和16:00左右(0.108985 mol·m-2·s-1),阿拉善四合木实生苗的主峰值分别出现在11:00时(0.066286 mol·m-2·s-1)和13:00左右(0.274574 mol·m-2·s-1),其中鄂尔多斯实生苗上午的叶片气孔导度明显高于下午,而阿拉善四合木实生苗的叶片气孔导度明显低于下午。叶片气孔导度表现出明显的分化特征(图2A)。这可能与不同生境条件下光照及湿度等环境因子胁迫有关。
从四合木的胞间CO2浓度(Ci)的日变化特征看,Ci的日变化趋势均为一“V”型抛物线,早晨为1 d中最大值然后开始逐渐下降,在13:30左右出现低谷,然后又逐渐回升。三者的日变化曲线均为一单峰曲线,和Ci的变化趋势相反,早上为1 d最低值,日间不断增大,在13:30左右出现峰值,然后下午逐渐下降。据此判断四合木午间光合速率降低的同时伴随着Ci的降低和Ls的升高,原因主要是由气孔导度降低引起的。而15:30-16:30光合速率呈下降趋势,对应了Ci上升和Ls降低表明主要原因是非气孔因素叶肉细胞光合活性决定的。
蒸腾速率是反映植物蒸腾作用强弱最重要的生理指标。四合木实生苗叶片的Tr日变化均呈“双峰”曲线,其中原生境四合木实生苗的主峰值分别出现在12:00时(10.9512 μmol·m-2·s-1)和16:00左右(7.6473 μmol·m-2·s-1),鄂尔多斯和阿拉善四合木实生苗的主峰值分别出现在13:00时(7.4768、8.31087 μmol·m-2·s-1)和 16:30左右(5.07958、4.2475 μmol·m-2·s-1),其中乌海实生苗上午的蒸腾速率明显高于下午,而鄂尔多斯和阿拉善四合木实生苗生物的蒸腾速率却低于下午。迁地保护的四合木与原生境四合木实生苗的蒸腾速率呈现出明显的差异性(图2D)。
2.3 原生境条件下四合木成株与实生苗光合作用日变化特征的差异性
研究表明,原生境条件下四合木成株、实生苗与其伴生种白刺植株叶片光合速率 Pn日变化均呈“双峰”曲线(图3)。但光合作用日变化特征存在明显差异。白刺的光合效率明显高于四合木实生苗和成株,四合木实生苗又明显高于四合木成株。具体而言,白刺、四合木实生苗和成株三者的第1次主峰值均出现在09:00时(13.8057、14.7676、9.4122 μmol·m-2·s-1),但第2次主峰值分别出现在13:00、12:00和 13:00时(15.3805、14.4388、11.8537 μmol·m-2·s-1),表明白刺具有明显的光合生理适宜性和种间竞争优势(图3A)。从图3B可以看出,白刺和四合木气孔导度的日变化曲线均表现为递降的“双峰”曲线,与Pn与Tr的变动趋势基本相似,白刺、四合木实生苗和成株三者的第1次峰值均出现在09:00时(0.6216、0.8909、0.6532 mol·m-2·s-1),第2次峰值均不同步(图3C)。从三者的胞间CO2浓度(Ci)的日变化特征看,Ci的日变化趋势同样均为“V”型抛物线,早晨和晚上为最大值,在13:30左右出现低谷。白刺、四合木成株和实生苗三者的最低值从小到大依次为 132.2898、176.3196、209.930 μmol·m-2·s-1。
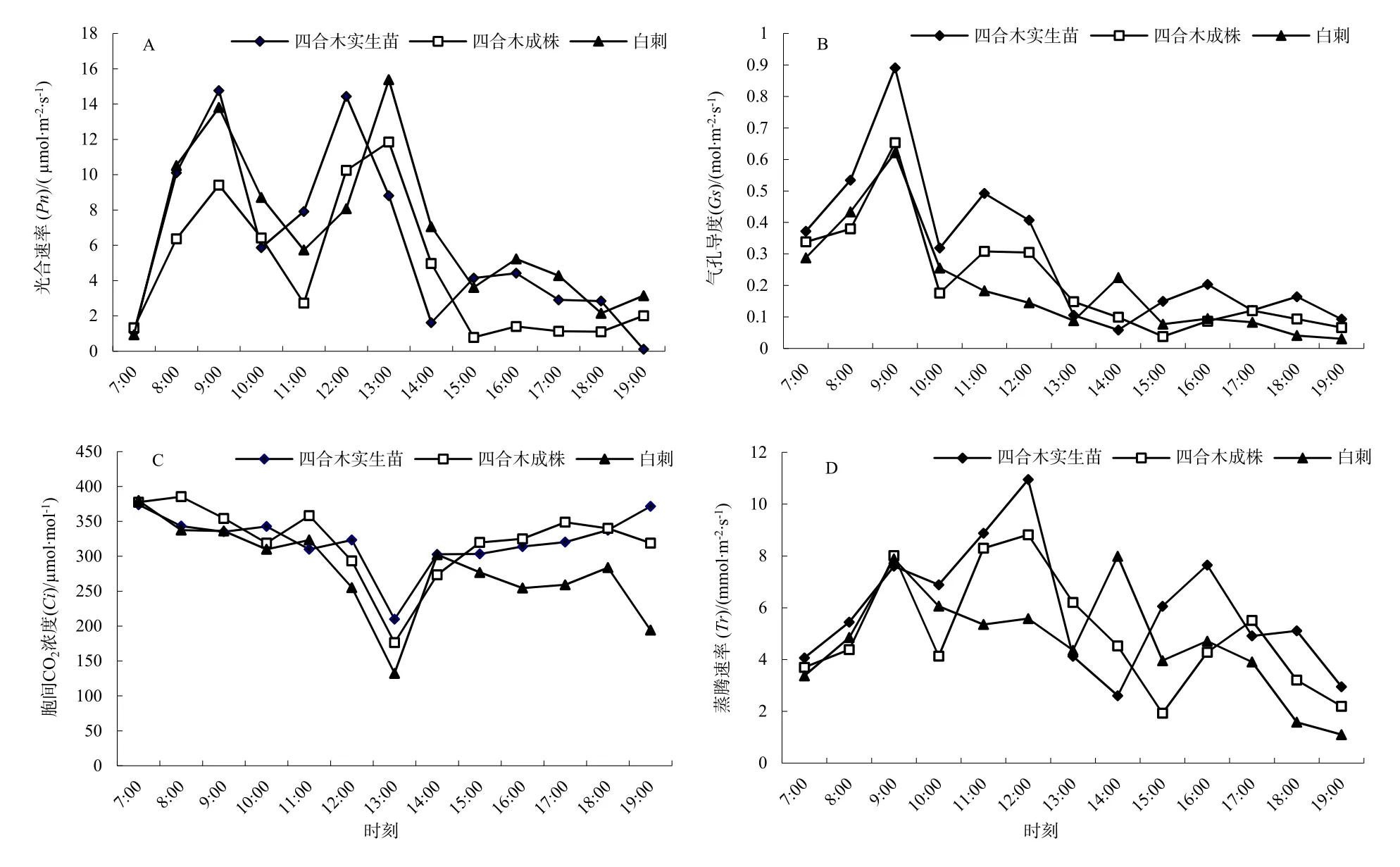
图3 原生境条件下四合木与其伴生种白刺光合作用日变化特征的差异性Fig. 3 The photosynthetic characteristic among the original distributive area of T. monglica and N. tangutorum
从蒸腾速率看,三者叶片的Tr日变化均呈“双峰”曲线,其中白刺、四合木成株和实生苗三者的主峰值分别出现在14:00、12:00和12:00时,峰值依次为7.9874、8.8182和10.9512 μmol·m-2·s-1,总的变化态势为以白刺的蒸腾速率为最低,四合木实生苗的蒸腾速率为最高(图3D)。
2.4 原生境生长的四合木与迁地保护成株四合木光合作用日变化特征的差异性
从迁地保护成株四合木的光合生理生态适宜性看,原生境生长的四合木与迁地保护成株四合木光合作用日变化也存在明显的差异性(图 4)。研究表明,原生境条件下四合木成株具有明显的光合生理生态适应性,其光合效率明显高于乌海栽培的四合木实生苗和阿拉善移植的成株。其中原生境条件下四合木成株光合的主峰值分别为为 9.4122和11.8537 μmol·m-2·s-1,而阿拉善移植的成株的主峰值分别为为2.872482和10.6842 μmol·m-2·s-1,乌海实生苗的光合速率介于二者之间(图4A)。
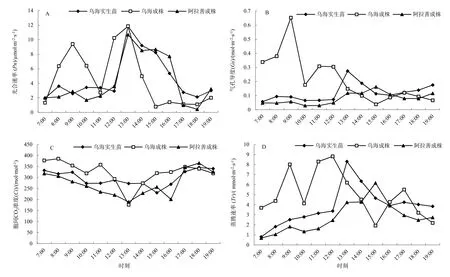
图4 原生境生长的四合木与迁地保护成株四合木光合作用日变化特征的差异性Fig. 4 The photosynthetic characteristic differences among the original distributive area and different ex-situ conservation test area of T.monglica
从图4B可以看出,原生境生长的四合木成株的气孔导度远高于阿拉善移植的四合木成株。原生境生长的四合木成株的气孔导度峰值分别为0.6532和0.3049 mol·m-2·s-1,而阿拉善移植的四合木成株峰值分别为0.0566和0.117373 mol·m-2·s-1。乌海实生苗的气孔导度在介于二者之间(图4C)。
从胞间CO2浓度看,Ci的日变化趋势同样均为“V”型抛物线。原生境生长的四合木成株、实生苗与迁地保护成株四合木胞间CO2浓度的最低值从小到大依次为 188.9385、176.3196、230.975 μmol·m-2·s-1,但总的变化趋势为以阿拉善移植的四合木成株为最低。
从蒸腾速率看,原生境生长的四合木成株的主峰值分别出现在12:00(8.8182 μmol·m-2·s-1)和17:00时(5.5094 μmol·m-2·s-1),而迁地保护成株四合木主峰值分别出现在09:00(1.815766 μmol·m-2·s-1)和15:00时(6.1633 μmol·m-2·s-1)(图4D)。
3 讨论
3.1 原生境条件下四合木的生长特性与光合日变化特征
光合作用是地球上规模最大的生物合成过程。由于内部和外部因素的限制,植物的光合作用往往不能高效率地进行。植物的气孔、RuBP羧化、RuBP再生、P再生和光合产物等生理因素以及光照、温度、水分、空气、矿质营养、环境等共同作用构成了光合作用的限制因素。其中光对光合作用的主要作用表现为提供同化力形成所需要的能量、活化光合作用的关键酶和促使气孔开放、调节光合机构的发育(余叔文和汤章城,1998)。在原生境土壤条件下生长的四合木成株的植株高度(73.75±3.17) cm和冠幅67.35 cm×65.447 cm明显高于客土栽培的四合木高度(49.77±2.58) cm 和冠幅 47.39 cm×47.66cm。白刺的植株高度(98.55±7.55) cm和冠幅105.57 cm×108.73 cm明显在原生境土壤条件下生长的四合木成株(P≥0.05)。同时,白刺的光合效率明显高于四合木实生苗和成株,四合木实生苗又明显高于四合木成株。已有的研究表明,在甘肃民勤绿洲荒漠过渡带,白刺比梭梭具有更好的生理生态适应性(赵长明等,2005)。在西鄂尔多斯,净光合速率日均值依次为沙冬青(5.97 μmol·m-2·s-1)>霸王(4.5 μmol·m-2·s-1)>四合木(3.05 μmol·m-2·s-1)。同生境下四合木的净光合速率、蒸腾速率、水分利用效率低于霸王和白刺,表明在三者混生的环境下,四合木光合能力及对干旱环境适应性弱于霸王和白刺,说明四合木在生理生态学方面存在生存力适应力弱化的特性(石松利等,2012)。四合木进入成年期后,在群落中处于不利的种间竞争中地位,有让位于伴生种白刺和近缘种霸王的趋势(智颖飙等,2008;石松利等,2012;祖元刚等,2002)。
3.2 迁地保护条件下四合木的光合日变化与生理生态适应性
大多数濒危植物的光合、呼吸和蒸腾等生理代谢速率比对照种在相同条件下要低(祖元刚等,2002)。研究表明,迁地保护条件下四合木生实生苗植株叶片光合速率Pn日变化均呈“双峰”曲线。其中原生境四合木实生苗的主峰值分别出现在09:00时和12:00时,鄂尔多斯和阿拉善四合木实生苗的主峰值分别出现在10:00时和13:00时,同时鄂尔多斯实生苗上午的光合速率明显高于下午,而阿拉善四合木实生苗的光合速率明显低于下午。乌海、鄂尔多斯和阿拉善种植的四合木的光合速率、气孔导度、胞间CO2浓度和蒸腾速率均表现出明显的分异。
由于濒危植物四合木种群的分布区日趋狭小,景观破碎化程度愈来愈高,形成了岛屿隔离状态,进而导致基因交流日趋困难和遗传力衰退。四合木生理生态学适应能力的逐步弱化,出现了生理代谢与环境条件相互适应方面的分化(智颖飙等,2008)。
4 结论
根据上述分析与讨论,认为尽管四合木自然分布区西鄂尔多斯与四合木异地保护区的东阿拉善和东鄂尔多斯地区的光照、水分等气候生物学特征和土壤类型存在明显差异(智颖飙等,2008;智颖飙等,2014),同时迁地种植的四合木实生苗的生长量也存在一定的差异,进而表现在光合作用日变化、蒸腾速率、胞间CO2浓度和气孔导度也存在明显差异。鉴此得出如下结论:
(1)四合木实生苗的生长南北冠幅大小依次为乌海四合木核心区实生苗(26.476 cm×27.257 cm)>鄂尔多斯实生苗(21.267 cm×21.745 cm)>阿拉善实生苗(19.249 cm×18.265 cm)。
(2)在原生境地乌海四合木核心区种植的实生苗与阿拉善异地种植实生苗之间的生长量存在显著差异(P≤0.01),与鄂尔多斯异地种植实生苗之间的生长量存在显著差异(P≤0.05)。
(3)迁地保护条件下四合木生实生苗植株叶片光合速率 Pn日变化均呈“双峰”曲线。不同试验区四合木光合作用日变化(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)均表现出明显的分异。迁地保护四合木条件下原生境地栽培的四合木实生苗的光合速率>鄂尔多斯栽培的四合木实生苗的光合速率>阿拉善栽培的四合木实生苗。
(4)由此做出进一步推断,孑遗植物四合木从原生境地东移进行迁地保护具有更高的生理生态适宜性和适应性。
目前进行的四合木活植物整体的迁地以及种子播种的实生苗的迁地保护,未来如何建立有效种群,提升其抗逆性和各个生育阶段的适宜性和适宜性,逐步建立多种类、多层次人工群落,完成“从种子到种子”的起码标准,进一步实现“保存性和代表性”,最终实现四合木迁地保护的“保持性和防止性”(Zhi等,2014;智颖飙等,2014;张金政等,2014;吴建国等,2009;许再富等,2008;许再富,199897-104),最终使孑遗濒危特有植物四合木的异地保护由可行变为现实,仍需做进一步的深入研究。
BRÜTTING C, HENSEN I, WESCHE K. 2013. Ex situ cultivation affects genetic structure and diversity in arable plants[J]. Plant Biology, 15(3): 505-513.
DANIEL LAUTERNACH, MICHAEL BURKART, BIRGIT GEMEINHOLZER. 2012. Rapid genetic differentiation between ex situ and their in situ source populations: an example of the endangered Silene otites (Caryophyllaceae) [J]. Botanical Journal of the Linnean Society, 168 (1): 64-75. HUANG HONGWEN. 2011. Plant diversity and conservation in China: planning a strategic bio-resource for a sustainable future[J]. Botanical Journal of the Linnean Society, 166(3): 282-300.
JASON D HOEKSEMA, V BALA CHAUDHARY, CATHERINE A GEHRING, et al. 2010. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi [J]. Ecology Letters, 13: 394-407.
MOIR M L, VESK P A, BRENNAN K E, et al. 2012. Considering extinction of dependent species during translocation, ex situ conservation, and assisted migration of threatened hosts [J]. Conservation Biology, 26(2): 199-207.
ZHI YINGBIAO, DING LING, WANG YUPENG, et al. 2014. Isolation and characterization of thirteen novel dinucleotide micro-satellite Loci from Tetraena mongolica Maxim [J]. Conservation Genetics Resources, 16: 297-299.
傅立国, 金鉴明. 1991. 中国植物红皮书——稀有濒危植物[M]. 北京:科学出版社: 712-713.
谷安琳, 王宗礼. 2009. 中国北方草地植物彩色图谱[M]. 北京: 中国农业科学技术出版社: 139-144.
国家林业局野生动植物与自然保护区管理司, 中国科学院植物研究所. 2013. 中国珍稀濒危植物图鉴[M]. 北京: 中国林业出版社: 366-378.
黄会青, 钱荣. 2006. 浙江林业[J]. 1: 41.
刘钟龄, 雍世鹏, 王义风, 等. 1985. 内蒙古植被[M]. 北京: 科学出版社.
马毓泉. 1989. 内蒙古植物志: 第3卷[M]. 2版. 呼和浩特: 内蒙古人民出版社: 428-429.
石松利, 王迎春, 周红兵, 等. 2012. 濒危种四合木与其近缘种霸王水分关系参数和光合特性的比较[J]. 生态学报, 32(4): 1163-1173.
王峰, 杨持. 2003. 四合木的自然更新和就地保护的研究[J]. 内蒙古大学学报: 自然科学版, 34(2): 196-202.
吴建国, 吕佳佳, 艾丽. 2009. 气候变化对生物多样性的影响: 脆弱性和适应[J]. 生态环境学报, 18(2): 693-703.
徐朗然, 黄成就. 1998. 中国植物志(第43卷第1分册)[M]. 北京: 科学出版社: 144-145.
许再富, 黄加元, 胡华斌, 等. 2008. 我国近 30年来植物迁地保护及其研究的综述[J]. 广西植物, 28(6): 764-774.
许再富. 1998. 稀有濒危植物迁地保护的原理与方法[M]. 昆明: 云南科技出版社.
余叔文, 汤章城. 1998. 植物生理与分子生物学 [M]. 2版. 北京: 科学出版社: 155-276.
张金政, 张起源, 孙国峰, 等. 2014. 干旱胁迫及复水对玉簪生长和光合作用的影响[J]. 草业学报, 23(1): 167-176.
张韬, 王炜, 安慧君, 等. 2005. 东阿拉善-西鄂尔多斯地区特有濒危植物种群斑块变化与优先保护级相关分析[J]. 干旱区资源与环境, 19(5): 179-184.
赵长明, 魏小平, 尉秋实, 等. 2005.民勤绿洲荒漠过渡带植物白刺和梭梭光合特性[J]. 生态学报, 25(8): 1908-1913.
赵丽丽, 王普昶, 陈超, 等. 2014. 干旱胁迫对百脉根叶绿素荧光特性的影响[J]. 中国草地学报, 36(1): 18-23.
智颖飙, 王再岚, 王中生, 杨持, 等. 2008. 孑遗植物四合木(Tetraena mongolica)的濒危肇因与机制[J]. 生态学报, 28(2): 767-776.
智颖飙, 杨持, 李红丽, 等. 2014. 孑遗植物四合木(Tetraena mongolica)异地保护条件下的气候生物学特征与光合效率[J]. 中国沙漠, 34(1): 1-8.
智颖飙. 2004. 四合木(Tetraena mongolica)异地保护生物学基础研究[D].内蒙古大学博士学位论文.
周云龙. 2011. 植物生物学[M]. 3版. 北京: 高等教育出版社: 162-196.
祖元刚, 王文杰, 杨逢建. 2002. 植物生活史型的多样性及动态分析[J].生态学报, 22(11): 1811-1818.
The Studies on the Photosynthetic Characteristics of the Endemic Relict Shrub Tetraena mongolica Maxim for the ex-situ Conservation
ZHI Yingbiao1,2,3, LI Hongli4, CUI Yan5, LU Zhanyuan5, LIU Pei1, YE Xuehua6, ZHANG HeLiang3*, YANG Chi2, LIU Zhongling2, WANG Yunfei3, HUA Yupeng3, HONG Ge3, ZHAO Kai7, WEI LingLing6, WANG Qiang8
1. College of Environment and Resources, Inner Mongolia University, Huhhot 010021, China; 2. College of Life Science, Inner Mongolia University, Huhhot 010021, China; 3. Ordos College, Inner Mongolia University, Ordos 017000, China; 4. College of Nature Conservation, Beijing Forestry University, Beijing 100083, China; 5. Inner Mongolia Academy of Agricultural and Animal Husbandry, Huhhot 010031, China; 6. State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, the Chinese Academy of Sciences, Beijing 100093, China; 7. College of Geographical Science, Nanjing Normal University, Nanjing 210039, China; 8. Management Bureau of Natural Conservation Region in Western Ordos, Wuhai 016000, China
In order to further clarify the photosynthesis and physiological adaptability of relict plant Tetraena mongolica under the condition of ex-situ conservation, instantaneous photosynthetic efficiency is analyzed by applying the LI-6400 photosynthes measurement system and the T. mongolica in the ex-situ conservation area and the diurnal photosynthesis changes of Nitraria tangutorum as well as the growth are determined. The results show that: the growth of T. mongolica seedlings crown is in the order of the core area of Wuhai T. mongolica seedlings (26.48 cm×27.26 cm)>Ordos seedlings (21.27 cm×21.75 cm)>Alxa seedlings (19.25 cm×18.27 cm). There are significant differences (P≤0.01) in growth between the original habitat Alashan seedlings and the Wuhai T. mongolica in the core area planted and also there is a significant difference among the growth of Ordos seedlings (P≤0.05). Under the conditions of ex situ conservation, the photosynthetic rate of T. mongolica seedlings leaves shows diurnal variation of“bimodal” curve. T. mongolica diurnal variation of photosynthesis (Pn) in different test areas, transpiration rate(Tr), stomatal conductance(Gs), intercellular CO2and concentration show a clear difference. Under the condition of ex-situ conservation, photosynthetic rate of T. mongolica in the original habitat > photosynthetic rate of Ordos T. mongolica seedlings>Alxa T. mongolica seedlings. Furthermore, relict plant T. mongolica, with a higher physiological habitat adaptability and eco-suitability, can be carried eastward from the former ex-situ conservation. But in order to realize “seed to seed” ex-situ to “conserve and represent”, and ultimately to achieve “retention and prevention” of T. mongolica ex situ conservation, the ultimate success of ex-situ conservation of T. mongolica remains to be continued to further research.
ex-situ conservation; habitat fitness; photosynthesis; retention; multiple genepool; Tetraena mongolica Maxim
Q945.79
A
1674-5906(2015)01-0014-08
10.16258/j.cnki.1674-5906.2015.01.003
智颖飙,李红丽,崔艳,路战远,刘珮,叶学华,张荷亮,杨持,刘钟龄,王云飞,华宇鹏,红鸽,赵凯,魏玲玲,王强. 孑遗植物四合木(Tetraena mongolica)迁地保护中的光合作用日变化特征与生理生态适应性[J]. 生态环境学报, 2015, 24(1): 14-21.
ZHI Yingbiao, LI Hongli, CUI Yan, LU Zhanyuan, LIU Pei, YE Xuehua, ZHANG HeLiang, YANG Chi, LIU Zhongling, WANG Yunfei, HUA Yupeng, HONG Ge, ZHAO Kai, WEI LingLing, WANG Qiang. The Studies on the Photosynthetic Characteristics of the Endemic Relict Shrub Tetraena mongolica Maxim for the ex-situ Conservation [J]. Ecology and Environmental Sciences, 2015, 24(1): 14-21.
国家自然科学基金项目(30160020);内蒙古科技创新引导基金项目(20121604);内蒙古创新人才团队基金项目(20120341);内蒙古自然科学基金项目(2011MS0501);内蒙古人才开发基金(2014RC1218)项目
智颖飙(1965年生),男,教授,博士后,主要从事植物种群生态学研究。*通信作者:Email: zhl_77@imu.edu.cn
2014-11-20
